




Несмотря на деятельность шаперонов и шаперонинов в клетке регулярно возникают «испорченные» белковые молекулы, исправить которые шаперонинам не удается. Кроме того, многие белки, особенно регуляторные, выполнив свою функцию, становятся ненужными. Такие молекулы должны быть уничтожены. Для этого существуют специальные ферменты, разрушающие белки – протеазы. Но как клетке удается разрушить только определенные белковые молекулы, не затронув все  остальные? Для этого существуют специальные механизмы. К молекуле, предназначенной для уничтожения, присоединяется специальная «метка» – цепочка, состоящая из нескольких молекул небольшого белка убиквитина (рис. 27[O27], Б). Ученые назвали это событие «поцелуем смерти». Помеченный белок попадает в протеасому – белковый «стаканчик», похожий на шаперонин (рис. 27, А, В), но предназначенный не для исправления, а для уничтожения молекулы. В протеасоме белок расщепляется на аминокислоты, которые могут быть использованы для построения новых белков.
остальные? Для этого существуют специальные механизмы. К молекуле, предназначенной для уничтожения, присоединяется специальная «метка» – цепочка, состоящая из нескольких молекул небольшого белка убиквитина (рис. 27[O27], Б). Ученые назвали это событие «поцелуем смерти». Помеченный белок попадает в протеасому – белковый «стаканчик», похожий на шаперонин (рис. 27, А, В), но предназначенный не для исправления, а для уничтожения молекулы. В протеасоме белок расщепляется на аминокислоты, которые могут быть использованы для построения новых белков.
Посттрансляционная модификация белков. Многие белки выполняют свои функции в клетке не «поодиночке», а будучи соединенными с другими молекулами – липидами (липопротеиды), углеводами (гликопротеиды) или образовав макромолекулярный комплекс с другими белками. Как правило, присоединение этих молекул происходит в аппарате Гольджи под действием специальных ферментов.
Посттрансляционная модификация белков очень широко используется клеткой для регуляции активности белков. Например, обратимое фосфорилирование-дефосфорилирование «включает» и «выключает» многие регуляторные белки, такие как транскрипционные факторы. Включение активности некоторых белков происходит в результате отщепления определенных участков молекулы. Таким способом активируются белковый гормон инсулин, образующийся из проинсулина, фактор свертывания крови тромбин, образующийся из протромбина и некоторые другие белки. В настоящее время известно более 100 способов посттрансляционной модификации белков.
§ 11. Регуляция синтеза белка.
Как мы уже знаем, все процессы жизнедеятельности любого организма определяются тем, какие белки, когда и в каком количестве производятся клетками. Все эти параметры постоянно меняются в зависимости от потребностей организма. Это означает, что все процессы, связанные с синтезом белков, должны регулироваться. Они и регулируются, причем на разных уровнях – на уровне транскрипции, на уровне трансляции, на уровне посттрансляционной модификации и разрушения белков. Регуляторные системы клетки очень сложны и разнообразны. Мы познакомимся только с некоторыми.
Регуляция синтеза белка у прокариот. У бактерий гены ферментов, участвующих в одной цепи биохимических реакций, объединены в один оперон, имеющий один промотор. Многие гены являются конститутивными, то есть постоянно находятся в активном состоянии, транскрибируются и транслируются. Например, у бактерии E. coli ферменты метаболизма (обмена) глюкозы кодируются генами, находящимися в нескольких постоянно транскрибируемых оперонах. Однако количество мРНК, считанных с разных оперонов, может быть различным. Это зависит от «силы» промоторов. К одним промоторам сродство РНК-полимеразы может быть бóльшим, чем к другим, с ними за единицу времени связывается и начинает транскрипцию большее количество холофермента.
|
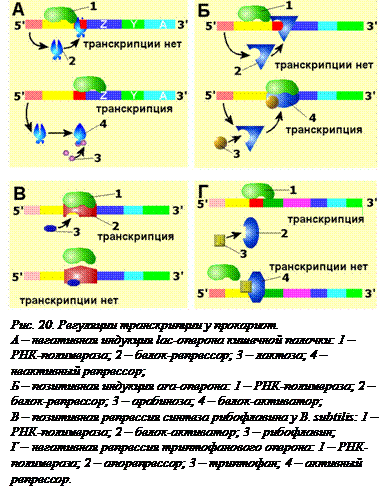 экономично регулировать синтез ферментов в самом начале пути – во время считывания информации с ДНК, то есть транскрипции. Работа оперонов регулируется путем индукции (включения) или репрессии (подавления) транскрипции, которые могут быть как негативной (с участием «выключателя»), так и позитивной, с участием «включателя».
экономично регулировать синтез ферментов в самом начале пути – во время считывания информации с ДНК, то есть транскрипции. Работа оперонов регулируется путем индукции (включения) или репрессии (подавления) транскрипции, которые могут быть как негативной (с участием «выключателя»), так и позитивной, с участием «включателя».Негативная индукция. Так называемый l ac -оперон E. coli содержит 3 гена, отвечающие за образование белков, участвующих в переносе в клетку дисахарида лактозы и в ее расщеплении. Белок-репрессор, регулирующий работу lac -оперона, кодируется в гене другого оперона, не имеющего оператора. Поэтому он постоянно образуется в клетке. В отсутствие субстрата (лактозы), lac -оперон выключен (рис. 20, А). Только наличие лактозы инактивирует белок-репрессор, и lac -оперон начинает работать. Как только лактоза появляется в среде и попадает в клетку, две ее молекулы соединяются с белком-репрессором, изменяют его конформацию, и он отсоединяется от оператора. Тут же начинается транскрипция lac -оперона и трансляция образующейся мРНК; три разных синтезируемых белка участвуют в утилизации лактозы. Когда вся лактоза переработана, очередная порция репрессора, свободного от лактозы, выключает lac -оперон. Эта схема называется негативной, потому что контролирующим транскрипцию фактором является негативный фактор, «выключатель» – белок-репрессор. Индукция происходит при потере сродства белка репрессора к оператору.
Другой оперон кишечной палочки, также содержащий три цистрона, кодирующие разные ферменты, расщепляющие сахар арабинозу, регулируется с помощью позитивной индукции (рис. 20, Б). Обычно ara -оперон не работает, так как белок-репрессор связан с оператором. Взаимодействие попавшей в клетку арабинозы с белком-репрессором превращает его в активатор, облегчающий посадку РНК-полимеразы на промотор. Эта схема регуляции называется позитивной индукцией, поскольку контролирующий элемент – белок-активатор «включает» работу оперона.
Третий тип регуляции – позитивную репрессию, рассмотрим на примере регуляции транскрипции оперона синтеза рибофлавина у другой бактерии – Вacilus subtilis. В опероне располагаются цистроны ферментов синтеза рибофлавина. Позитивная репрессия так называется потому, что в ней участвует белок-активатор, а сама регуляция заключается в его инактивации. Она противоположна позитивной индукции. Существует белок-активатор, обеспечивающий посадку РНК-полимеразы на промотор. Обычно оперон открыт, так как рибофлавин нужен постоянно. Но синтезироваться должно столько молекул, сколько в данный момент нужно клетке. Лишняя молекула рибофлавина взаимодействует с белком-активатором, и он теряет способность обеспечивать посадку РНК-полимеразы на промотор (рис. 20, В).
Негативная репрессия используется клетками кишечной палочки для регуляции работы оперона, отвечающего за синтез аминокислоты триптофана. Негативная репрессия так называется потому, что есть белок-репрессор, а сама регуляция заключается в «выключении» работающего оперона. В этом опероне имеется 5 цистронов, которые кодируют ферменты последовательной цепи реакций синтеза триптофана. Обычно оперон включен. Белок-репрессор есть, но он неактивен, так как находится в форме так называемого апорепрессора. В этой форме он не способен садиться на оператор. Клетке нужно определенное количество молекул триптофана. Лишняя молекула взаимодействует с апорепрессором. Он меняет конформацию, связывается с оператором, и синтез РНК прекращается (рис. 20, Г).
 Одновременная транскрипция и трансляция. В отличие от эукариот, у которых транскрипция происходит в ядре, а трансляция – в цитоплазме, у прокариот оба процесса происходят в одном и том же месте. Как уже говорилось, прокариоты должны оперативно реагировать на любые изменения окружающей среды, в первую очередь на появление или исчезновение тех или иных питательных веществ. Поэтому когда определенный оперон включается, то есть начинается транскрипция, то клетка не ждет, пока молекула мРНК будет полностью синтезирована, а начинает трансляцию сразу же, как только размер синтезированного участка будет достаточен для посадки рибосомы. По мере продвижения РНК-полимеразы по ДНК и удлинения молекулы мРНК рибосома также продвигается, освобождая начало молекулы, куда может сесть новая рибосома. Чем дальше продвигается полимераза по оперону, тем длиннее оказывается молекула РНК, тем больше рибосом, ведущих трансляцию оказывается на ней. Такая структура – нить мРНК и много (иногда несколько десятков) рибосом на ней называется полисомой. Каждый оперон одновременно транскрибируется несколькими одинаковыми РНК-полимеразами. Из каждой молекулы фермента «свисает» полисома, которая тем длиннее, что дальше продвинулась РНК-полимераза (рис. 21).
Одновременная транскрипция и трансляция. В отличие от эукариот, у которых транскрипция происходит в ядре, а трансляция – в цитоплазме, у прокариот оба процесса происходят в одном и том же месте. Как уже говорилось, прокариоты должны оперативно реагировать на любые изменения окружающей среды, в первую очередь на появление или исчезновение тех или иных питательных веществ. Поэтому когда определенный оперон включается, то есть начинается транскрипция, то клетка не ждет, пока молекула мРНК будет полностью синтезирована, а начинает трансляцию сразу же, как только размер синтезированного участка будет достаточен для посадки рибосомы. По мере продвижения РНК-полимеразы по ДНК и удлинения молекулы мРНК рибосома также продвигается, освобождая начало молекулы, куда может сесть новая рибосома. Чем дальше продвигается полимераза по оперону, тем длиннее оказывается молекула РНК, тем больше рибосом, ведущих трансляцию оказывается на ней. Такая структура – нить мРНК и много (иногда несколько десятков) рибосом на ней называется полисомой. Каждый оперон одновременно транскрибируется несколькими одинаковыми РНК-полимеразами. Из каждой молекулы фермента «свисает» полисома, которая тем длиннее, что дальше продвинулась РНК-полимераза (рис. 21).
Для того, чтобы синтез белка прекратился, когда потребность в нем минует, прекращения транскрипции недостаточно, должна быть ликвидирована матрица мРНК, чтобы рибосомы не имели возможности продолжать трансляцию. Другими словами, мРНК должна не только синтезироваться в клетке, но и разрушаться– ведь без баланса процессов синтеза и разрушения молекул невозможна тонкая регулировка процессов жизнедеятельности. Разрушение РНК происходит в результате действия РНКазы, фермента, «откусывающего» по одному нуклеотиду с 5’-конца. Пока конец занят рибосомой, мРНК недоступна для действия фермента, но если не происходит посадки новой рибосомы на освободившийся конец, он начинает разрушаться РНКазой.
В результате оказывается, что нередко РНК-полимераза еще не дошла до терминатора в опероне, рибосомы еще не закончили синтез закодированного в гене белка, а части мРНК, соответствующей началу гена, уже нет. Это позволяет синтезировать столько молекул белка, сколько нужно клетке, не тратя ресурсы на производство лишних молекул. Таким образом, транскрипция, трансляция и разрушение мРНК у прокариот не разобщены во времени и пространстве. Это и позволяет им быстро реагировать на изменяющиеся условия среды.
Регуляция транскрипции у эукариот. Операторов в эукариотических генах нет, а регуляция генов, кодирующих белки, осуществляется особыми элементами – энхансерами. Энхансеры – это последовательности нуклеотидов в ДНК, усиливающие транскрипцию при взаимодействии со специфическими белками. Энхансеры не представляют собой непрерывных последовательностей нуклеотидов. Существуют так называемые модули – отдельные части энхансеров. Одинаковые модули могут встречаться в разных энхансерах, но для каждого энхансера набор модулей уникален. Модули – это короткие последовательности, не более 2-х витков спирали (20 п.н.), которые могут находиться перед, за и даже внутри гена (рис 22, А). Отдельные модули могут располагаться на расстоянии нескольких тысяч пар нуклеотидов от 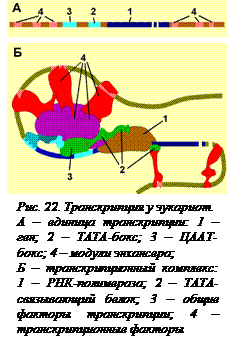 регулируемого гена. С каждым модулем связывается специфический белок, называемый транскрипционным фактором. Эти белки имеют сродство друг к другу и к основным белковым факторам транскрипции. Сближаясь при изгибании ДНК, они активируют инициаторный комплекс (рис. 22, Б). Инициаторный комплекс – это агрегат РНК-полимеразы с основными общими белковыми факторами транскрипции. Для того чтобы транскрипция началась, все транскрипционные факторы должны занять свое место. Только тогда РНК-полимераза правильно устанавливается на промоторе и может начать работу. Кроме энхансеров существуют и сайленсеры – ослабители. Это нуклеотидные последовательности, с которыми связываются белки-репрессоры, которые не активируют, а подавляют образование инициаторного комплекса транскрипции.
регулируемого гена. С каждым модулем связывается специфический белок, называемый транскрипционным фактором. Эти белки имеют сродство друг к другу и к основным белковым факторам транскрипции. Сближаясь при изгибании ДНК, они активируют инициаторный комплекс (рис. 22, Б). Инициаторный комплекс – это агрегат РНК-полимеразы с основными общими белковыми факторами транскрипции. Для того чтобы транскрипция началась, все транскрипционные факторы должны занять свое место. Только тогда РНК-полимераза правильно устанавливается на промоторе и может начать работу. Кроме энхансеров существуют и сайленсеры – ослабители. Это нуклеотидные последовательности, с которыми связываются белки-репрессоры, которые не активируют, а подавляют образование инициаторного комплекса транскрипции.
Образование рибосом у эукариот. В некоторых случаях описанных способов регуляции синтеза белка оказывается недостаточно. Например, эукариотической клетке нужно огромное количество рибосом – сотни тысяч, а иногда и миллионы. Даже самая интенсивная транскрипция одного гена не может обеспечить такого количества молекул рРНК. Поэтому в данном случае используется другой «метод» – увеличение количества одинаковых генов в одной клетке. Гены рибосомных РНК присутствуют в количестве от 10 до 105 копий в клетках разных видов эукариот. Например, у человека в каждой клетке находится около 200 копий генов, в которых закодированы рРНК. Все рибосомные гены (кроме генов 5 S рибосомной РНК) расположены группами один за другим, а эти группы, называемые кластерами, многократно повторяются в хромосомах. В световой микроскоп можно видеть ядрышки – места синтеза и рибосомных РНК и образования субчастиц рибосом (рис. 23, А). Ядрышки представляют собой сближенные кластеры рибосомных генов, находящиеся в разных хромосомах. Плотная структура ядрышек образована большим количеством продуктов рибосомных генов – рРНК и готовых субчастиц рибосом. Ядрышек в клетке может быть одно или несколько. Группа расположенных рядом генов рРНК называется ядрышковым организатором.
 На электронно-микроскопических фотографиях транскрибирующихся генов рибосомной РНК видна картина, напоминающая «рождественскую елочку» (рис. 23, Б). «Ствол елочки» – это молекула ДНК. РНК-полимераза движется от «верхушки елочки» к ее основанию. Каждая «ветка елочки» – это одна молекула транскрипта, синтезируемая одной РНК-полимеразой. Чем дальше от «вершины», тем длиннее «ветка».На «ветках» «висят» рибосомные белки. Для рибосомных белков существуют свои гены. Синтезируемые на них мРНК поступают на готовые рибосомы в цитоплазму, где синтезируются рибосомные белки для новых рибосом. Эти вновь синтезированные рибосомные белки поступают в ядро и «запутываясь в ветвях елки», соединяются с образующейся рРНК. Так формируются рибосомные субчастицы. Одновременно в эукариотическом ядре находятся тысячи субчастиц рибосом.
На электронно-микроскопических фотографиях транскрибирующихся генов рибосомной РНК видна картина, напоминающая «рождественскую елочку» (рис. 23, Б). «Ствол елочки» – это молекула ДНК. РНК-полимераза движется от «верхушки елочки» к ее основанию. Каждая «ветка елочки» – это одна молекула транскрипта, синтезируемая одной РНК-полимеразой. Чем дальше от «вершины», тем длиннее «ветка».На «ветках» «висят» рибосомные белки. Для рибосомных белков существуют свои гены. Синтезируемые на них мРНК поступают на готовые рибосомы в цитоплазму, где синтезируются рибосомные белки для новых рибосом. Эти вновь синтезированные рибосомные белки поступают в ядро и «запутываясь в ветвях елки», соединяются с образующейся рРНК. Так формируются рибосомные субчастицы. Одновременно в эукариотическом ядре находятся тысячи субчастиц рибосом.
§ 12. Процессинг РНК.
Процессинг у прокариот. Многие синтезированные в ходе транскрипции молекулы РНК оказываются еще не готовыми к выполнению своих функций. Они должны еще «созреть». Это созревание называется процессингом. У прокариот процессингу подвергаются предшественники рРНК и тРНК. Они синтезируются в виде больших молекул, так называемых первичных транскриптов, из которых специальные ферменты вырезают куски, которые затем «подстригаются» до зрелых молекул рРНК и тРНК. Матричные РНК прокариот процессингу не подвергаются. Рибосомы синтезируют отдельные белки по единой полицистронной молекуле мРНК.
Процессинг у эукариот. Процессинг рибосомных РНК у эукариот принципиально не отличается от такового у прокариот и заключается в том, что из первичного транскрипта – гигантской молекулы так называемой 45 S -РНК ферменты «выкраивают» зрелые молекулы рРНК. Рибосомные белки, поступающие в ядро, соединяются с рРНК, входящей в состав еще только растущего первичного транскрипта, а разрезание его происходит после того, как транскрипция закончится.
Процессинг мРНК у эукариот очень сложен и состоит из нескольких этапов – кепирования, полиаденилирования, сплайсинга и редактирования. Все стадии процессинга мРНК проходят в РНП-частицах (рибонуклеопротеидных комплексах). По мере синтеза про-мРНК, она сразу же образует комплексы с ядерными белками – информоферами. Поэтому мРНК никогда не бывает свободной от белков. На всем пути следования от ДНК, по которой она синтезируется, до завершения трансляции на рибосоме мРНК защищена от нуклеаз. Кроме того, белки придают ей необходимую конформацию. И в ядерные, и в цитоплазматические комплексы (информосомы) мРНК с белками входят так называемые малые РНК – s РНК (от англ. small – маленький). В составе информосом иРНК может жить от нескольких минут до нескольких дней, не подвергаясь действию нуклеаз.
Кепирование. Кепирование означает «надевание шапочки». Шапочка (англ. с ap) представляет собой метилированный ГТФ, присоединенный в необычной позиции 5'-5' и две метилированные рибозы в первых двух нуклеотидах мРНК. Кепируются мРНК еще до завершения транскрипции. По мере образования про-мРНК (еще до 30-ого нуклеотида), к 5'-концу, несущему пуринтрифосфат, присоединяется гуанин, после чего происходит метилирование. Кепируются все информационные РНК, синтезируемые в ядре клетки. «Шапочка» защищает 5'-конец мРНК от действия экзонуклеаз и помогает взаимодействию мРНК с рибосомой. За счет ее узнавания сар- связывающими белками происходит правильная установка мРНК на рибосоме. У прокариот за это отвечает последовательность Шайна-Дальгарно, которой у эукариот нет.
Полиаденилирование. В большинстве генов эукариот в конце гена находится так называемый сигнал полиаденилирования. В транскрибированной с гена мРНК он 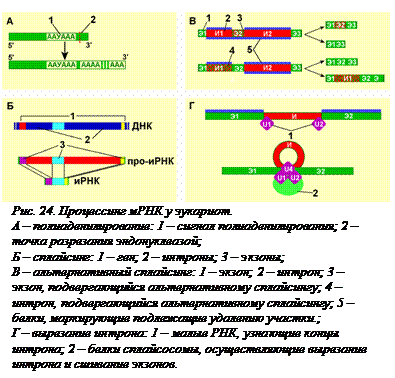 представляет собой последовательность нуклеотидов ААУААА. Когда синтез про-мРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3'-концу от последовательности 5'-AAУAAА-3' происходит разрезание молекулы специфической эндонуклеазой и к новому 3'-концу без использования матрицы присоединяется от 30 до 300 адениловых нуклеотидов (рис. 24, А).
представляет собой последовательность нуклеотидов ААУААА. Когда синтез про-мРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3'-концу от последовательности 5'-AAУAAА-3' происходит разрезание молекулы специфической эндонуклеазой и к новому 3'-концу без использования матрицы присоединяется от 30 до 300 адениловых нуклеотидов (рис. 24, А).
Каждый вид мРНК имеет «поли-А хвост» определенной длины. Он защищает 3'-конец от гидролиза, так как покрыт полиА-связывающими белками. В значительной степени время жизни мРНК коррелирует с длиной полиА-хвоста. Полиаденилируется более 95% мРНК. Информационные РНК некоторых генов не полиаденилируются. Такие мРНК живут несколько минут, например, гистоновые мРНК, тогда как полиаденилированные – несколько часов или даже дней.
Сплайсинг. Одним из важнейших отличий организации генетического материала у про- и эукариот является то, что большинство генов эукариот «разорваны», то естькодирующие участки генов – экзоны, разделяются некодирующими участками – интронами (подробнее об этом будет рассказано в § 18). Количество экзонов в гене всегда на один больше, чем интронов. Сплайсинг – это вырезание копий интронов из про-мРНК и сшивание копий экзонов с образованием мРНК (рис 24, Б). Вырезанные копии интронов гидролизуются до нуклеотидов. Сплайсингу подвергается большинство мРНК – примерно 95%. Кроме информационных РНК сплайсингу подвергаются и некоторые тРНК, а также рРНК у простейших.
Сплайсинг подавляющего большинства про-мРНК в ядрах высших эукариот осуществляется белковыми комплексами – сплайсосомами. Сплайсосома непосредственно связана с ферментами, занимающимися полиаденилированием, поэтому сплайсируются только полиаденилированные мРНК.
Некоторые гены эукариот, например, гены гистонов, не содержат интронов, их мРНК не полиаденилируются и не подвергаются сплайсингу.
Альтернативный сплайсинг. У высших организмов первичный РНК-транскрипт может подвергаться альтенативному сплайсингу. При этом некоторые из экзонов могут удаляться вместе с интронами, а некоторые интроны или только части интронов оставаться в зрелой мРНК и служить матрицей для части полипептидной цепочки (рис. 24, В). В результате по одному гену может образоваться несколько разных мРНК, по которым синтезируются разные белки. Именно альтернативным сплайсингом объясняется тот факт, что у человека синтезируется около 90·103 разных типов белков, кодируемых всего ~20·103 генами. Все гены человека, содержащие интроны, а это 94% генов, служат матрицами для синтеза про-мРНК, подвергаемых альтернативному сплайсингу. Уже описано более 10 белковых комплексов, называемых SR -белками (сплайсинг-регулирующми), которые различны в разных тканях и даже в одной ткани на разных этапах развития организма. SR-белки связываются с определенными нуклеотидными последовательностями внутри экзонов и сигнализируют о вырезании или сохранении конкретного экзона в мРНК.
В результате альтернативного сплайсинга по каждому гену высших эукариот может образовываться в среднем три разных мРНК, а по некоторым генам – даже несколько сотен.
Единого механизма сплайсинга не существует. Описано несколько разных механизмов. Обычно в сплайсинге помимо ферментов, вырезающих и сшивающих участки про-мРНК, участвуют белки, придающие про-мРНК нужную конформацию, и несколько s n PНК (малых ядерных РНК). Размер этих РНК – от 100 до 300 нуклеотидов. Поскольку в большинстве случаев эти РНК обогащены урацилом, они называются U1, U2... Они входят в состав рибонуклеопротеидов, участвующих в полиаденилированиии и сплайсинге. s n PНК узнают короткие последовательности нуклеотидов, маркирующие концы интронов, и сближают их друг с другом (рис. 24, Г). Благодаря этому разрезание и сшивание концов происходит очень точно, с точностью до одного нуклеотида. Это принципиально важно, так как при «собирании из кусков» не должно происходить сдвига рамки считывания.
В митохондриях сплайсинг осуществляют специальные ферменты- матуразы. Как оказалось, они кодируются в интронах (вернее, часть белка кодируется интроном, а часть – прилежащим участком экзона) генов, мРНК которых подвергается сплайсингу! Синтезированный фермент осуществляет сплайсинг, вырезая интрон и тем самым уничтожая собственную матрицу. Это предотвращает синтез лишних молекул фермента.
Особый способ сплайсинга, так называемый автосплайсинг, был открыт Томасом Чеком (США) в 1982 году. Он работал с инфузорией Tetrаchymenа thermophyla. У этой инфузории образуется 35 S про-рРНК длиной 6400 нуклеотидов. Из этой про-рРНК вырезается внутренний участок длиной в 414 нуклеотида, два экзона сшиваются с образованием 26 S рРНК. Оказалось, что это происходит без какого-либо участия соединений белковой природы. Эта про-рРНК имеет особую третичную структуру и обладает каталитической активностью. Так впервые было показано, что каталитической активностью обладают не только белки, но и РНК. РНК с каталитической активностью назвали рибозимами по аналогии с белковыми ферментами, которые называют также энзимами. В настоящее время описано несколько десятков рибозимов.
Редактирование. [O28] Редактирование – это изменение генетической информации на уровне мРНК. Открыто это явление было при изучении экспрессии генов трипаносомы.
Трипаносома – одноклеточный паразит, вызывающий тяжёлые, часто смертельные заболевания человека и животных. У трипаносом сложный путь индивидульного развития, включающий смену хозяев – теплокровных (млекопитающих и птиц) и насекомых. В клетке трипаносомы есть митохондрии и ферменты окислительного фосфорилирования. Один из ключевых ферментов этого процесса – цитохромоксидаза. Это белок с четвертичной структурой, состоящий из трех разных субъединиц. Когда трипаносома живет в теплокровных млекопитающих, ей достаточно молекул АТФ, образующихся в процессе гликолиза. При этом синтезируются только две субъединицы цитохромоксидазы, и фермент не работает. Насекомые-переносчики, промежуточные хозяева трипаносомы, холоднокровны, поэтому трипаносома нуждается в окислительном фосфорилировании для получения большого количества молекул 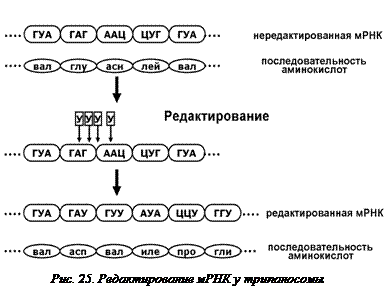 АТФ. Поэтому, когда трипаносома находится в организме насекомого, у нее синтезируются все три субъединицы фермента. В геноме трипаносомы есть только два гена для двух разных субъединиц. В мРНК одного из них происходит разрезание и встраивание четырех уридинов (на небольшом расстоянии друг от друга, но не подряд) (рис. 25). Происходит сдвиг рамки считывания, и отредактированная иРНК кодирует совершенно другой, новый полипептид – третью субъединицу цитохромоксидазы.
АТФ. Поэтому, когда трипаносома находится в организме насекомого, у нее синтезируются все три субъединицы фермента. В геноме трипаносомы есть только два гена для двух разных субъединиц. В мРНК одного из них происходит разрезание и встраивание четырех уридинов (на небольшом расстоянии друг от друга, но не подряд) (рис. 25). Происходит сдвиг рамки считывания, и отредактированная иРНК кодирует совершенно другой, новый полипептид – третью субъединицу цитохромоксидазы.
Еще несколько лет назад редактирование мРНК считалось «экзотикой», свойственной только трипаносомам. Сейчас установлено, что редактирование широко распространено среди эукариот. Редактируются не только мРНК, но и некоторые регуляторные малые РНК.
Регуляторная роль РНК.
В течение примерно полувека считалось, что РНК в клетке играет только подсобную роль посредника между ДНК и белком. Однако результаты, полученные исследователями на рубеже тысячелетий, показали, что РНК – одно из самых главных «действующих лиц» в клетке. Она не только переносит информацию от ДНК к белку, но и непосредственно участвует в его синтезе. Кроме того, как оказалось, существуют особые РНК-регуляторы разных сторон жизни клетки.
РНК-переключатели. В 2002 году была открыта регуляция транскрипции у прокариот без участия белков-репрессоров или активаторов. Она осуществляется с помощью особых участков уже транскрибированной мРНК – так называемых РНК-переключателей (рис. 26, А). В ДНК эти участки находятся в транскрибируемой, но не транслируемой области оперона, перед началом кодирующей области, то есть в лидерной последовательности. Другими словами, они транскрибируются первыми. В этой области оперона находятся небольшие палиндромные последовательности, чередующиеся с непалиндромными. В результате, как только этот участок ДНК будет транскрибирован, еще далеко не законченная молекула РНК может приобрести третичную структуру со шпильками и одноцепочечными петлями. В третичной структуре переключателя, уникальной для каждого регулируемого таким способом оперона, есть рецепторный участок. Пространственная форма этого 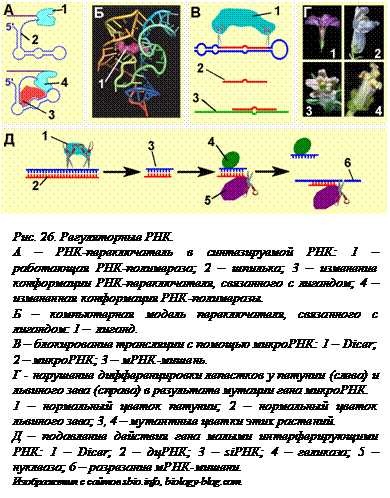 участка такова, что к ней, как ключ к замку, подходит молекула того вещества, которое является продуктом кодируемого этим опероном фермента. Связывание лиганда с рецептором и позволяет переключателю принять такую конформацию, что продолжение синтеза РНК становится невозможным. Таким образом, если продукта произведено достаточно для нужд клетки, лишняя молекула свяжется с рецептором, переключатель изменит конформацию, и транскрипция закончится, едва успев начаться. К настоящему времени открыты не только «РНК-выключатели», но и «РНК-включатели». Изменение конформации «РНК-включателей» позволяет транскрипции успешно идти до конца. Лигандом в таком случае может являть сё убстрат, для обработки которого и нужен фермент, кодируемый данным геном.
участка такова, что к ней, как ключ к замку, подходит молекула того вещества, которое является продуктом кодируемого этим опероном фермента. Связывание лиганда с рецептором и позволяет переключателю принять такую конформацию, что продолжение синтеза РНК становится невозможным. Таким образом, если продукта произведено достаточно для нужд клетки, лишняя молекула свяжется с рецептором, переключатель изменит конформацию, и транскрипция закончится, едва успев начаться. К настоящему времени открыты не только «РНК-выключатели», но и «РНК-включатели». Изменение конформации «РНК-включателей» позволяет транскрипции успешно идти до конца. Лигандом в таком случае может являть сё убстрат, для обработки которого и нужен фермент, кодируемый данным геном.
Как оказалось, такой способ регулирования транскрипции широко распространен в природе. Сейчас он найден у представителей всех надцарств живой природы – не только у бактерий, но и у архей и у эукариот. Исследователи считают, что это, возможно, самый древний из способов регуляции активности генов, возникший еще в «РНК-мире», на стадии «добелковой» жизни (см. § 23).
МикроРНК. То, что в клетке есть молекулы РНК малого размера, было известно давно. Однако на них долго не обращали внимания, считая их «обломками» более длинных молекул. Потом было обнаружено, что некоторые малые РНК (sРНК) участвуют в процессинге информационных и рибосомных РНК. Но в самом конце ХХ века неожиданно выяснилось, что такие РНК могут выключать работу некоторых генов. Сначала короткие регуляторные РНК (длиной всего в 22 нуклеотида) были обнаружены у круглого червя C. elegans. Они контролировали правильное чередование личиночных стадий развития этого организма. Довольно долго считали это редким случаем, исключением, присущим именно этим червям. Но в начале ХХI века обнаружилось, что такой способ остановки действия генов, без участия белков, а только с помощью коротких молекул РНК, распространен в живой природе очень широко. Такие РНК, получившие название микроРНК (mi РНК), обнаружены у растений, дрозофилы, человека. Недавно их обнаружили и у одноклеточных организмов. Сейчас известны сотни видов микроРНК. Правда, функции известны только для некоторых из них, но эти функции чрезвычайно важны. МикроРНК участвуют в регуляции формирования осей тела и дифференцировке тканей в эмбриогенезе животных (и человека), направляют развитие нервной системы у рыб, направляют дифференцировку некоторых органов у растений. Так, мутация по одному из генов микроРНК приводит к тому, что у растений-мутантов вместо лепестков образуются тычинки (рис. 26, Г). У дрозофилы блокирование действия некоторых микроРНК приводит к гибели развивающихся эмбрионов. Недавно было показано, что нарушение работы некоторых микроРНК может приводить к развитию рака и некоторых других заболеваний человека.

