




§ 24. Возникновение клеток.
Пробионты. Гипотетические живые системы, составлявшие РНК-мир, ученые назвали пробионтами – предшественниками современного живого мира. Были ли эти древнейшие живые системы клетками? В принципе возникновение липидных мембран и мембранных пузырьков представить себе несложно. Жирные кислоты могли возникать абиогенно, а образование липидных мембран – хорошо известный пример самоорганизации (см. § 22). Вполне возможно, что такие липидные пузырьки и возникали, но все они неминуемо должны были быть разными, в зависимости от того, что находилось в растворе в момент образования пузырьков. Чтобы действительно стать предшественниками клеток, пробионты должны были самовоспроизводиться, поэтому те из пузырьков, где не было самовоспроизводящейся РНК, просто «вымерли».
Какими же были «выжившие» пробионты? Очевидно, что в основе их функционирования были автокаталитические гиперциклы, включавшие РНК. Предполагают, что содержавшиеся в них молекулы РНК были небольшими, но их было много – порядка 300 разных молекул. По-видимому, четкого самовоспроизведения пробионтов еще не существовало, поэтому они все были разными. Безусловно, среди них была широко распространена рекомбинация, то есть обмен молекулами (или частями молекул) РНК. Механизмы защиты собственного генетического материала от чужеродной информации возникли позже. Вряд ли каждый отдельный пробионт мог в полной мере осуществлять все метаболические и информационные процессы, поэтому в каждом конкретном месте обитания (теплой луже, окрестности черного курильщика и т.п.) среди пробионтов должна была существовать широкая кооперация. Способные к кооперации и рекомбинации пробионты представляли собой своеобразную «первобытную коммуну», в которой разные члены осуществляли разные метаболические пути, а продукты поступали в более или менее общее пользование. Другими словами, жизнь должна была возникать сразу в виде экосистемы!
В таких условиях неизбежно должны были появиться «паразиты», которые использовали продукты деятельности «честных коммунаров», сами ничего не производя. Предполагается, что именно так могли возникнуть вирусы (разумеется, только содержащие РНК). О происхождении вирусов существует три группы гипотез, согласно которым вирусы представляют собой: 1) остатки доклеточных форм жизни; 2) потомки клеток, которые в результате перехода к паразитизму потеряли собственный метаболизм; 3) «одичавшие гены», например, вырезанные в результате сплайсинга интроны, которые не были расщеплены на нуклеотиды, а приобрели способность к размножению внутри клеток. Скорее всего, верны все три гипотезы, а вирусы являются сборной группой. Нередко говорят, что вирусы не могут быть доклеточными формами, так как для своей жизнедеятельности нуждаются в клетках. Это не совсем так – вирусы нуждаются не в клетках, а в метаболизме. Если вирусу предоставить все необходимое для репликации его нуклеиновой кислоты, то он прекрасно размножается и быстро эволюционирует, теряя ставшие ненужными в таких условиях гены. Так что для возникновения вирусов клетки были не нужны, достаточно было «коммунаров», производивших нужные молекулы. В настоящее время нередко вирусы рассматривают как реликты доклеточной жизни.
Переход к ДНК-миру. Однако, те системы, в которых паразиты возникли, для того, чтобы выжить, должны были защищаться. По-видимому, такими средствами защиты стали рестрикция (расщепление РНК вируса) и РНК-интерференция. Этот древний способ защиты от вирусов растения сохранили до настоящего времени. Так началась «гонка вооружений» между пробионтами (а потом и клетками) и вирусами, которая продолжается до сегодняшнего дня. Считается, что именно вирусы для защиты от рестриктаз «изобрели» ДНК (рис. 47, А). По-видимому, сначала была модифицирована рибоза. Отсутствие одной из гидроксильных групп сделало молекулу неузнаваемой для клеточных ферментов. Возникла так называемая У-ДНК, то есть ДНК, содержащая не тимин, а урацил. Несколько таких бактериофагов, содержащих У-ДНК, существует и в настоящее время. Затем модификация была продолжена – урацил был метилирован, получился тимин. Такие содержащие ДНК вирусы, если они были умеренными (а какая-то часть из них определенно таковыми была) могли, попадая в клетку и оставаясь там на некоторое время, передать клетке часть своей ДНК. Сначала получились клетки химерные, содержавшие в качестве генетического материала и РНК и ДНК, впоследствии вся РНК была заменена на ДНК, а РНК сохранила за собой роль «дирижера» в синтезе белков. Этот процесс обмена генетическим материалом между вирусами и клетками продолжается до сего времени. Полностью РНКовые клетки (или пробионты) не выдержали конкуренции с ДНКовыми именно из-за большей надежности ДНК. Получившиеся ДНКовые клетки послужили предками всех клеток, которые существуют в настоящее время. Они получили имя LUCA (Last Universal Common Ancestor – Последний Универсальный Общий Предок). LUCA дал начало всем клеткам, существующим сейчас на Земле – и бактериям и эукариотам.
Происхождение эукариот (рис. 47, Б). В настоящее время никто не сомневается в симбиотическом происхождении эукариотических клеток. Гипотеза о происхождении митохондрий и растительных пластид из внутриклеточных бактерий-эндосимбионтов была высказана еще в 1890 г. Альтманом. Эта гипотеза долгое время считалась слишком экзотической, и только молекулярно-генетические исследования дали доказательства того, что она соответствует действительности. В 1983 году эту же гипотезу выдвинула Л. Маргелис. Она предположила, что симбиотическое происхождение имеют не только пластиды и митохондрии, но и жгутики и центриоли.
Общепризнанная схема возникновения эукариотической клетки такова. С ростом концентрации кислорода в атмосфере жизнь анаэробных гетеротрофов усложнялась, так как кислород является сильным окислителем, способным разрушить все макромолекулы. Часть анаэробов для защиты от кислорода стала 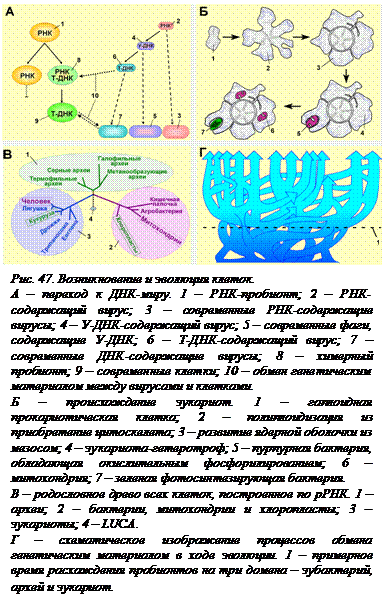 использовать специальные ферментативные системы для его контролируемой утилизации в различных реакциях окисления. Поскольку кислородное окисление дает почти в 20 раз больше энергии, чем бескислородное брожение, такие прокариоты вскоре перешли к использованию окислительного фосфорилирования для получения энергии. Такие аэробные гетеротрофы могли с бóльшим КПД, чем анаэробные бактерии, расщеплять органические вещества, производимые фото- и хемосинтетиками.
использовать специальные ферментативные системы для его контролируемой утилизации в различных реакциях окисления. Поскольку кислородное окисление дает почти в 20 раз больше энергии, чем бескислородное брожение, такие прокариоты вскоре перешли к использованию окислительного фосфорилирования для получения энергии. Такие аэробные гетеротрофы могли с бóльшим КПД, чем анаэробные бактерии, расщеплять органические вещества, производимые фото- и хемосинтетиками.
Существует представление, что разные царства эукариот имели разных предков, и эндосимбиоз бактерий возникал на разных этапах эволюции живых организмов. В пользу этого говорят отличия в строении митохондриальных геномов простейших, грибов, растений и высших животных.
Сравнение геномов показало, что предками митохондрий были прокариоты, принадлежавшие к группе альфапротеобактерий. К этой же группе принадлежат современные облигатные внутриклеточные паразитические бактерии вольбахия и риккетсии. Изучение взаимоотношений вольбахии и риккетсий с эукариотическими клетками позволило сделать предположение, что и предок митохондрий скорее также был не жертвой хищника, а «прирученным» паразитом.
Симбиоз предков растительных клеток с фотосинтетиками возникал в эволюции как минимум, трижды. На это указывает в частности наличие разных хлорофиллов у разных групп фотосинтетических эукариот. Считается твердо установленным, что первые фотосинтетики – цианобактерии, были предками красных водорослей. И те, и другие содержат хлорофилл а и пигменты фикобилины. У других организмов фикобилинов нет. Зеленые водоросли и произошедшие от них высшие растения содержат хлорофиллы а и b. Относительно недавно были обнаружены прокариоты, содержащие эти же хлорофиллы. Примечательной особенностью такой бактерии, называющейся Prochloron, является наличие развитой системы внутриклеточных фотосинтетических мембран. Независимое происхождение имеют хлоропласты золотистых водорослей. Он возникли в результате поглощения и «приручения» гетеротрофом не фотосинтезирующей бактерии, а эукариоты-фотосинтетика, уже имевшей свои хлоропласты!
Возникновение ядра. Но прежде, чем все это произошло, предшественники эукариот должны были приобрести способность к фагоцитозу, ведь у прокариот его нет. Для этого нужны, во-первых, белки цитоскелета, а во-вторых, в несколько раз больший объем клетки. По-видимому, увеличение объема было достигнуто в результате увеличения количества копий генома – полиплоидизации (в большой клетке продукты транскрипции единственной молекулы ДНК не смогут достаточно быстро достичь отдаленных частей клетки из-за малой скорости диффузии). Такие огромные полиплоидные бактерии существуют и в настоящее время.
Ядро возникло в результате того, что мезосомы, (впячивания плазмалеммы, к которым прикреплялись кольцевые ДНК), углублялись, расширялись, и, в конце концов, образовали разветвленную эндоплазматическую сеть, частью которой и является ядерная мембрана.
В дальнейшем основная часть генов из промитохондрий и прохлоропластов была перенесена в ядро, скорее всего, с помощью мобильных генетических элементов. При этом некоторая часть генов хлоропластов была перенесена не в ядро, а в митохондрию! В результате включении части генома одного из симбионтов в геном другого интеграция симбионтов становится необратимой. Новый геном может обеспечивать создание метаболических путей, приводящих к образованию полезных продуктов, которые не могут быть синтезированы ни одним из партнеров по отдельности. Так например, синтез стероидных гормонов позвоночных представляет собой сложную цепь реакций, часть из которых происходит в митохондриях, а часть – в эндоплазматической сети. «Захватив» гены предшественников органоидов, ядро получило возможность надежно контролировать функции симбионтов.
Три домена живой природы. Какая же прокариота была предшественницей цитоплазмы и ядра? Дело в том, что прокариоты – это не только бактерии. В 1977 Карл Вёзе решил с помощью методов молекулярной биологии изучить родственные отношения всех живущих организмов с целью прояснения их происхождения. Для этого была выбрана не ДНК, а РНК малой субъединицы рибосом – 16 S РНК для прокариот и 18 SРНК для эукариот. Этот выбор объясняется тем, что рибосомы есть у всех организмов во всех клетках. Кроме того, большинство мутаций в этой РНК смертельно для клетки, поэтому носители таких мутации не сохраняются. Безвредных мутаций в рибосомных РНК очень мало, поэтому они накапливаются очень медленно, и за более чем 3,5 млрд. лет РНК не изменились до неузнаваемости, что дает возможность их сравнивать. Были исследованы представители разных таксонов. Кроме ожидаемого результата – попадания РНК хлоропластов и митохондрий в ветвь прокариот, был получен результат и вполне неожиданный. Прокариоты оказались разделенными на две неродственные группы (им был позже присвоен ранг доменов) – собственно бактерий (эубактерий) и архей (рис. 47, В). Археи были известны и раньше под именем археобактерий, но считались небольшой малозначительной частью царства бактерий. Оказалось же, что они столь же далеки от бактерий, как и от эукариот. Поэтому сразу же встал вопрос, кем же был LUCA – бактерией или археей? Изучение самых разных сторон метаболизма, устройства и функционирования систем транскрипции и трансляции показало, что эукариотические клетки являются химерами – «смесью» бактерий и архей. Считается вполне вероятным, что эукариотическая клетка является результатом объединения (слияния) бактерии и археи. Возможно, именно это и положило начало полиплоидизации предшественника эукариот.
Археи микроскопически отличить их бактерий, практически невозможно. Средний размер и морфология клеток архей такие же, как и у большинства известных бактерий. Геном архей представлен двуцепочечной кольцевой ДНК и кольцевыми плазмидами. Большинство архей являются экстремофилами – они живут в содовых озерах, в экстремально соленых водоемах, в горячих вулканических котлах, в кипящих серных источниках, окрестностях «черных курильщиков» в океанских глубоководных впадинах при огромном давлении, в горячей серной кислоте, в толще земной коры до глубины по меньшей мере 6000 метров. Исходя из результатов глубоких бурений, ученые подсчитали, что биомасса всех архей на Земле может составлять порядка 1014 тонн, что в 40 раз больше, чем биомасса всех остальных форм жизни (включая бактерий и растения).
Среди архей есть гетеротрофы, питающиеся, например, метаном, и хемосинтетики. Некоторые экстремальные галофилы, живущие в водоемах с крайне высокой минерализацией обычно имеют в своих клетках красные (каротиноиды) и пурпурные (родопсины) пигменты, что позволяет им использовать фотосинтез для получения АТФ (хотя только за счет этого они жить не могут).
В морских осадках, в болотах, почвах рисовых полей живут метаногенные археи – это единственные представители живой природы, синтезирующие метан. Многие из них являются симбионтами некоторых простейших и многих животных (и человека). В частности, именно они снабжают своих хозяев витамином В12.
Одно из основных отличий архей от бактерий – различия в строении плазмалеммы. В состав фосфолипидов архейных мембран вместо D-глицерола входит L-глицерол, вместо жирных кислот – разветвленный высокомолекулярный (С20) спирт фитанил. Ветвящиеся молекулы фитанила могут создавать двуслойную мембрану, но могут и объединяться между слоями с образованием бифитанила. Результатом является образование однослойных мембран (но с двумя гидрофильными слоями), к чему не способны никакие другие живые организмы.
Структуры, связанные с механизмами хранения и реализации генетической информации (репликация, транскрипция) архей подобны эукариотическим: наличие гистоноподобных белков и нуклеосом, наличие интронов, ДНК-полимераза, РНК-полимераза, структура промотора, транскрипционные факторы. Хотя рибосомы архей по размеру ближе к прокариотическим, их белковый состав ближе к рибосомам эукариот. Это дает серьезные основания для того, чтобы считать, что предшественником ядра и всех информационных процессов эукариот были археи.
Горизонтальный перенос информации. Огромную роль в переносе генетического материала между разными клетками играли вирусы и мобильные генетические элементы. У прокариот и в настоящее время процессы «горизонтального переноса» (то есть переноса, не связанного с размножением, фактически в пределах одного поколения) очень масштабны, а на ранних этапах эволюции они были особенно важны, так как значительно ускоряли эволюцию. Анализ генов, являющихся общими или уникальными для всех трех доменов живого, показал, что, по-видимому, на самом раннем этапе развития жизни единого предка, скорее всего, не было. Поэтому «родословное древо всего живого» является не столько ветвящимся древом, сколько сетью (рис. 47, Г). Единство живого оказывается обусловленным не происхождением от одного уникального добиологического предка, а теснейшим переплетением линий, идущих от разных предковых систем, «великим объединением» достижений на пути от неживого к живому.
§ 25. Генные сети
Функциональные сети – способ существования и работы сложных систем. Живые системы являются, пожалуй, самыми сложными из известных систем. Они обладают всеми теми свойствами, о которых уже говорилось: эмерджентностью (см. Введение), иерархичностью устройства, способностью к эволюции; наследственностью; способностью к саморегуляции – поддержанию гомеостаза в изменяющихся условиях внешней среды. На всех иерархических уровнях живого действует огромное количество прямых и обратных регулирующих связей между элементами. Как правило, ученые-биологи в каждом конкретном исследовании изучают какой-нибудь один процесс или цепь процессов: например, пищевую цепь в биоценозе, или последовательность реакций окислительного фосфорилирования в митохондрии, или влияние антибиотика на репликацию вируса и т. п. В действительности же каждый член биоценоза входит не в одну, а во многие пищевые цепи, на протекание биохимических реакций оказывает влияние огромное количество других процессов, антибиотик влияет не только на вирус, но и многие другие процессы в клетке, не связанные с репликацией инфекционного агента.
Таким образом, все эти цепи – последовательности событий – могут разветвляться и сливаться, образуя сеть. Такие функциональные сети – способ существования и работы всех систем, в том числе и живых. Именно сетевой принцип обеспечивает надежность и устойчивость систем к разнообразным внешним воздействиям и их способность к саморегуляции.
Усложнение биологических систем. Мы уже неоднократно говорили о том, что сложные системы способны к самоорганизации. Самоорганизация – это образование все более и более сложных систем. За счет чего идет это усложнение – за счет увеличения количества элементов в системе? Или за счет образования все новых и новых связей между ними? Еще несколько лет назад считали, что количество генов у человека должно быть не менее 100000. Потом это цифру снизили до 50000, а сейчас считается, что у человека только около 23000 генов. В то же время расшифрованный в 2011 году геном планктонного рачка дафнии содержит около 31000 генов. Выходит, человек устроен проще, чем беспозвоночные? Конечно, это не так. Усложнение организма человека по сравнению с червем было достигнуто в ходе эволюции не за счет увеличения числа элементов – генов, а за счет более эффективного использования возможностей взаимодействия и взаимного регулирования имеющихся в системе элементов, за счет создания более сложных и более разветвленных функциональных сетей.
На каждом уровне организации живых систем существуют свои функциональные сети. На уровне экосистем это пищевые сети, на уровне организма – нейрогуморальные взаимодействия. На уровне клетки работают генные сети.
Информационная биология. Изучение свойств генных сетей является одной из задач новой научной дисциплины – информационной биологии. За последние десятилетия биологические науки стремительно развиваются, накапливая гигантские объемы информации. Только геном человека, почти полностью к настоящему времени прочитанный, содержит более 3 миллиардов пар нуклеотидов. А ведь сейчас прочитаны геномы мыши, дрозофилы, риса, некоторых других животных и растений, тысяч бактерий и вирусов. Расшифрованы сотни тысяч аминокислотных последовательностей разнообразных белков, для десятков тысяч белков определена пространственная конформация. Осмыслить и понять такое количество информации можно только, используя современные информационные технологии. Для таких сложных взаимодействий многочисленных элементов биологических систем в принципе невозможны такие простые законы как, например, закон всемирного тяготения. Тем не менее, и они подчиняются своим закономерностям. Моделирование функциональных сетей помогает найти эти закономерности, помогает понять, как та или иная сеть будет реагировать на определенные воздействия. Этим и занимается информационная биология. Один из разделов этой науки – изучение структуры и свойств генных сетей в живых организмах.
Генные сети – это группы координировано работающих генов, контролирующих все основные процессы, протекающие в живых системах – рост, индивидуальное развитие, функционирование систем органов, ответ на внешние воздействия и др. Когда говорят о генной сети, учитывают все составляющие части того или иного процесса. Например, в клетках человека существует несколько генов, отвечающих за реакцию на проникновения вируса. Молекулярный биолог будет изучать структуру каждого из этих генов, особенности их транскрипции, сплайсинга, регуляции, свойства белков, кодируемых этими генами. Вирусолог изучит динамику развития инфекционного процесса внутри клетки, особенности процессов проникновения вируса, сборки вирионов, выхода частиц из клетки. Врача будут интересовать интенсивность действия собственных защитных механизмов клетки, реакция на лекарства. В клетке же все эти разнонаправленные процессы происходят одновременно, взаимно влияя друг на друга и взаимно друг друга регулируя. Для того, чтобы расшифровать генную сеть того или иного процесса, нужны все данные, полученные самыми разными специалистами. Нужно проанализировать все реакции, все вещества, участвующие в этом процессе, все гены, кодирующие все белки, участвующие в нем, и, главное, все регуляторные пути, связывающие все в одну сеть.
Генная сеть состоит из нескольких частей: 1) ансамбля координировано работающих генов; 2) кодируемых этими генами белков; 3) путей передачи к генам сигналов, включающих или выключающих их транскрипцию в ответ на внешние воздействия; 4) путей передачи сигналов обратной связи; 5) веществ, осуществляющих эти переключения генной сети – гормонов, низкомолекулярных веществ, АТФ и др. Очень часто координация работы генов, входящих в сеть, обеспечивается тем, что эти гены регулируются одним и тем же белком-регулятором, кодируемым «центральным» геном. Этим достигается одновременное включение или выключение целой группы генов. Соответственно сложности системы и схема генной сети может иметь несколько уровней сложности. Например, на рисунке 48, А приведена простая схема генной сети осуществления ответа клетки на попадание вируса. Она включает всего три гена и три белка – продукта этих генов. При попадании вируса в клетку происходит активация гена интерферона-β (IFN-β). Белок (фактор транскрипции) IRF-1 вначале усиливает работу гена, продуцирующего интерферон, а позднее запускает синтез другого белка IRF-2, который подавляет производство интерферона, обеспечивая возвращение системы в исходное состояние. Однако следует иметь в виду, что работа каждого из этих генов контролируется разными процессами, что синтез этих белков зависит от разных условий – наличия ферментов, аминокислот и т. д. Регуляция действия генных сетей происходит на разных уровнях – транскрипции, трансляции, сплайсинга, деградации белков, активного мембранного транспорта и т.д. Если мы примем во внимание хотя бы часть этих процессов и внесем их в схему, она примет значительно более сложный вид. Если же учесть, что в каждой клетке одновременно происходит большое количество самых разнообразных процессов, каждый из них имеет свою генную сеть, но при этом сети могут пересекаться, то станет понятной невероятная сложность осуществления всех процессов жизнедеятельности клетки и организма. Генная сеть целого организма, даже такого простого как нематода С. elegance, все тело которого состоит всего из 945 клеток, уже выглядит очень сложной и запутанной. Геном этой нематоды полностью расшифрован, все его гены и их взаимоотношения известны (рис. 48, Б). Генные сети более сложных организмов неизмеримо сложнее. Например, генная сеть, контролирующая процессы дифференцировки и созревания эритроцитов человека, состоит из примерно 500 генов и огромного количества других компонентов (информационных и транспортных РНК, белков, АТФ и других метаболитов). И это – только одна из генных сетей человека. Одновременно с ней существуют и работают тысячи других сетей, не менее важных для организма человека. Они объединяются в общую глобальную генную сеть организма.
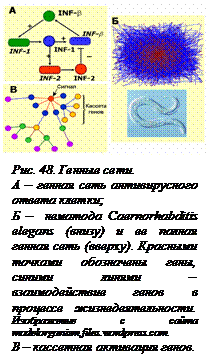 Наиболее сложными являются так называемые интегративные генные сети. Они включают в себя структурные и регуляторные гены, отвечающие за регуляцию таких сложных процессов, как например, индивидуальное развитие организма. В таких случаях один регуляторный ген может одновременно, с помощью одного и того же сигнала включать сразу много разных генов. Это так называемая кассетная активация генов (рис. 48, В). Сигналом активации обычно служит транскрипционный фактор – белок, без которого не может начаться транскрипция. Гены узнают только «свой» транскрипционный фактор. Это позволяет регулировать работу генов в многоклеточном организме – в разных тканях и клетках синтезируются разные факторы транскрипции и поэтому работают разные гены. Активированный ген тоже может иметь свою кассету генов. В результате один сигнал может запустить целый каскад самых различных реакций в клетке. Изучение всех этих взаимодействий может показаться невозможным. Однако делу помогает одно из основных свойств сложных систем – иерархичность устройства. Поведение высших уровней организации системы очень мало зависит от того, как конкретно устроены и работают низшие «этажи» иерархии, важны только сигналы, информация, посылаемая ими «наверх». Тем не менее, чтобы понять, что именно важно, а что не очень важно для работы той или иной системы, нужно изучить и проанализировать работу подсистем каждой сложной системы клетки или организма. Ясно, что это возможно только с применением мощной вычислительной техники. Почему же нельзя ограничиться изучением отдельных компонентов и процессов в клетке? Это следует из другого системного свойства – эмерджентности. Ведь именно в системе могут возникнуть новые свойства, которые мы не сможем обнаружить, изучая только элементы этой системы, без принятия во внимание связей между ними.
Наиболее сложными являются так называемые интегративные генные сети. Они включают в себя структурные и регуляторные гены, отвечающие за регуляцию таких сложных процессов, как например, индивидуальное развитие организма. В таких случаях один регуляторный ген может одновременно, с помощью одного и того же сигнала включать сразу много разных генов. Это так называемая кассетная активация генов (рис. 48, В). Сигналом активации обычно служит транскрипционный фактор – белок, без которого не может начаться транскрипция. Гены узнают только «свой» транскрипционный фактор. Это позволяет регулировать работу генов в многоклеточном организме – в разных тканях и клетках синтезируются разные факторы транскрипции и поэтому работают разные гены. Активированный ген тоже может иметь свою кассету генов. В результате один сигнал может запустить целый каскад самых различных реакций в клетке. Изучение всех этих взаимодействий может показаться невозможным. Однако делу помогает одно из основных свойств сложных систем – иерархичность устройства. Поведение высших уровней организации системы очень мало зависит от того, как конкретно устроены и работают низшие «этажи» иерархии, важны только сигналы, информация, посылаемая ими «наверх». Тем не менее, чтобы понять, что именно важно, а что не очень важно для работы той или иной системы, нужно изучить и проанализировать работу подсистем каждой сложной системы клетки или организма. Ясно, что это возможно только с применением мощной вычислительной техники. Почему же нельзя ограничиться изучением отдельных компонентов и процессов в клетке? Это следует из другого системного свойства – эмерджентности. Ведь именно в системе могут возникнуть новые свойства, которые мы не сможем обнаружить, изучая только элементы этой системы, без принятия во внимание связей между ними.
§ 26. Детерминация и дифференцировка.
Разнообразие клеток. Каждый многоклеточный организм представляет собой сложную систему, составленную из разных типов клеток. Клетки многоклеточного организма, объединенные в разные органы и ткани, отличаются друг от друга строением и выполняемой функцией. Специализация клеток позволила многоклеточным организмам значительно расширить возможности приспособления к самым разным условиям существования. Возникновение разных типов клеток – пример самоорганизации в ходе эволюции. Как правило, ДНК в ядрах всех клеток организма одинакова, гены «домашнего хозяйства» работают во все клетках, но в каждом типе клеток работает только небольшая часть «генов роскоши» (см. § 17). Продукты этих генов нужны не только производящим их клеткам, но и всему организму в целом – это, например, гормоны, пищеварительные ферменты, антитела и многие другие вещества. Таких веществ в организме очень много, и производятся они специализированными, дифференцированными, клетками. В дифференцированных клетках работают те гены, которые «молчат» во всех остальных клетках. Набор работающих генов специфичен для каждого типа клеток.
Свойства любой дифференцированной клетки закодированы в ДНК зиготы. Каким же образом клетки «выбирают», какую часть генетической информации им реализовать в процессе дифференцировки, а какую оставить неиспользованной, когда «включить» или «выключить» те или иные гены?
Дифференцировка и детерминация. Дифференцировка – это приобретение клетками специфических признаков и свойств в ходе индивидуального развития организма. Клетки становятся дифференцированными в результате того, что включается работа определенных генов, тогда как другие выключаются. Исходный момент в процессе дифференцировки – детерминация (обусловленность), выбор программы для дальнейшего развития клетки. Явление детерминации охватывает небольшой интервал времени, когда исходные клетки еще никак внешне не отличаются друг от друга. В ходе онтогенеза многоклеточных животных таких моментов детерминации бывает несколько, и захватывают они клетки, являющиеся предшественниками всего организма или только отдельных тканей. Как правило, еще в неоплодотворенной яйцеклетке уже предопределено, где будут спинная и брюшная сторона, передний и задний концы зародыша. Это определяется градиентами распределения некоторых белков и долгоживущих мРНК, запасенных в цитоплазме в процессе формирования яйцеклетки. Долгоживущие матрицы информационной РНК для такого белка синтезированы еще в неоплодотворенной яйцеклетке. Молекулы закреплены на одном из полюсов яйца, поэтому белок, который начинает синтезироваться после оплодотворения, распространяется из одного места. Поскольку белок живет недолго, а синтезируется в течение довольно длительного времени, градиент его концентрации сохраняется все время. Другими словами, определение осей тела зародыша осуществляется материнскими генами еще до оплодотворения.
Клетки многоклеточного зародыша синтезируют сигнальные вещества – морфогены, запускающие работу определенных генов тех клеток, которых они достигают в процессе диффузии и которые имеют рецепторы для этого морфогена. Морфогены являются сигналами, под действием которых происходит детерминация, а затем и дифференцировка клеток. Дифференцирующиеся клетки также могут синтезировать свои морфогены, которые служат сигналами детерминации и дифференцировки следующих клеток (рис. 49, А). Так последовательно включаются гены клеток, занимающих разное положение относительно переднего и заднего концов зародыша. Первыми активируются гены, определяющие сегментацию зародыша (рис. 49, Б, В). Дело в том, что тело большинства многоклеточных животных разделено на сегменты. У некоторых из  них (членистоногие, кольчатые черви) эта сегментация видна очень хорошо, у других в результате интенсивной миграции клеток в ходе формирования систем органов следы этой сегментации становятся мало заметными. Но даже у млекопитающих и птиц следы этой сегментированности остаются на всю жизнь – это в основном позвонки и ребра.
них (членистоногие, кольчатые черви) эта сегментация видна очень хорошо, у других в результате интенсивной миграции клеток в ходе формирования систем органов следы этой сегментации становятся мало заметными. Но даже у млекопитающих и птиц следы этой сегментированности остаются на всю жизнь – это в основном позвонки и ребра.
Гомеозисные гены. Продукты генов сегментации включают гены, которые отвечают за особенности развития каждого конкретного сегмента. Эти гены, называемые гомеозисными, определяют общий план тела – где и какие возникнут конечности, где будет голова, грудь, глаза и т. д. Продукты гомеозисных генов – транскрипционные факторы (см. § 11), белки, обладающие сродством к регуляторным областям ДНК. Они подавляют или активируют действие генов, запуская каскад событий, приводящий к развитию уже конкретной структуры. Мутации гомеозисных генов могут привести к включению или выключению генов, которые не должны работать в определенном участке организма. Это приведет к возникновению правильных структур тела, но на неправильном месте. Например, мутация в гене bithorax у дрозофилы приводит к тому, что третий грудной сегмент у мухи развивается так, как должен развиваться второй. Поэтому у взрослой мухи оказывается лишняя пара крыльев (рис. 49, Г). Мутация в гене antennapedia приводит к тому, что в головном сегменте не происходит дифференцировки антенн, а вместо этого развивается лишняя пара ног (рис. 49, Д). Интересно, что очень похожие гены определяют план тела как у насекомых, так и у позвоночных.

