




Выше были рассмотрены основные виды пассивного транспорта веществ в клетках. Как указывалось, при массивном транспорте энергия метаболических процес-
157
сов не расходуется, а перенос веществ в клетках осуществляется по электрохимическому градиенту. Вследствие этого градиенты клетки имеют тенденцию к уменьшению — пассивный перенос всегда стремится выровнять неравномерность в распределении веществ между клеткой и средой. Но, как показывает опыт, клеточное содержимое резко отличается по своему составу от окружающей клетку среды. В клетке содержатся в больших количествах ионы калия, недиффундирующие ионы белков, фосфолипидов, анионы аминокислот и другие ионы органических веществ, содержание которых в жидкости, окружающей клетку, незначительно. Другие вещества, напротив, в значительно более высоких концентрациях содержатся в окружающей жидкости, чем в клетке, например ионы натрия. Такое неравномерное распределение веществ между клеткой и средой не может быть полностью объяснено наличием только пассивного транспорта веществ, который, как указывалось, всегда приводит к уменьшению электрохимических градиентов.
Вышеприведенные данные, а также ряд других, позволяют сделать заключение, что в клетках наряду с пассивным происходит и активный транспорт веществ. Перенос молекул и ионов против электрохимического градиента, осуществляемый клеткой за счет энтропии метаболических процессов, называется активным транспортом.
Осуществляя перенос веществ против градиента, клетка совершает определенную работу, которая называется концентрационной или осмотической. Эта работа состоит, с одной стороны, в накоплении клеткой веществ,
присутствующих в окружающей среде в малых количествах, а с другой — в выведении в окружающую среду веществ, которые в самой этой среде присутствуют в высоких концентрациях.
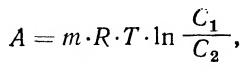 Величина концентрационной работы при переносе незаряженных частиц только против сил диффузии (против концентрационного градиента) может быть найдена из уравнения: (7)
Величина концентрационной работы при переносе незаряженных частиц только против сил диффузии (против концентрационного градиента) может быть найдена из уравнения: (7)
| где А — работа; т — количество молей вещества, перенесенного через мембрану из области с активной кон- |
| 158 |
центацией вещества С 2 в область с концентрацией С 1 R — газовая постоянная; Т — абсолютная температура. Если происходит перенос ионов через электрически поляризованную мембрану, то работа совершается не только против сил диффузии, но и против сил электрического поля мембраны. В этом случае общая работа А может быть найдена из уравнения:
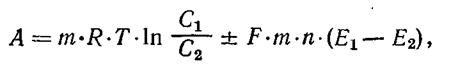 (8)
(8)
где m— количество перенесенных грамм-ионов вещества; п — валентность ионов; F = 96 500 Кл — число Фарадея — заряд 1 г-экв ионов; Е 1— Е 2— разность потенциалов между поверхностями мембраны; С 1и С 2 — активные концентрации ионов по одну и по другую стороны мембраны. Знаки плюс и минус в уравнении (8) показывают, что в зависимости от знака заряда ионов электрический градиент мембраны может иметь направление, либо совпадающее с направлением концентрационного градиента (плюс), либо противоположное ему (минус). Соответственно и работа по переносу ионов в электрохимическом градиенте равна либо сумме работ по переносу в концентрационном и электрическом градиентах, либо их разности.
Большинство гипотез механизма активного переноса базируется на предположении о существовании специальных переносчиков, захватывающих переносимое
сквозь мембрану вещество и доставляющих его на другую сторону клеточной мембраны. Возможно, что этот
переносчик пространственно ориентирован внутри мембраны в направлении, перпендикулярном ее поверхности, что дает ему возможность образовывать комплекс с переносимой молекулой и в то же время ориентировать её проникновение в клетку.
Исходя из данных, имеющихся к настоящему времени, активный транспорт веществ разделяют на два вида: активный перенос ионов и активный перенос органических веществ, главным образом Сахаров и аминокислот. По-видимому, к активному транспорту следует отнести и такое явление, как пиноцитоз — «питье клеток», когда вещества поступают в клетку «упакованными» в клеточную мембрану.
Явление активного переноса ионов к настоящему времени обнаружено у большого количества клеток и тка-
159
ней. Интересный пример активного переноса представляет кожа лягушки. Крогом было обнаружено, что эта ткань специализируется на переносе ионов натрия из окружающей среды в интерстициальную жидкость, где их концентрация в 10 000 раз выше, чем в среде. Именно такой механизм лежит в основе снабжения тканей лягушки этими необходимыми для ее жизнедеятельности ионами. Впоследствии Уссингом с сотрудниками был разработан метод изучения активного транспорта натрия через кожу лягушки. Если разделить две камеры, содержащие нормальный раствор Рингера, свежеизолированной кожей лягушки, то внутренняя поверхность кожи заряжается положительно по отношению к наружной. Разность потенциалов при этом достигает 100 мВ. Это так называемая «насосная» разность потенциалов, возникновение которой обусловлено активным переносом натрия. Подавая от внешнего источника ЭДС напряжение противоположного направления, можно скомпенсировать разность потенциалов через кожу. В этих условиях перенос ионов через кожу под влиянием электрического поля невозможен, так же как и путем диффузии, так как растворы по обе стороны кожи идентичны. Однако продолжает иметь место перенос ионов натрия от наружной поверхности к внутренней, в чем можно убедиться, используя меченный натрий. О том, что имеет место однонаправленный перенос ионов натрия, можно судить и по наличию тока в компенсационной цепи. Поскольку разность потенциалов скомпенсирована, то ток в компенсационной цепи численно равен току, возникающему благодаря одностороннему переносу через кожу положительных зарядов, т. е. ионов натрия.
Перенос натрия через кожу лягушки непосредственно связан с обменом веществ. Многочисленные ингибиторы окислительных процессов приводят немедленно к снижению или прекращению переноса ионов натрия и к исчезновению разности потенциалов через кожу. Аналогичные явления наблюдались и в опытах с другими сложными мембранными системами.
Перенос ионов отчетливо выражен в различных секреторных клетках — в слюнных и потовых железах и особенно в железах желудка, выделяющих большие количества ионов водорода и хлора.
О высокой интенсивности процессов активного переноса ионов в клетках свидетельствуют данные термоди-
160
намических расчетов. Высчитано, что 10% энергии метаболизма мышцы лягушки в покое расходуется на перенос ионов натрия. Этот показатель возрастает до 50% в определенных условиях опыта, стимулирующих (перенос ионов.
Большое количество работ посвящено исследованию активного переноса ионов калия и натрия через клеточные мембраны. Это объясняется их большой ролью в таких важных явлениях, как генерирование биоэлектрических потенциалов и проведение возбуждения.
Как указывалось, ионы калия и натрия неравномерно распределены между цитоплазмой клеток и межклеточной жидкостью. В то же время данные, полученные с помощью изотопных методов, показали, что эти ионы могут диффундировать через мембраны. С помощью этих же методов было установлено, что при каждом акте возбуждения клетки приобретают определенное количество ионов натрия и теряют эквивалентное количество ионов калия. В свете этих данных становится ясным, что поддержание электрохимических градиентов калия и натрия неизменными на протяжении всей жизни клеток возможно только в том случае, если эти ионы переносятся специальными механизмами против градиентов. Этот перенос будет компенсировать уменьшение градиентов, вызванное диффузией. И действительно, непосредственные измерения с помощью радиоактивных 42K и 24Na показали, что большинство изученных клеток находятся в стационарном состоянии. При этом скорость поступления данного иона внутрь клетки равна скорости потока ионов того же вида из клетки во внешнюю среду.
Кроме того, было установлено, что при выключении гликолиза и окислительного фосфорилирования ингибиторами метаболических процессов (йод уксусная кислота, цианиды, динитрофенол и др.) неравномерность в распределении ионов уменьшается вплоть до полного выравнивания их концентраций в клетке и во внешней среде. Если же путем микроинъекции в отравленное нервное волокно кальмара ввести некоторое количество ЛТФ, то перенос ионов против градиентов возобновляется. Неравномерность в распределении ионов уменьшается также при охлаждении клеток, приводящем к замедлению метаболических процессов. Так, при охлаждении эритроцитов человека до 2°С они начинают выделять ионы калия и поглощать ионы натрия до тех пор,
Медицинская биофизика
161
пока не установится ионное равновесие. Процесс полностью обращается повышением температуры до 37 °С, На основе подобного рода опытов был сделан вывод, что перенос ионов калия и натрия против градиентов следует рассматривать как активный процесс, поскольку он совершается за счет энергии клеточного метаболизма.
При более детальном изучении переноса калия и натрия было установлено, что выход ионов натрия из клеток зависит от концентрации ионов калия во внешней среде а перенос калия в свою очередь зависит от концентрации натрия в цитоплазме. Во-вторых, обнаружено, что те вещества, которые уменьшают выход ионов натрия из клеток, оказывают такое же влияние и на поглощение ионов калия. При этом уменьшение переноса калия не является простым электрическим следствием уменьшения переноса натрия, поскольку мембранный потенциал при этом не изменяется. Эти наблюдения наряду с другими позволили сформулировать гипотезу сопряженного натрий-калиевого насоса, которой в настоящее время придерживаются большинство крупных ученых (Ходжкин, Катц, Шоу, Глинн, Кейнес и др.).
Предполагают, что перенос натрия и калия осуществляется специальным переносчиком белковой или белковолипидной природы. Переносчик на внутренней поверхности мембраны захватывает из цитоплазмы ионы натрия и переносит их на другую поверхность. На наружной поверхности мембраны ионы натрия отщепляются, а к переносчику присоединяются ионы калия, с которыми он движется снова к внутренней поверхности мембраны.
Для описанного транспорта ионов против градиентов необходимо поступление энергии АТФ, реализация которой происходит при действии фермента аденозинтрифосфатазы (АТФ-азы). Из всех АТФ-аз, имеющихся в клетках, решающее значение для транспорта ионов натрия и калия имеет АТФ-аза, активируемая этими же ионами (Nа+ — К+ — АТФ-аза). Na+ — К+— АТФ-аза была открыта Скоу (1954) в мембранных фракциях нервных клеток. Впоследствии этот фермент был обнаружен во многих других клетках. Свое предположение об участии Na+— К+ — АТФ-азы в переносе ионов Скоу подтвердил тем, что обнаружил инактивацию этого фермента сердечными гликозидами (оуабаином, строфантинами), которые тормозят перенос ионов.
162
В настоящее время факт локализации молекулярного механизма активного переноса ионов, а также Na+—К+—АТФ-азы в структуре самой мембраны не вызывает сомнения. Решающую роль в этом сыграли эксперименты с «тенями» эритроцитов и с заменой цитоплазмы клеток растворами различных электролитов. Так, из аксона кальмара (диаметром 0,5 мм) можно удалить всю аксоплазму и заполнить его вновь раствором электролитов с добавкой АТФ. На таком препарате, в котором сохранена лишь мембрана и отсутствует эктоплазма, удается наблюдать перенос ионов против градиентов концентрации и регистрировать потенциалы покоя и распространяющиеся потенциалы действия (см. также с. 189).
В нервной и почечной тканях, как установлено экспериментально, весь процесс переноса ионов протекает в три стадии: киназную, ионообменную и фосфатазную.
Первая стадия активируется ионами натрия. В этой стадии происходит фосфорилирование белка-переносчика и присоединение к нему ионов натрия:
АТФ + белок + Na+ <===> Na+ — белок ~ Ф + АДФ.
Возможно, что белок-переносчик и Nа+ — К+ — АТФ-аза — одно и то же вещество, подобно тому как актомиозин является и сократительным комплексом мышц и АТФ-азой одновременно. Эта стадия заканчивается переносом ионов натрия на внешнюю поверхность мембраны. Каков молекулярный механизм этого переноса, пока не ясно. По предположению Опита и Чарнока, он заключается в конформационных изменениях белковых молекул при фосфорилировании и присоединении ионов, в результате чего центры связывания ионов оказываются локализованными то с внешней, то с внутренней стороны мембраны. Как уже упоминалось в главе 5, подобный же механизм, по мнению Кеннеди, лежит в основе активного транспорта аминокислот и Сахаров. Эти предположения можно считать вероятными в связи с тем, что по данным ряда авторов (Василец, Нейфах и др.), из клеточных мембран удалось выделить сократительные белки, близкие к актомиозину мышц. Эти белки обладали Nа+ — К+ — АТФ-азными свойствами и чувствительностью к сердечным гликозидам.
Вторая стадия, протекающая на внешней поверхности мембраны и активируемая ионами калия, заклю-
II*
163
чается в обмене ионов натрия на ионы калия:

и в переносе последних через мембрану.
Цикл заканчивается на внутренней поверхности мембраны дефосфорилированием переносчика и освобождением ионов калия:

До настоящего времени не удалось выяснить один из важных вопросов при работе натрий-калиевого насоса: чем объясняется, что на внутренней поверхности мембраны переносчик обладает сродством к натрию, а на внешней — к калию.
Исследуя энергетику описанного процесса, Ходжкин показал, что энергия гидролиза одной молекулы АТФ может обеспечить перенос одного иона против разности электрохимического градиента приблизительно в 400 мВ. Такой перенос был бы экономичен три транспорте водородных ионов через мембраны клеток желудка, имеющих примерно соответствующую величину разности электрохимических потенциалов. Но в нервных клетках, имеющих разность электрохимических потенциалов всего 120 мВ, такой перенос был бы неэкономичен. Поэтому Ходжкин предположил, что при распаде одной молекулы АТФ должно переноситься 2 — 3 иона натрия. Впоследствии этот расчет был подтвержден многими исследователями для ряда тканей; было показано, что при гидролизе одного моля АТФ переносится 2 — 3 г-экв натрия.
Существуют данные, что натрий-калевый насос может работать в нескольких режимах. Он может переносить ионы калия и натрия в соотношениях 1:1, 1:2, 1: 3. Наконец, может наблюдаться перенос одних ионов натрия без переноса ионов калия. Было показано, что выход ионов натрия из клетки продолжается, хотя и с меньшей скоростью, после удаления из наружной среды ионов калия. Глинном и Постом была установлена точная стехиометрия ионного обмена в эритроцитах человека. Оказалось, что в этих клетках на каждые 2 поглощаемых иона калия выделяется 3 иона натрия.
До настоящего времени не было обнаружено явлений активного переноса анионов, в связи с чем считают, что
164
их распределение между клеткой и средой происходит пассивно1.
Кроме активного переноса минеральных ионов, существует активный перенос органических веществ, в частности Сахаров, аминокислот, нуклеотидов. Механизмы активного переноса органических веществ в настоящее время исследованы недостаточно.
Как и при активном транспорте ионов, при активном транспорте органических веществ перенос осуществляется против градиентов концентрации и требует затраты химической энергии клеток. В результате активного транспорта органических веществ наблюдается их неравномерное распределение между цитоплазмой и внешней средой. При ингибировании обменных процессов клетки неравномерность в распределении органических веществ уменьшается.
Имеются данные, что транспорт органических веществ также связан с Nа+—К+ — АТФ-азой. Было установлено, что транспорт органических веществ зависит от концентрации ионов натрия внутри клеток и угнетается строфантином, который, как указывалось, является ингибитором Na+ — К+ — АТФ-азы. Следовательно, транспорт органических веществ непосредственно сопряжен с транспортом ионов.
Все виды активного переноса веществ являются в то
же время и избирательным переносом, так как клетка
поглощает этим путем из окружающей среды только те
вещества, которые необходимы для ее жизнедеятель-
ности.
Как предполагает ряд исследователей (Рубинштейн, Драбкин, Уссинг, Пасынский), активный перенос необязательно предполагает участие активного переносчика типа белковых молекул, меняющих свою конформацию. Частично эти представления были рассмотрены выше при изложении сущности теории мембранных градиентов Д. Л. Рубинштейна.
Локализация некоторых ферментов в мембране позволяет несколько по другому объяснить активный перенос. Вещество А из внешней среды может на поверхности мембраны перейти в результате ферментативной реакции в какой-либо производное X (например, глюкоза в
1 Исключением из этого правила является, возможно, транспорт попав йода в клетках щитовидной железы.
165

Рис. 27. Модель активного переноса вещества.
A — исходное вещество; X — производное вещество; пунктирные стрелки диффузия; сплошные стрелки — ферментативное превращение вещества.
глюкозофосфат при участии АТФ и фермента гексокиназы). В результате этого концентрация вещества X около мембраны возрастает, а концентрация исходного вещества А уменьшается. Это приводит к тому, что производное вещество X начинает поступать по градиенту концентрации через мембрану в глубь клетки (рис. 27). С другой стороны, исходное вещество А также по градиенту концентрации движется из внешней среды к поверхности клетки. Внутри клетки производное X вновь превращается в исходное вещество А (например, глюкозофосфат в глюкозу под действием фермента фосфатазы), которое может накапливаться в клетке в сколь угодно больших количествах, поскольку (перенос осуществляется не по градиенту этого вещества, а по градиенту вещества X. Таким же образом вещество А может проникнуть через следующую мембрану и т.д. Если мембрана проницаема для вещества А, то оно будет диффундировать из клетки в среду, когда его концентрация в клетке превысит концентрацию в среде, т. е.. будет наблюдаться утечка вещества А из клетки.
Таким образом, активный перенос не означает, что вещество А в неизменном виде переносится против градиента его концентрации. Возможно, это энергия метаболических процессов как раз затрачивается на такие молекулярные превращения Л, при которых на каждом этапе переноса движение происходит по градиенту концентрации производных данного вещества, но суммарный баланс процесса переноса А (сахаров, аминокислот
166
и др.) соответствует передвижению против градиента концентрации.
Таким образом, между пассивным и активным транспортом веществ нет непереходимой грани. С одной стороны, пассивный транспорт происходит по градиентам, которые возникают в результате активного транспорта; с другой стороны, сам активный транспорт — противоградиентный процесс — сопряжен с градиентным процессом более высокого порядка — гидролизом АТФ. Здесь понятие «градиентный процесс» применяется в широком смысле и обозначен процесс, протекающий с уменьшением свободной энергии. Как уже указывалось в главе 2, противоградиентные процессы всегда имеют местный характер, поскольку они всегда сопряжены с другими — градиентными процессами. В конечном счете суммарный процесс в любой системе является градиентным и приводит к уменьшению свободной энергии системы и возрастанию энтропии. Убыль свободной энергии открытой системы компенсируется ее новым поступлением из окружающей среды; а в изолированной системе устанавливается термодинамическое равновесие. Окисление продуктов питания, а затем гидролиз АТФ с освобождением свободной энергии и являются теми градиентными процессами, которые обусловливают противоградиентиые (процессы в клетках, одним из которых является активный транспорт веществ.
Глава 7 БИОЭЛЕКТРИЧЕСКИЕ ПОТЕНЦИАЛЫ
Все процессы жизнедеятельности организмов сопровождаются появлением в клетках и тканях электродвижущих сил. Электрические явления играют большую роль в важнейших физиологических процессах: возбуждении клеток и проведении возбуждения по клеткам. Благодаря непосредственной связи биопотенциалов с метаболическими процессами и физологическим состоянием клеток они являются чувствительным и точно измеримым показателем различных изменений в клетках и норме и при патологии. Для более эффективного и
167
плодотворного использования электрофизиологических методов в медицине необходимо выяснение механизмов возникновения биоэлектрических потенциалов.

