




В некоторых случаях мобильные генетические элементы прокариот могут передаваться ими эукариотическим клеткам. Например, почвенная бактерия Agrobacterium tumefaciens вызывает развитие так называемых корончатых галлов у растений, передавая им свою плазмиду, которая встраивается в ДНК зараженной растительной клетки. Подробнее об этом будет сказано в § 34.
Значительно более многочисленны и разнообразны ретроэлементы (от лат. retro - прежде, обратно) эукариот. Всех их объединяет общая черта – использование особого матричного синтеза – обратной транскрипции (см. § 6). Активные (не «испорченные» мутациями) ретроэлементы обычно содержат гены ферментов обратной транскриптазы (иногда ее называют ревертазой) и интегразы, необходимой для встраивания в геном синтезированной копии. Образующаяся в результате транскрипции РНК служит матрицей для синтеза ревертазы и интегразы. Затем ревертаза осуществляет обратную транскрипцию этой РНК, а интеграза встраивает полученную ДНК в геном клетки. В результате в геноме оказывается новая копия элемента, при этом старая остается на месте. Поэтому в отличие от ДНК-транспозонов ретроэлементы активно размножаются в геноме. Из-за этого значительную долю геномов большинства эукариот составляют копии ретроэлементов. К счастью (ведь геном не может увеличиваться бесконечно), значительная часть их неактивна из-за накопившихся мутаций.
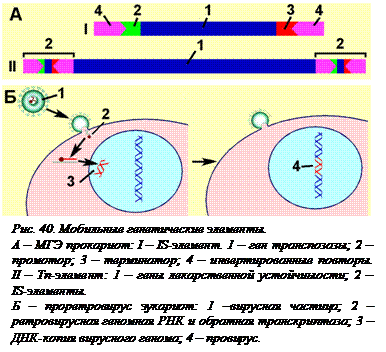 Другую группу ретроэлементов составляют ретровирусы. Ретровирусоми называют такие вирусы, для которых обратная транскрипция является обязательным этапом в жизненном цикле. Они отличаются от ретроэлементов тем, что содержат также гены, кодирующие белки оболочки вируса. Их геномные РНК, попав в клетку, так же как и ретроэлементы, служат матрицами для синтеза ДНК. На обоих концах этой двухцепочечной ДНК находятся LTR – длинные концевые повторы нуклеотидов (long terminal repeat), которые нужны для встраивания этой ДНК в геном клетки. Образующаяся в результате обратной транскрипции ДНК встраивается в геном клетки хозяина с помощью фермента интегразы. Этих ферментов в клетке нет, поэтому вирусы приносят их с собой. Встроенную ДНК называют провирусом (рис. 40, Б).
Другую группу ретроэлементов составляют ретровирусы. Ретровирусоми называют такие вирусы, для которых обратная транскрипция является обязательным этапом в жизненном цикле. Они отличаются от ретроэлементов тем, что содержат также гены, кодирующие белки оболочки вируса. Их геномные РНК, попав в клетку, так же как и ретроэлементы, служат матрицами для синтеза ДНК. На обоих концах этой двухцепочечной ДНК находятся LTR – длинные концевые повторы нуклеотидов (long terminal repeat), которые нужны для встраивания этой ДНК в геном клетки. Образующаяся в результате обратной транскрипции ДНК встраивается в геном клетки хозяина с помощью фермента интегразы. Этих ферментов в клетке нет, поэтому вирусы приносят их с собой. Встроенную ДНК называют провирусом (рис. 40, Б).
К ретровирусам относится вирус иммунодефицита человека, вызывающий СПИД и так называемые онкорнавирусы. Онкорнавирусы – это онкогенные РНК-содержащие вирусы. Как и другие ретровирусы, они могут долгое время существовать в зараженной клетке в виде провируса. При действии на клетку неблагоприятных условий (стресс, радиация, химические вещества) провирус транскрибируется, образуются вирусные белки, в том числе и те, которые приводят к злокачественной транформации клетки (см. §§ 37 и 38).
Совершенно очевидно близкое родство ретротранспозонов и ретровирусов. Однако еще не вполне ясно, кто от кого произошел. Ученые склоняются к мысли, что ретровирусы произошли от ретротранспозонов в результате приобретения генов белков оболочки. Однако, по-видимому, возможен и обратный процесс – деградация вируса. На это указывается присутствие в геноме человека и других животных множества так называемых эндогенных ретровирусов — встроенных вирусных геномов. Если ретровирусу удается проникнуть в половые клетки, он сможет передаваться по наследству. Такие ретровирусы, встроившиеся когда-то в геном половых клеток и устойчиво передающиеся от родителей к потомкам, и называют эндогенными ретровирусами. Если в череде поколений возникает мутация, портящая один из генов вируса, то он теряет способность вызывать инфекционный процесс. Отбором такие мутации не отсеиваются, и накопление мутаций приводит к постепенной деградации вируса. Если в результате мутации будет испорчен ген, кодирующий белок вирусной оболочки, а гены ревертазы и интегразы останутся целыми, такой провирус превратится в ретротранспозон. Его дальнейшая деградация может привести к тому, что останутся только одни LTR – длинные концевые повторы. Эти повторы могут содержать промотор, сигнал полиаденилирования и различные регуляторные последовательности, влияющие на уровень транскрипции. Эти «беспризорные» регуляторные участки могут быть использованы клетками для создания новых генов.
Псевдогены. Этим термином обозначают нефункциональные «испорченные» гены. Чтобы такие гены в геноме вообще существовали, необходимо предварительное удвоение гена с тем, чтобы нормальная, неиспорченная, версия гена в геноме была и выполняла свою функцию. Известно два типа псевдогенов – процессированные и непроцессированные. Процессированные образуются с помощью ретровирусов или ретротранспозонов. Это происходит в том случае, когда транскрибированная с обычного гена матричная РНК, подвергшаяся сплайсингу, случайно оказывается матрицей для обратной транскриптазы и интегразы. Получившаяся ДНК-копия матричной РНК будет при этом встроена в геном. Но эта копия не будет содержать интронов, характерных для клеточного гена и, самое главное, не будет содержать промотора, который не транскрибируется и поэтому отсутствует в мРНК. Так получится псевдоген – он содержит все кодирующие белок области, но не может считываться, так как не имеет промотора. Непроцессированные пседогены – результат удвоения сегмента ДНК, содержащего этот ген. Если в одной копии гена произойдет мутация, делающая невозможным нормальное функционирование гена, то это и будет псевдоген. Организм при этом не пострадает, так как нормально работающая копия гена у него останется. В геноме человека содержится не менее 30000 псевдогенов, больше чем нормальных генов. Среди них больше всего (11% генома) так называемых Alu-повторов. Alu-повторы являются процессированными псевдогенами 7SL-РНК – одной из клеточных малых регуляторных РНК. Подавляющее большинство этих повторов неактивно, но некоторые продолжают транскрибироваться (в них есть внутренний промотор). Особенно активно они транскрибируются в клетках головного мозга. Предполагается, что они могут как-то участвовать в функционировании мозга.
Псевдогены сохраняются в геноме просто потому, что не существует активного механизма их удаления. Поскольку они ни для чего не нужны, они не подвержены действию отбора. Поэтому в них накапливаются мутации (это позволяет установить время появления этого псевдогена). Однако иногда случается, что псевдоген может дать начало новому гену. Мы знаем, что в геномах много так называемых LTR – повторов, оставшихся от исчезнувших ретротранспозонов и содержащих промотор. Если при перемещении по геному такой сохранивший активность промотор пристроится к процессированному псевдогену, тот может начать работать. Если за время его существования в нем произошли мутации, новый ген будет отличаться от первоначального гена не только отсутствием интронов, но и тем, что будет кодировать отличающийся белок. Другими словами, возникнет новый ген. Если псевдоген непроцессированный, то произошедшая в нем новая мутация может восстановить активность этого гена также с изменением функции, то есть с возникновением нового гена. Возникшие таким образом новые гены будут отличаться от старых не намного. Предполагают, что так возникли так называемые семейства генов. Многие гены человека повторены в геноме от нескольких единиц до нескольких сотен раз и образуют мультигенные семейства. Эти гены обычно сгруппированы в определенных районах одной либо нескольких хромосом. Во многих мультигенных семействах наряду с функционально активными генами содержатся псевдогены. Примерами мультигенных семейств могут служить гены рибосомных, транспортных и ядерных малых РНК, гены глобинов, тубулинов, миоглобина, актина, гены обонятельных рецепторов и многих других.
Особую группу составляют ретроэлементы прокариот (а также митохондрий и пластид эукариот) – так называемые ретроинтроны. Эти элементы содержат только один ген белка, обладающего активностью обратной транскриптазы и эндонуклеазы. Кроме того, РНК этого элемента обладает рибозимной активностью, благодаря которой осуществляется самосплайсинг – вырезание интрона из пре-мРНК, считанной с этого гена. В результате трансляции мРНК синтезируется фермент. Его эндонуклеазная активность позволяет создать разрыв в клеточной ДНК. Вырезанный интрон встраивается в этот разрыв, а ревертазный активный центр белка синтезирует ДНКовую копию интрона.
Мобильные генетические элементы и эволюция (см. также § 30). Перемещение мобильных элементов не остается без последствий для клеток. Таким последствием может быть, например, включение или выключение гена, может нарушиться регуляция гена.
У прокариот транспозоны вместе с плазмидами и фагами (в которые они легко интегрируются) способны осуществлять обмен находящихся в них генов между весьма отдаленными видами бактерий, поэтому они играют чрезвычайно важную роль в эволюции бактерий, включая адаптацию их к лекарствам и продуцирования ими новых токсинов.
Как правило, у эукариот обычно транспозиции МГЭ происходят чрезвычайно редко. Однако они могут в определенных условиях (ухудшение условий обитания, слишком близко родственное скрещивание) учащаться. Известны так называемые транспозиционные взрывы, ведущие к одновременным перемещениям (транспозициям) целого ряда разных мобильных элементов.
Таким образом, перемещение и размножение мобильных генетических элементов – важный процесс, создающий генетическое разнообразие, которое служит материалом для эволюции. Однако, слишком интенсивное перемещение мобильных элементов может нарушить нормальную жизнедеятельность. Но оказалось, что клетки эукариот могут целенаправленно регулировать активность своих МГЭ. Например, у дрозофилы во время образования половых клеток начинают работать гены особых малых (24-29 нуклеотидов) регуляторных РНК, так называемых пиРНК, и образующих с ними комплекс специальных белков Piwi. Действуют они аналогично интерферирующим РНК (§ 13). Последовательности нуклеотидов пиРНК, комплементарные участкам разных МГЭ, узнают РНК, считанные с мобильных элементов, а находящиеся в комплексе с ними Piwi -белки уничтожают молекулы РНК. Белки этого семейства есть и у млекопитающих (они называются MIWI и MILI); они тоже синтезируются во время гаметогенеза и образуют комплексы с пиРНК.
«Молекулярное одомашнивание». Некоторые ферменты или другие белки, свойственные вирусам или транспозонам, обычным клеткам не свойственны. Однако иногда клеткам удается «приручить» какой-нибудь транспозон, сделав его собственным геном с определенной функцией. Это явление и назвали «молекулярным одомашниванием». Ученым удалось установить, что такие события действительно имели место в ходе эволюции. Несколько «прирученных» генов оболочечных белков эндогенных ретровирусов человека, работают, например, в плаценте. «Прирученные» гены транспозаз дали начало генам так называемых RAG-белков, которые создают разнообразие генов антител путем вырезания и перекомбинирования фрагментов геномной ДНК (см. § 27). Растения, приручив гены транспозаз, используют их для другой цели – слежения за длиной светового дня. От ретротранспозона происходит и клеточный фермент теломераза.
Эволюция геномов. Геномная дупликация, возникающая при нерасхождении хромосом в ходе деления клетки – один из важнейших путей увеличения размера генома эукариот в ходе эволюции. В эволюционной истории позвоночных было, по крайней мере, два раунда полногеномной дупликации. Кроме того, значительная часть геномов высших животных и растений является результатом сегментных дупликаций (то есть удвоения отдельных частей генома хромосом или их участков). Удвоение генов создает возможность постепенного накопления мутаций и создания новых генов, не нарушая при этом работы старых. Новые гены могут также появиться в результате «молекулярного одомашнивания», то есть «экспроприации» и приспособления для собственных целей чужеродных генов.
Другим важным путем создания новых генов является возникновение новых комбинаций уже существовавших экзонов. Это может быть результатом перемещения транспозонов, «прихвативших» с собой часть генетического материала. В результате появятся разные гены, содержащие одинаковые экзоны, и, соответственно, белки, содержащие одинаковые домены, но выполняющие разные функции. В ходе эволюции эукариот такое происходило неоднократно.
Важным может быть не только приобретение новых экзонов, но и новых интронов, которые создают новые возможности для альтернативного сплайсинга. Кроме того, они могут содержать различные регуляторные элементы. Ученые считают, что в эволюции млекопитающих (и человека) основную роль играло именно усложнение регуляции, а не возникновение новых генов.
§ 21. Геном митохондрий и хлоропластов.
У всех эукариот генетическая информация содержится не только в хромосомах клеточного ядра, но и в митохондриях и пластидах — самовоспроизводящихся полуавтономных органеллах клетки, имеющих собственный геном.
Геном митохондрий. В то время как ядерный геном представляет собой совокупность линейных молекул ДНК гаплоидного набора хромосом, митохондриальные геномы больше похожи на геномы прокариот. У высших животных они представляют собой одну или несколько кольцевых молекул ДНК (мтДНК), прикрепленных к внутренней мембране митохондрий. У растений такие кольцевые ДНК (их всегда несколько) могут быть разных размеров. Большие кольца образованы объединенными малыми. Митохондрии некоторых видов растений наряду с кольцевыми содержат и линейные молекулы ДНК. Крупные клетки простейших обычно содержат одну большую митохондрию с огромным количеством колец ДНК самого разного размера (огромных макси-колец и маленьких мини-колец), которые к тому же продеты друг в друга (рис. 41). Такие цепи колец (катенаны) обычно образуются при репликации кольцевых молекул, но у прокариот специальные ферменты их расцепляют, так как у них дочерние клетки после деления должны получить по одной неповрежденной молекуле.
В матриксе митохондрий, поскольку там есть ДНК, то есть и собственный аппарат белкового синтеза. Собственные рибосомы митохондрий по многим характеристикам отличаются от эукариотических рибосом, которые находятся в цитоплазме тех же самых клеток. Но бóльшая часть белков, составляющих структурные и функциональные компоненты митохондрий, кодируется ядерным геномом, синтезируется на рибосомах эндоплазматической сети и транспортируется в митохондрии. На 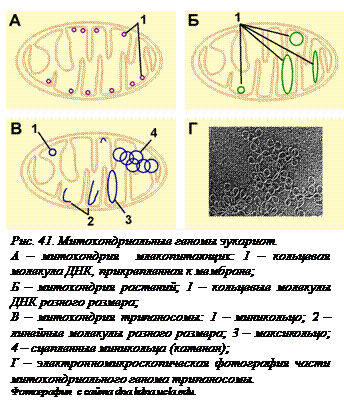 рибосомах же митохондрий образуется не более 5% от всех белков, входящих в их состав.
рибосомах же митохондрий образуется не более 5% от всех белков, входящих в их состав.
Размер генома митохондрий разных организмов колеблется от менее 6 тыс. пар нуклеотидов у малярийного плазмодия (в нем, помимо двух генов рРНК, содержится только три гена, кодирующих белки) до сотен тысяч пар нуклеотидов у наземных растений (например, у Arabidopsis thaliana из семейства крестоцветных 366 924 пар нуклеотидов). Длина мтДНК позвоночных животных отличается незначительно: у человека — 16 569 пар нуклеотидов, у дельфина — 16 330, у лягушки— 17 533, у карпа — 16 400. Эти геномы сходны также и по количеству и расположению генов, между которыми отсутствуют межгенные промежутки. В ряде случаев гены даже перекрываются, обычно на один нуклеотид, так что последний нуклеотид одного гена оказывается первым в следующем.
У большинства высших животных геном митохондрий содержит 37 генов: 13 для белков дыхательной цепи, 22 для тРНК и 2 для рРНК (рис. 39, А). Ключевые ферменты матричных синтезов, такие как ДНК-полимераза, осуществляющая репликацию митохондриальной ДНК, и РНК-полимераза, транскрибирующая геном митохондрий, кодируются в ядре и синтезируются на рибосомах цитоплазмы. Этот факт указывает на относительность автономии митохондрий в сложной иерархии эукариотической клетки.
Все мтДНК в отличие от ядерных и прокариотических ДНК, не метилируются. Известно, что метилирование – один из механизмов программируемой инактивации генов (см. § 28).По-видимому, это связано с тем, что митохондриальные гены не нужно выключать – они кодируют белки и РНК, которые необходимы для выполнения основной функции митохондрий – производства АТФ.
Транскрипция мтДНК (рис. 42, Б). В отличие от ядерного генома, митохондриальный геном очень компактен. В нем мало межгенных некодирующих областей. Это делает возможным транскрипцию полигенных РНК с каждой цепи мтДНК. Поэтому каждая из цепей мтДНК млекопитающих, представляющая собой полигенную структуру, транскрибируется с образованием одной молекулы РНК, начинающейся в районе ori H. Помимо этих двух длинных молекул РНК, комплементарных Н - и L -цепям, образуются и более короткие транскрипты Н -цепи, начинающиеся в той же точке и заканчивающиеся на 3’-конце гена 16 S рРНК. Этих коротких транскриптов в десять раз больше, чем длинных. В результате процессинга («созревания») из них образуются 12 S рРНК и 16 S рРНК, идущие на формирование митохондриальных рибосом, а также фенилаланиновая тРНК и валиновая тРНК. Из длинных транскриптов вырезаются остальные тРНК и образуются транслируемые иРНК, которые полиаденилируются по 3’-концам. Но 5’-концы этих иРНК не кепируются, что необычно для эукариот. Сплайсинга обычно не происходит, поскольку большинство митохондриальных генов млекопитающих не содержит интронов.
Репликация ДНК митохондрий млекопитающих (рис. 42, В). У большинства животных комплементарные цепи в мтДНК значительно различаются по удельной плотности в солевом растворе, поскольку имеют неодинаковое содержание «тяжелых» пуриновых и «легких» пиримидиновых нуклеотидов. Цепи так и называются – H (англ. heavy – тяжелая) и L (англ. light – легкая) цепь. В начале репликации молекулы мтДНК образуется так называемая D -петля (от англ. displacement loop – петля смещения). Эта структура, видимая в электронный микроскоп, состоит из двуцепочечного и одноцепочечного участков. Одноцепочечный участок – это отодвинутая часть Н -цепи. Двуцепочечный участок образован частью L -цепи и комплементарным ей вновь синтезированным фрагментом ДНК длиной 450-650 (в зависимости от вида организма) нуклеотидов, имеющим на 5’-конце рибонуклеотидную затравку. Этот конец соответствует точке начала синтеза Н -цепи (ori H).Синтез L -цепиначинается лишь тогда, когда дочерняя Н -цепь доходит до точки ori L. Это обусловлено тем, что область инициации репликации L -цепи доступна для ферментов синтеза ДНК лишь в одноцепочечном состоянии, а это имеет место только при расплетании двойной спирали при синтезе Н -цепи. Таким образом, в отличие от ядерной ДНК, митохондриальная ДНК реплицируется непрерывно по обеим цепям, но со значительном сдвигом во времени и в пространстве, так как участки ori в цепях далеко отодвинуты друг от друга.
Особенности митохондриального генома. В целом похожий на геном бактерий, митохондриальный геном обладает рядом особенностей. Например, некоторые гены митохондрий дрожжей содержит интроны. Так, в гене box (кодирующем цитохромоксидазу b) имеется два интрона. Но интроны далеко не всегда являются некодирующими участками. В данном случае те участки ДНК, которые представляют собой интроны для гена цитохромоксидазы b, для гена матуразы – фермента, участвующего в сплайсинге, играют роль экзонов. Из первичного РНК-транскрипта вырезается копия большей части первого интрона, причем происходит это автокаталитически, без участия каких-либо белков (пример автосплайсинга). Оставшаяся РНК служит матрицей для образования матуразы. Часть аминокислотной последовательности матуразы закодирована в оставшихся копиях интронов. Матураза вырезает их, разрушая свою собственную мРНК, копии экзонов сшиваются, и образуется мРНК для цитохромоксидазы b. Открытие этого феномена заставило пересмотреть представление об интронах, как о «ничего не кодирующих последовательностях».
Генетический код митохондрий млекопитающих несколько отличается от универсального значениями нескольких кодонов. Возможно, этим митохондрии «охраняют свою независимость», ведь митохондриальная мРНК не может быть прочитана цитоплазматическими рибосомами.
Геном хлоропластов. Так же как и митохондрии, хлоропласты обладают собственным геномом. ДНК хлоропластов кольцевая, Один хлоропласт обычно содержит несколько десятков копий генома. В настоящее время расшифрованы хлоропластные генома многих растений – кукурузы, табака, риса. В геномах хлоропластов содержится около 120 генов. Это гены всех рРНК, тРНК, рибосомных белков, часть генов, кодирующих белки фотосистем и электронно-транспортной цепи. При этом, как и в митохондриях, значительная часть белков или их субъединиц кодируется ядерным геномом.
Хлоропласты значительно сильнее, чем митохондрии, похожи на своих прокариотических предков (о происхождении митохондрий и пластид см. § 24). Например, часть генов хлоропластного генома объединена в опероны, имеющие одинаковые с прокариотами промоторы, терминаторы, некоторые регулирующие элементы, трансляция у них начинается не с метионина, а с формилметионина, как у бактерий. Но вместе с тем геному хлоропластов свойственны и некоторые черты, общие с эукариотиами. Так, в некоторых хлоропластных генах имеются интроны, есть не свойственный прокариотам процесс редактирования РНК.

