




Биологическим объектам присущи пассивные электрические свойства: сопротивление и емкость (диэлектрическая проницаемость). Изучение пассивных электрических свойств биологических объектов имеет большое значение для понимания структуры и физико-химического состояния биологического вещества.
Биологические объекты обладают свойствами как проводников, так и диэлектриков. Наличие свободных ионов в клетках и тканях обусловливает проводимость этих объектов. Диэлектрические свойства биологических объектов и величина диэлектрической проницаемости определяются структурными компонентами и явлениями поляризации.
ЭЛЕКТРОПРОВОДНОСТЬ КЛЕТОК
и тканей для постоянного тока
|
|

| Сопротивление выражается формулой: |
|
|

Электропроводность L — это величина, обратная сопротивлению R проводника:
где ρ — удельное сопротивление; l — длина проводника; S — сечение проводника.
Сопротивление является коэффициентом пропорциональности между разностью потенциалов V и током I (закон Ома):

При пропускании постоянного тока через живые ткани было установлено, что сила тока не остается постоянной во времени, хотя прикладываемое напряжение не изменяется. Сила тока после наложения разности потенциалов начинает непрерывно уменьшаться и через некоторое время устанавливается на постоянном уровне. При этом она уменьшается в сотни и даже тысячи раз по сравнению с исходным значением (рис. 38, б).
14* 211
|
|

Получается как бы отклонение от закона Ома, согласно которому при постоянной разности потенциалов ток в проводнике тоже должен быть постоянным. Если бы в данном смысле биологический объект подчинялся закону Ома, то наш график был бы представлен прямой линией (см. рис. 38, а).
Уменьшение тока во
времени обусловлено явлениями поляризации, проходящими в ткани.
При прохождении постоянного тока через биологическую систему в ней возникает нарастающая. до некоторого предела ЭДС противоположного направления — ЭДС поляризации, которая уменьшает приложенную к объекту эффективную ЭДС, что и приводит к уменьшению тока. ЭДС поляризации Р(t) является функцией времени. Тогда закон Ома для биологического объекта следует записать:
 (4)
(4)
Возникновение ЭДС поляризации связанного способностью живых клеток накапливать заряды при прохождении через них тока,т. е. с емкостными, диэлектрическими свойствами биологических объектов, обусловленными явлениями поляризации.
ВИДЫ ПОЛЯРИЗАЦИЙ
Вещества обладают свободными и связанными зарядами. Свободные заряды — электроны и ионы — ион действием поля имеют возможность перемещаться от одного электрода к другому, создавая ток проводимости. Следует отметить, что в клетках свободные ноны могут перемещаться под действием поля в ограниченных объемах — от одной мембраны до другой. Связанные заряды под действием поля имеют возможность перемещаться только в некоторых, часто очень ограниченных пределах. При своем перемещении они создают токи смещения.
212
|
|

Процесс перемещения связанных зарядов под действием, электрического поля и образование вследствие этого электродвижущей силы, направленной против внешнего поля, называется поляризацией. Поляризация по своей природе делится на несколько видов.
| Рис. 39. Схема возникновения ди-польной (А) и макроструктурной {Б) поляризации вещества при наложении электрического поля. |
Электронная поляризация представляет собой смещение электронов на своих орбитах относительно положительно заряженных ядер в атомах и ионах. В результате такого смещения атом или ион превращается в индуцированный, наведенный диполь с направлением, противоположным внешнему полю. Время возникновения электронной поляризации после мгновенного наложения поля, называемое временем релаксации, равняется 10-16—10-14 с. Возникающий дипольный момент имеет небольшую величину.
Ионная поляризация — смещение иона относительно кристаллической решетки. Вследствие этого возникает дипольный момент с направлением, противоположным внешнему полю. Время релаксации ионной поляризации 10-14— 10-12 с.
Дипольная (ориентационная) поляризация. Если вещество содержит полярные молекулы и эти молекулы свободны, то под действием внешнего поля они ориентируются в соответствии с этим полем (рис. 39, А).
Дипольная поляризация имеет большое значение в веществах, молекулы которых обладают большим дипольным моментом (в воде, спиртах). Молекулы белков, а также других высокомолекулярных соединений вследствие диссоциации ионогенных групп, а также вследствие адсорбции ионов обладают значительными дипольными моментами. Поэтому в растворах данных
213
веществ дипольная поляризация, обусловленная вращением их полярных молекул, имеет большое значение.
Время возникновения дипольной поляризации — время релаксации — совпадает со временем поворота молекул. Время релаксации полярных молекул т зависит от вязкости среды ή, температуры Т, радиуса молекул г и вычисляется приближенно по формуле Стокса:
 (5)
(5)
где k — постоянная Больцмана.
Время релаксации дипольной поляризации изменяется в зависимости от указанных факторов в пределах от 10-13 до 10-7 с.
Макроструктурная поляризация возникает под действием электрического поля вследствие неоднородности электрических свойств вещества. Для ее возникновения необходимо наличие слоев с различной электропроводностью. Под действием поля свободные ионы и электроны, содержащиеся в проводящих субстанциях, перемещаются в пределах каждого включения до границы проводящего слоя. Дальнейшее перемещение свободных зарядов невозможно вследствие низкой проводимости соседних слоев (рис. 39, Б). В результате этого процесса проводящее включение приобретает дипольный момент и ведет себя подобно гигантской поляризационной молекуле. Время релаксации макроструктурной поляризации лежит в пределах 10-8—10-3 с.
Биологические объекты представляют собой гетерогенные структуры. Гетерогенность тканей в большой степени обусловлена наличием мембран. К ним относятся клеточные поверхностные мембраны и мембраны, окружающие клеточные органоиды и образующие эндо-плазматическую сеть. Если собственно цитоплазма клеток обладает малым сопротивлением в силу наличия в ней большого количества свободных ионов, то мембраны обладают очень большим сопротивлением (1000 Ом/см2) в результате их малой проницаемости для ионов. Макроструктурная поляризация происходит во всем объеме клеток, а не только на клеточной мембране, как считали раньше, поскольку гетерогенность структуры имеется во всем объеме клеток. За счет макроструктурной поляризации, которая играет основную роль в биологических объектах, диэлектрическая проницаемость тканей, изме-
214
ренная в постоянном электрическом поле, достигает очень больших величин — до нескольких миллионов.
Поверхностная поляризация происходит на поверхностях, имеющих двойной электрический слой. При наложении внешнего поля происходит перераспределение ионов диффузионной части двойного электрического слоя: частицы дисперсной фазы смещаются в одну сторону, а ионы диффузионного слоя — в другую. В результате этого частицы дисперсной фазы с противоионами диффузионного слоя превращаются в наведенные диполи. Время релаксации поверхностной поляризации лежит в пределах от 10-3 до 1 с.
Электролитическая поляризация возникает между электродами, опущенными в раствор электролита, при пропускании через них электрического тока. До протекания тока концентрация потенциалопределяющих ионов у обоих электродов была одинаковой, потенциалы электродов Е также были равны:
 (6)
(6)
где Е0 — нормальный потенциал электрода; R — газовая постоянная; Т —абсолютная температура; п — валентность ионов; F— число Фарадея; а — активность потенциалопределяющих ионов в приэлектродной зоне.
При наложении разности потенциалов на электроды произойдет перераспределение потенциалопределяющих ионов в диффузионной части двойного электрического слоя: в области катода концентрация ионов (катионов) увеличится, а в области анода — уменьшится. Обозначим активности ионов в приэлектродной зоне катода и анода через а1 и а2 соответственно. Тогда собственные потенциалы катода Е k и анода Е a станут равны:

и между электродами возникнет ЭДС поляризации Р(t), направленная против внешней ЭДС:
 (7)
(7)
В процессе дальнейшего изменения концентрации ионов в приэлектродных слоях могут начаться электрохимические реакции на электродах: ионы будут перехо-
215
дить из раствора на один электрод, а с другого электрода уходить в раствор. Поляризация в этом случае будет протекать уже по другому закону. Однако и в этом случае между электродами будет существовать ЭДС поляризации, направленная против приложенной извне ЭДС.
Таким образом, и в случае электролитической поляризации появление ЭДС поляризации обусловлено смещением зарядов, которое в данном случае проявляется как изменение концентрации ионов в приэлектродной зоне. ЭДС электролитической поляризации можно обнаружить следующим способом: отключить источник тока и измерить разность потенциалов между электродами прибором с высоким входным сопротивлением. По существу в данном случае электролитическая ячейка представляет собой концентрационный элемент. Время релаксации электролитической поляризации измеряется величинами порядка 10-4—102 с.
Все описанные явления поляризации в той или иной степени присущи биологическим объектам. При наложении внешней разности потенциалов в тканях возникает противоположно направленное электрическое поле, которое значительно уменьшает внешнее поле и обусловливает высокое удельное сопротивление тканей постоянному току (порядка 106—107 Ом-см). При этом вначале возникают те виды поляризации, которые имеют меньшее время релаксации.
Все явления поляризации могут быть описаны с помощью диэлектрической проницаемости вещества. Диэлектрическая проницаемость е характеризует уменьшение величины электрического поля в веществе по сравнению с величиной электрического поля в вакууме. Если напряженность однородного поля, образуемого некоторыми зарядами, в вакууме равняется Е 0, а напряженность поля, создаваемого этими же зарядами, в веществе — Е, то

Если, например, диэлектрическая проницаемость воды равняется 80, то это значит, что напряженность поля между двумя пластинами, между которыми находится вода, в 80 раз меньше напряженности поля, создаваемого этими же зарядами в вакууме. И это уменьшение
216
напряженности поля обусловлено дипольной поляризацией, вызванной вращением (ориентацией) полярных молекул воды в электрическом поле.
Величина е определяется как отношение емкости конденсатора С, между обкладками которого находится данное вещество, к емкости С0 того же конденсатора в вакууме:

ЭЛЕКТРОПРОВОДНОСТЬ КЛЕТОК И ТКАНЕЙ ДЛЯ ПЕРЕМЕННОГО ТОКА
Более полную информацию о биологическом объекте можно получить при измерении его электропроводности на переменном токе, поэтому в настоящее время изучение электрических свойств биологических систем обычно производится на переменном токе.
Так как биологические системы способны накапливать электрические заряды при прохождении через них тока, то их электрические свойства недостаточно описывать с помощью омического сопротивления. Необходимо пользоваться также понятием электрической емкости.
Емкость является коэффициентом пропорциональности между зарядом и потенциалом и определяется как отношение изменения заряда Δq проводника к изменению его потенциала ΔFi:
 (10)
(10)
Электрическая емкость при равных условиях геометрии объекта определяется его диэлектрической проницаемостью, т. е. явлениями поляризации. Для плоского конденсатора она определяется по формуле:
 (11)
(11)
где S — площадь пластин; d — расстояние между ними.
Измеряемая емкость биологического объекта определяется поляризационной емкостью, которая возникает в момент прохождения тока. Поляризационная емкость
217
отражает отношение изменения заряда объекта к изменению его потенциала при прохождении переменного тока. Изменение заряда за время I

а изменение потенциала

тогда поляризационная емкость Ср будет определяться следующим уравнением:
 (12)
(12)
где I0 и It — начальное и конечное значения силы тока; I — ток мгновенный; R — сопротивление объекта.
К поляризационной емкости биологического объекта присоединяется значительная по величине статическая емкость мембран (1 мкФ/см2). Как следует из уравнения (12), величина поляризационной емкости зависит от времени действия поля и может на низких частотах превышать величину статической емкости. На более высоких частотах (порядка 10 кГц) статическая емкость на несколько порядков выше поляризационной. А так как эти емкости соединены последовательно, то на высоких частотах общая величина емкости определяется меньшей по величине поляризационной емкостью.
Электрическая модель биологического объекта может быть представлена в виде различных комбинаций
 емкостей и сопротивлений — в виде различных эквивалентных схем. Наиболее простыми являются эквивалентные схемы с I последовательным и параллельным соединением С и R (рис. 40).
емкостей и сопротивлений — в виде различных эквивалентных схем. Наиболее простыми являются эквивалентные схемы с I последовательным и параллельным соединением С и R (рис. 40).
Так как биологические объекты обладают как проводимостью, так и ем костью, то они будут ха-
218
рактеризоваться как активным, так и реактивным сопротивлением. Реактивное емкостное сопротивление Rx определяется по формуле:
 (13)
(13)
где со — круговая частота тока.
Суммарное сопротивление объектов называется импедансом. Для последовательного соединения С и R импеданс определяется по формуле:
 (14)
(14)
для параллельного — по формуле:
 (15)
(15)
гдеZ— импеданс; i — мнимая единица, т. е. 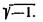
Из формул (14) и (15) следует, что импеданс объектов изменяется при изменении частоты тока, на котором производится измерение: при увеличении частоты реактивная составляющая импеданса уменьшается. Частотнозависимый характер емкостного сопротивления является одной из причин зависимости импеданса биологических объектов от частоты тока, т. е. дисперсии импеданса.
Изменение импеданса с частотой обусловлено также зависимостью поляризации от периода действия переменного тока. Если время, в течение которого электрическое поле направлено в одну сторону, больше времени релаксации какого-либо вида поляризации, то поляризация достигает своего максимального значения и вещество будет характеризоваться определенным постоянным значением эффективной диэлектрической проницаемости и проводимости. До тех пор пока полупериод переменного тока больше времени релаксации, эффективная диэлектрическая проницаемость и проводимость объекта не будут изменяться с частотой. Если же при увеличении частоты полупериод переменного тока становится меньше времени релаксации, то поляризация не успевает достигнуть максимального значения. После этого диэлектрическая проницаемость начинает уменьшаться с частотой, а проводимость — возрастать. При значительном увеличении частоты данный вид поляри-
219
|
|

| Рис. 41. Принципиальная схема моста для определения емкости и сопротивления биологических объектов. |
| R — омическое сопротивление уравновешенных плеч моста; Cxи Rx— параметры объекта; Cn и Rn—емкость и сопротивление, компенсирующие параметры объекта; U — измерительный прибор — индикатор баланса моста. |
зации практически будет от- сутствовать и диэлектриче- ская проницаемость и про- водимость снова станут по- стоянными величинами. Они будут определяться другими видами поляризации с меньшим временем релаксации. Электрические свойства тканей и клеток изучают с помощью мостовых схем. На рис. 41 изображена принципиальная схема моста переменного тока, применяемого для измерения емкости и сопротивления биологических объектов.
Обычно емкость и сопро
тивление тканей измеряют с
помощью металлических
электродов. При этом на
электродах происходят электрохимические реакции, ко
торые эквивалентны включению последовательно с объ
ектом дополнительного импеданса, что искажает ре
зультаты измерения. При использовании платиновых
электродов на низких частотах электродная емкость
составляет 20—50% от измеренной величины, а на высо
ких—10—25%. Снижение электродной поляризации до
стигается увеличением эффективной поверхности элек
тродов путем покрытия их платиновой чернью; поверх
ность образующегося при этом губчатого слоя в.
несколько сотен раз больше геометрической поверхности
электрода. При этом и величина электродного импедан-
са уменьшается в сотни раз. В последнее время разра-
ботано и несколько аналитических методов, позволяю-
щих учитывать электродную поляризацию.
Электродная поляризация должна учитываться так- же при регистрации биопотенциалов и при электрофорезе. В этих случаях для ее уменьшения часто пользуются так называемыми неполяризующимися электродами. Неполяризующийся электрод может быть представ- лен в виде металлического стержня, опущенного в раствор соли этого же металла. За счет обратимой химической реакции через соединение металла с электролитом
220
|
|

осуществляется непрерывное прохождение тока и поляризация не достигает большой величины. Электроды соединяются с объектом обычно агар-агаровыми солевыми мостиками и пипетками.
При изучении частотных зависимостей сопротивления и емкости биологических объектов было обнаружено три области дисперсии. На рис. 42 показана частотная зависимость диэлектрической проницаемости мышечной ткани. С увеличением частоты е уменьшается, образуя три зоны дисперсии: а, (3 и у.
α-Дисперсия занимает область низких частот звукового диапазона, примерно до 1 кГц" В данной области уменьшение диэлектрической проницаемости биологических систем обусловлено только уменьшением эффекта поляризации поверхности клеток, поскольку электрический ток с частотой до 1 кГц протекает практически только по межклеточникам, так как сопротивление клеток для токов низкой частоты велико. Как гомогенные, так и окруженные мембранами частицы обнаруживают одинаковые свойства на данных частотах. α-Дисперсия была получена для стеклянных частиц, полистироловых сфер и жировых частиц, взвешенных в электролите и обладающих двойным электрическим слоем. Это убедительно показывает, что наблюдаемая дисперсия обусловлена свойствами поверхности всей частицы.
По мнению Швана, Коула и некоторых других, явления поляризации, происходящие на биологических поверхностях, аналогичны явлениям поляризации на электродах: в обоих случаях поляризация возникает за счет перераспределения ионов в двойном электрическом слое. В таком случае поверхностная поляризация по своей сущности сводится к электролитической поляризации.
По мере увеличения частоты переменного тока эффект поверхностной поляризации уменьшается, что проявляется как уменьшение диэлектрической проницаемости и сопротивления ткани.
22 1
β-Дисперсия занимает более широкую область частот: 103—107 Гц. В прошлом для объяснения дисперсии диэлектрической проницаемости и сопротивления в данной области частот часто обращались к теории дипольной поляризации. Согласно этой теории, крупные молекулы органических веществ (белков, нуклеиновых кислот и др.) обладают дипольными моментами большой величины. Ориентация дипольных молекул под действием электрического поля обусловливает большие значения диэлектрической проницаемости в данной области частот. При увеличении частоты тока диполи не успевают поворачиваться вслед за полем, что приводит к уменьшению ε. При высокой частоте дипольная поляризация совсем не будет иметь места и диэлектрическая проницаемость снова станет стабильной.
При развитии теории дипольной поляризации предполагалось, что основная часть ионов в цитоплазме находится в связанном состоянии, поэтому ионные механизмы поляризации существенного значения не имеют. Теория дипольной поляризации тканей и клеток соответствовала фазовой теории в объяснении механизмов возникновения биопотенциалов.
Однако дальнейшее применение этой теории сопряжено с серьезными затруднениями. В частности, было обнаружено, что основная часть ионов цитоплазмы находится в свободном состоянии и, следовательно, может принимать участие в поляризации. Теория дипольной поляризации не может объяснить уменьшения емкости и сопротивления ткани при увеличении проницаемости клеточных мембран. При отмирании ткани ее электрические свойства приближаются к свойствам электролита, что также не находит удовлетворительного объяснения в дипольной теории. -
Наиболее полно электрические свойства биологических объектов в области β-дисперсии описываются теорией макроструктурной поляризации. Цитоплазма клеток представляет собой сложную систему с множеством границ раздела, создающих возможность для макро-
структурной поляризации. Согласно этой теории, емкость и проводимость биологических объектов в данной области частот определяются макроструктурной поляризацией, обусловленной гетерогенностью структуры — в первую очередь наличием мембран. Вначале считали, что явления поляризации обусловлены только клеточной
222
мембраной. Затем установили, что поляризация происходит во всем объеме клетки и вызывается наличием не только клеточной мембраны, но и мембран цитоплазмы. Последнее положение более справедливо для процессов поляризации, происходящих на относительно высоких частотах.
Однако теория макроструктурной поляризации в чистом виде не может описать всей сложности процессов поляризации в биологических системах. Теория дипольной, а также теория макроструктурной поляризации были перенесены из физики твердого тела и не учитывали биологических особенностей объектов, в частности клеточную проницаемость и зависимость электрических свойств клеток от величины ионных потоков (пассивных и активных) через мембраны.
В настоящее время развивается электрохимическая теория поляризации биологических объектов. Согласно этой теории, механизмы поляризации на границах раздела в биологических системах и на границе метал— раствор аналогичны. Данное направление дополняет теорию макроструктурной поляризации теорией электролитической поляризации в том смысле, что сущностью макроструктурной поляризации является перераспределение ионов на границе макроструктурного объема, т. е. электролитическая поляризация границы раздела.
Ценность данного подхода состоит в том, что он позволяет учитывать при описании электрических свойств тканей биологические особенности объекта —- клеточную проницаемость и наличие ионных потоков через мембрану.
Справедливость указанного подхода подтверждается экспериментальными данными. Так, при исследовании частотных зависимостей емкости модельной системы «электроды — раствор электролита» в области β-дисперсии было получено совпадение с частотной зависимостью емкости тканей. При определенных условиях было получено также совпадение температурных зависимостей электрических свойств тканей и указанной модельной системы.
Согласно данному представлению, биологический объект может быть представлен в виде эквивалентной схемы, изображенной на рис. 43. Поляризационные явления на границе раздела — на мембране — обусловливают возникновение поляризационного сопротивления
223
|
|

R Р и емкости Ср. R Р и СР зависят от величины результирующих ионных потоков через мембрану, т. е. от ее проницаемости, и статической емкости мембраны. Возможность оценки влияния клеточной проницаемости на электрические свойства биологических объектов и составляет, как указывалось, основную ценность данной аналогии с электролитической поляризацией на электродах. При этом аналогом электрохимической реакции на электродах является проникновение ионов через мембрану, а аналогом величины результирующего ионного потока через мембрану служит ток обмена, протекающий через поверхность электрода и зависящей от константы скорости электрохимической реакции.
Последовательно с R Р и Ср включено сопротивление содержимого клеток Ri— сопротивление собственно цитоплазмы. При условии, что основная часть ионов в клетке находится в свободном состоянии, можно считать Riсопротивлением электролита. Емкость, поляризационное сопротивление и сопротивление цитоплазмы шунтируются параллельно включенным сопротивлением межклеточной жидкости R м.
В данной схеме Ср и R Р являются частотнозависимыми элементами, а Ri н R м (сопротивления электролитов) — частотнонезависимыми элементами. Несложный анализ рассматриваемой схемы показывает, что при измерении на низких частотах в случае большой величины межклеточных промежутков объекта общее измеряемое сопротивление будет обусловлено в основном сопротивлением межклеточников, которое на несколько порядков ниже поляризационного сопротивления. Если же межклеточные промежутки имеют малую величину, то измеряемое сопротивление будет обусловлено в основном поляризационным сопротивлением R Р. А так как поляризационное сопротивление зависит от проницаемо-
224
стй мембран, то низкочастотное сопротивление является в определенных условиях мерой проницаемости мембран биологической системы.
При увеличении частоты время протекания тока в одном направлении уменьшается. Это приводит к меньшему перераспределению ионов на границах раздела и к уменьшению эффекта поляризации, что проявляется уменьшением R р, Ср и эффективной диэлектрической проницаемости ткани. Таким образом, β-дисперсия обусловлена уменьшением эффекта поляризации границ раздела в биологических системах. Это не исключает и влияния дипольной поляризации, которая вносит небольшой вклад в общие величины электрических параметров.
На достаточно высоких частотах, порядка нескольких мегагерц, поляризация границ раздела — мембран — будет отсутствовать совсем, и электрические параметры ткани станут стабильными. На этих частотах R р и Ср можно исключить из эквивалентной схемы, и высокочастотное сопротивление будет представлено сопротивлениями электролитов: Riи R м. Поскольку сопротивление электролитов определяется концентрацией свободных ионов, то высокочастотное сопротивление биологических объектов является показателем содержания в них свободных ионов.
γ-Дисперсия диэлектрической проницаемости тканей наблюдается на частотах выше 1000 МГц. Уменьшение диэлектрической проницаемости в данном диапазоне обусловлено ослаблением эффекта поляризации, вызываемой диполями воды. Величина γ-дисперсии будет зависеть от содержания свободной воды в исследуемых тканях. В области 400 МГц (между β- и γ-дисперсией) величина е для тканей (кроме жировой, костной и мозговой) лежит в пределах 40—60 в зависимости от содержания свободной воды.
В области сверхвысоких частот (больше 1010 Гц) эффект поляризации, обусловленный диполями воды, будет отсутствовать. Диэлектрическая проницаемость будет иметь небольшие значения, определяемые ионной и электронной поляризацией, имеющей самое малое время релаксации. На три основных дисперсионных явления, обусловленных поверхностной поляризацией, макроструктурной поляризацией и дипольной поляризацией воды, накладываются дополнительные, сравнительно меньшие эффекты поляризации:
15 Медицинская биофизика
225
1) поляризация белковых и других органических молекул, обладающих дипольными моментами. Дисперсия данного вида поляризации происходит на частотах в несколько мегагерц;
2) поляризация гидратных оболочек макромолекул (связанной воды). Дисперсия наблюдается в диапазоне частот 100—1000 МГц;
3) поляризация связанных полярных групп макро
молекул, имеющих меньшее время релаксации, чем макродиполи молекул. Предполагается, что область дисперсии этого вида поляризации 100—1000 МГц.
Общая картина частотной зависимости электрических параметров сохраняется для всех тканей, что обусловлено единством структуры и химического состава клеток. В каждом конкретном случае индивидуальные особенности объектов обусловливают характер частотной зависимости. К таким особенностям относятся размеры и форма клеток, величина их проницаемости, соотношение между объемом клеток и межклеточных пространств, содержание свободных ионов в клетках, содержание свободной воды и пр.

