




Перемещение веществ в клетку или из нее в окружающую среду может осуществляться многими способами. В зависимости от того, чем представлен источник энергии для переноса веществ, что является движущей силой перемещения, все виды переноса веществ можно разделить на пассивный и активный транспорт. Пассивный транспорт веществ всегда осуществляется за счет энергии, сконцентрированной в каком-либо — градиенте, и энергия метаболических процессов клеток (энергия гидролиза АТФ) непосредственно на этот перенос не расходуется. Пассивный транспорт всегда проходит по направлению градиентов, т.е. от более высокого энергетического уровня к более низкому. В результате этого градиенты уменьшаются, если нет других процессов, обеспечивающих их поддержание на постоянном уровне. Часто в клетках одновременно имеется несколько градиентов, которые накладываются друг на друга, — суперпозиция градиентов. В таком случае пере-
139
нос вещества будет определяться результирующей всех градиентов. Если градиенты имеют одинаковое направление, то результирующий перенос будет равен сумме потоков вещества по отдельным градиентам. В случае двух противоположных по направлению градиентов результирующий перенос вещества будет равен разности потоков по отдельным градиентам и иметь направление, совпадающее с направлением того градиента, абсолютная величина которого больше. Понятно, что при наличии двух равных по абсолютной величине, но противоположных по направлению градиентов результирующий перенос вещества будет равен нулю.
Основными градиентами, присущими живым организмам, являются концентрационные, осмотические, электрические и градиенты гидростатического давления жидкости.
В соответствии с этими градиентами имеются следующие виды пассивного транспорта веществ в клетках и тканях: диффузия, осмос, электроосмос и аномальный осмос, фильтрация.
ДИФФУЗИЯ
Основным механизмом пассивного транспорта веществ, обусловленным наличием концентрационного градиента, является диффузия. Диффузия — это самопроизвольный процесс проникновения вещества из области большей в область меньшей его концентрации в результате теплового хаотического движения молекул.
 Математическое описание процесса диффузии дал
Математическое описание процесса диффузии дал
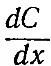 Фик. Согласно закону Фика, скорость диффузии прямо пропорциональна градиенту концентрации и
Фик. Согласно закону Фика, скорость диффузии прямо пропорциональна градиенту концентрации и
площади S, через которую осуществляется диффузия:

(1)
Под скоростью диффузии понимают количество вещества (в молях или других единицах), диффундирующего в единицу времени через данную площадь. Градиент концентрации — это изменение концентрации С вещества, приходящееся на единицу длины, в направлении диффузии. Знак минус в правой части уравнения (1) показы-
140
мает, что диффузия происходит из области большей концентрации в область меньшей концентрации вещества. Коэффициент пропорциональности D в уравнении (1) называется коэффициентом диффузии. Его физический
 |
смысл легко выяснять, если S и приравнять к единице. Коэффициент диффузии численно равен количеству вещества, диффундирующего в единицу времени через единицу площади при градиенте концентрации, равном единице. Коэффициент диффузии зависит от природы вещества и от температуры. Он характеризует способность вещества к диффузии.
Так как концентрационный градиент клеточной мембраны определить трудно, то для описания диффузии веществ через клеточные мембраны пользуются более простым уравнением, предложенным Коллендером и Берлундом:
 (2)
(2)
где С 1и С 2 — концентрации вещества по разные стороны мембраны; Р — коэффициент проницаемости, аналогичный коэффициенту диффузии. В отличие от коэффициента диффузии, который зависит только от природы вещества и температуры, Р зависит еще и от свойств мембраны и от ее функционального состояния.
Проникновение растворенных частиц, обладающих электрическим зарядом, через клеточную мембрану зависит не только от концентрационного градиента, но и от электрического градиента мембраны. В связи с этим перенос ионов может осуществляться в направлении, противоположном концентрационному градиенту, при наличии противоположно направленного электрического градиента. Совокупность концентрационного и электрического градиентов называется электрохимическим градиентом. Пассивный транспорт ионов через мембраны всегда происходит по электрохимическому градиенту.
Клетки как термодинамические системы могут находиться в стационарном и переходном состояниях. При стационарном состоянии скорости поступления и выведения веществ через клеточные мембраны неизменны во времени. Это обусловлено ролью метаболических процессов, обеспечивающих постоянство концентрационных градиентов клеток. Так, включение поступающего в
141
|
|

клетку аниона фосфора в молекулы нуклеотидов, нуклеиновых кислот, нуклеопротеидов и фосфолипидов приводит к снижению концентрации неорганического фосфата в клетке и тем самым поддерживает градиент этого аниона между клеткой и средой. Указанное положение справедливо не только для диффузии веществ в клетку, но и для диффузии продуктов обмена из клетки в среду. Однако этим роль метаболических процессов клетки по отношению к транспорту веществ не исчерпывается. Все клетки имеют механизмы обеспечивающие перенос веществ против их электрохимических градиентов с затратой энергии метаболических процессов клетки. Такой перенос называют активным транспортом веществ. В результате активного переноса клетки способны накапливать вещества в более высоких по сравнению со средой концентрациях и тем самым совершать концентрационную (осмотическую) работу. Более подробно эти процессы будут описаны ниже. Здесь отметим, что многие градиенты, присущие клетке и являющиеся условием для пассивного переноса, возникают именно в результате активного транспорта веществ. Так, концентрационные градиенты калия и натрия возникают в результате активного переноса этих ионов (в результате работы натрий-калиевого насоса); в последующем происходит диффузия этих ионов по градиентам и генерирование мембранных потенциалов.
Если молекулы диффундирующего вещества движутся, без образования комплекса с другими молекулами,
то такая диффузия называется простой. Помимо
этого, существует облегченная и обменная диффузия.
Было обнаружено, что скорость проникновения в клетку глюкозы, глицерина, аминокислот и некоторых
142
других веществ не имеет линейной зависимости от разности концентраций. При каких-то концентрациях веществ скорость их проникновения намного больше, чем следует ожидать для простой диффузии. При увеличении разности концентраций скорость проникновения возрастает в меньшей степени, чем это следует из уравнения (2). Приведенные особенности проникновения веществ объясняются тем, что в данном случае наблюдается не простая, а облегченная диффузия. Механизм ее состоит в том, что данное вещество самостоятельно слабо, диффундирует через мембрану, но скорость диффузии намного возрастает, если молекулы А этого вещества образуют комплекс с молекулами X вспомогательного вещества, которое, как полагают, растворяется в липидах. На поверхности мембраны молекулы А соединяются с молекулами X и в виде комплекса АХ диффундируют в клетку (рис. 24, I). В клетке молекулы А освобождаются, а молекулы вспомогательного вещества диффундируют наружу и связываются с новыми молекулами А. Понятно, что в приведенной схеме перенос вещества Л, как и при простой диффузии, будет идти до тех пор, пока его концентрации не выравниваются по обе стороны мембраны.
Если концентрацию вещества в среде повысить до такой степени, что все молекулы вещества X, облегчающего диффузию, будут использованы, то скорость диффузии при дальнейшем повышении концентрации вещества А возрастать не будет. В общем случае кинетика облегченной диффузии подчиняется правилу Михаэлиcа — Ментен для ферментативных реакций (см. главу 4).
Облегченная диффузия может осуществляться не только с помощью подвижных, но и с помощью фиксированных в мембране переносчиков. Цепочка переносчиков может выстилать изнутри пору, а молекула проникающего вещества способна передвигаться от одного се звена к другому, что несколько напоминает процессы и мультиферментных системах (рис. 24, II). При этом предполагается, что пространство в поре недостаточно велико для прохождения через нее частиц, если только они не способны к специфическому взаимодействию с переносчиками. Механизм участия вспомогательного вещества в диффузии подтверждается также наличием конкуренции между веществами при их проникновении в клетку. Так, поступление глюкозы в мышечные волокна
143
сердца крысы уменьшается при внесении в среду ксилозы, арабинозы и манозы. Это уменьшение обусловлено тем, что добавленные в среду сахара используют часть молекул вспомогательного вещества X для своего проникновения. Наконец, возможен и такой вариант, что какое-то вещество образует с X очень прочный комплекс, так что возможность соединения X с другими веществами исключается. Предполагают, что подобный механизм блокирования молекул вспомогательного вещества лежит в основе действия известных ингибиторов транспорта пентоз и гексоз — флоридзина и флоретина.
В настоящее время природа соединений, облегчающих диффузию, еще не установлена, хотя большинство авторов предполагают, что это подвижные комплексы, растворимые в липидах клеточной мембраны. Этим объясняется проникновение в клетку веществ, не растворимых в липидах и имеющих молекулы, большие по размерам, чем диаметр клеточных пор.
Все особенности кинетики облегченной диффузии показывают, что она является каталитическим процессом, аналогичным ферментативной реакции. Поэтому облегченную диффузию часто называют каталитической диффузией, где роль катализатора играет вещество, облегчающее проникновение. Феномен катализируемого проникновения добавляет к классической картине мембранной структуры новую подробность — наличие особых каталитических единиц, которые, подобно ферментам, специфически взаимодействуют с проникающим веществом. Как и ферменты, они не могут вызвать процессов невозможных по термодинамическим условиям, т. е. катализ сам по себе не означает перенос вещества против градиента. Он лишь ускоряет проникновение веществ через мембрану по сравнению с обычной диффузией.
Разновидностью облегченной диффузии является так называемая обменная диффузия, понятие о которой введено Уссингом. При обменной диффузии вспомогательное вещество образует соединение с диффундирующим веществом и перемещается к другой поверхности мембраны. На другой поверхности мембраны молекула проникающего вещества освобождается и на ее место присоединяется другая молекула этого же вещества и переносится обратно. Таким образом, образуется замкнутый круг и концентрация диффундирующего вещества по
144
обе стороны мембраны не изменяется. Наличие обменной диффузии было доказано методом радиоактивных изотопов на эритроцитах. Было показано, что натрий эритроцитов благодаря обменной диффузии быстро обменивается на натрий плазмы. Имеются также данные, что в результате обменной диффузии одно вещество может обмениваться в эквивалентных количествах на другое вещество, находящееся на противоположной стороне мембраны.
В последнее время получен ряд веществ, названных ионофорами, которые способны избирательно усиливать диффузию ионов через мембраны. Например, антибиотик валиномицин, образующий комплекс с ионами калия, повышает на несколько порядков проницаемость мембраны для калия. Открыты также соединения, способные избирательно блокировать диффузию ионов через мембраны. К последним относится яд рыбы фугу тетродотоксин, который блокирует диффузию натрия. Клетки, обработанные тетродотоксином, утрачивают способность к возбуждению.
Исследование механизма действия на мембрану подобных веществ представляет большой интерес для медицины, поскольку это открывает перспективы создания новых эффективных анестетиков.
Как уже отмечалось, диффузия является основным видом пассивного транспорта веществ в клетках. Все остальные виды пассивного переноса относятся главным образом к транспорту воды. Вода играет важную роль в жизнедеятельности клеток, поэтому изучение проницаемости клеток и тканей для нее имеет большое значение. Проницаемость клеток для воды весьма велика. Это связано с тем, что молекулы воды имеют небольшие размеры и легко проникают через поры мембран. Проникновение воды в клетки и ткани может осуществляться в результате осмоса, ультрафильтрации и электроосмоса.
ОСМОС
Клеточные мембраны обладают свойством полупроницаемости, т.е. способностью хорошо пропускать одни вещества, в частности воду, и плохо пропускать другие вещества. В связи с этим вода проходит через клеточные мембраны в основном в результате осмоса. Осмос — это движение молекул воды через полупроницаемую

