




Значительно сложнее устроены гены эукариот. Их структурные части состоят из чередующихся кодирующих и некодирующих участков – экзонов и интронов (рис. 31, А). Синтезированная по матрице ДНК РНК, содержащая копии интронов, подвергается сплайсингу – вырезанию интронов и сшиванию экзонов. Поэтому зрелая матричная РНК значительно короче считанной пре-мРНК. Большая часть эукариотических генов, кодирующих белки, содержат хотя бы один интрон. У человека средний ген, имеющий размер 27·103 пар нуклеотидов, содержит 8 интронов со средней длиной 1500 пар нуклеотидов. Однако некоторые гены, кодирующие белки, например, гены гистонов, гены некоторых трансмембранных рецепторов интронов не содержат. У млекопитающих таких генов 6 %, у плодовой мушки – 17 %, а у низших эукариот, таких как дрожжи, – 95 %. У низших всего несколько генов, кодирующих рибосомные и транспортные РНК, содержат по одному интрону. Регуляторные участки эукариотических генов значительно более протяженные, чем у прокариот. В среднем менее 5% гена несет информацию о последовательности аминокислот в полипептиде, остальная часть гена – это интроны и регуляторные участки.
Геном. Термин «геном» был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом одного биологического вида. В настоящее время термином «геном» обозначают совокупность молекул ДНК (не только генов) гаплоидного набора хромосом, которые заключены в отдельной клетке зародышевой линии многоклеточного организма или в одной прокариотической клетке. Можно говорить и о геноме митохондрии, хлоропласта, вируса.
Геномы вирусов. Самые разнообразные геномы имеют вирусы. Они могут быть представлены как ДНК, так и РНК. Молекулы могут быть одноцепочечными и двуцепочечными, линейными и кольцевыми. Количество генов может быть от всего 3 (фаг Qb) до 1 120 у самого крупного из известных в настоящее время вирусов – мегавируса, что в три раза больше, чем у некоторых бактерий. Соответственно и размер генома у вирусов может быть от менее 2000 нуклеотидов до >1,2·106 пар нуклеотидов. Геномы вирусов очень компактны, как правило, у них нет участков ДНК, не кодирующих белок. Для некоторых вирусов бактерий (фагов) показано перекрывание генов, когда один и тот же участок ДНК входит в состав двух генов. Белки они кодируют разные за счет сдвига рамки считывания (но в пределах одного гена кодоны не перекрываются! См. § 7). Сейчас к вирусам относят и самые маленькие инфекционные агенты – так называемые вироиды. Вироиды — это патогены растений, которые состоят из короткого фрагмента (несколько сотен нуклеотидов) кольцевой одноцепочечной РНК, не покрытой белковой оболочкой, характерной для вирусов. Самый маленький вироид scRNA имеет длину всего 220 нуклеотидов. Этого недостаточно даже для одного гена. Вироиды и не имеют собственных генов, но их РНК обладает свойствами рибозима, она способна к самоудвоению без участия белков. Для репликации РНК они используют клеточные мономеры. Поскольку у них нет оболочки, они не способны к существованию вне клетки.
Геномы прокариот представлены двуцепочечной кольцевой ДНК, прикрепленной к плазмалемме. Размеры прокариотических геномов – от 500 тыс. до 10 млн. пар нуклеотидов, количество генов – от 470 у самых мелких бактерий (микоплазм) до более 4000. Кроме основной молекулы ДНК бактерии могут содержать одну или несколько необязательных небольших кольцевых молекул – плазмид. Обычно плазмиды несут один или несколько генов, обеспечивающих устойчивость к различным факторам внешней среды – антибиотикам, солям тяжелых металлов. Геномы прокариот очень компактны, межгенные промежутки у них сведены до лидерных последовательностей, всего менее 15% нуклеотидных последовательностей ДНК не кодирует РНК и белки.
Геномы эукариот – самые большие и сложные. Даже у самых простых эукариот, дрожжей, геном уже значительно больше прокариотического – 12 млн. пар нуклеотидов, около 6000 генов.
У подавляющего большинства эукариот геном состоит из нескольких линейных молекул ДНК, соединенных со специальными белками – хромосом. В целом размеры геномов увеличиваются с усложнением организации живых существ. Исключение составляют лишь цветковые растения, некоторые виды которых имеют самые большие геномы. Внутри некоторых классов (рептилии, птицы, млекопитающие) диапазон размеров геномов мал, а внутри других (рыбы, амфибии) достигает двух и даже трех порядков. Между размером генома и числом хромосом никакой корреляции нет.
Все гены, входящие в геном, делят на две группы. Одна часть работает во всех клетках. Это гены, кодирующие белки рибосом, гистоны, структурные белки мембран и цитоскелета, ферменты, участвующие в биосинтезе биополимеров или в расщеплении веществ в реакциях энергетического обмена. Их называют генами «домашнего хозяйства». К ним относят и нетранслируемые гены транспортных и рибосомных РНК. Остальные гены, называемые «генами роскоши», транскрибируются и транслируются лишь ограниченное время и (или) не во всех клетках многоклеточного организма. Продукты этих генов нужны не производящей их клетке, а всему организму. Они производятся дифференцированными клетками, выполняющими функции, свойственные той или иной ткани. Так, например, ген инсулина работает лишь в производящих гормон клетках поджелудочной железы, а ген гормона роста – только в клетках гипофиза.
Когда определили размеры и структуру геномов эукариот, возникло такое понятие – «избыточность эукариотического генома». Действительно, на 1 ген Е. coli приходится в среднем около 1000 пар нуклеотидов, а у млекопитающих – около 160000 пар нуклеотидов. Почему это происходит? Во-первых, гены эукариот значительно больше прокариотических за счет наличия интронов и протяженных регуляторных областей. Во-вторых, некоторые гены в геноме многократно повторяются. Так, у млекопитающих имеется несколько сотен одинаковых генов рибосомных РНК, а у амфибий их несколько тысяч. В-третьих, в геномах эукариот присутствуют нефункциональные копии нормальных генов – так называемые псевдогены. Псевдогены не способны к кодированию и реализации полезной информации из-за изменений в их структуре, как правило, заключающихся в утрате важных фрагментов, например, промоторов. В-четвертых, в геномах эукариот много некодирующих последовательностей. Значительная часть их представляет собой обязательные структуры хромосом, отсутствующие у прокариот, такие как центромеры и теломеры, межгенные промежутки. Наконец, у эукариот значительно больше мобильных генетических элеменов, чем у прокариот (см. §§ 19, 20). В результате гены в геномах 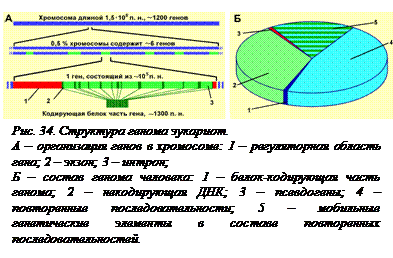 млекопитающих напоминают острова в океане некодирующих последовательностей (рис. 34).
млекопитающих напоминают острова в океане некодирующих последовательностей (рис. 34).
Своеобразен геном представителей третьего домена (кроме бактерий и эукариот) живых существ – архей. Хотя, как и у других прокариот, у архей геном представлен одной кольцевой молекулой ДНК, гены у них, как и у эукариот, имеют интроны. Структура промотора у них также ближе к эукариотам, чем к прокариотам. Исходя из этого именно археи считаются эволюционными предшественниками эукариот.
Геном человека. В каждой соматической (неполовой) клетке человека, имеющей ядро, содержится 46 хромосом – диплоидный (двойной) набор. Гаплоидный набор состоит из 23 хромосом. Результатом 15-летней работы международной программы «Геном человека», завершенной в 2003 году, стало определение последовательности всех 3,2·109 пар нуклеотидов, образующих геном. У человека почти в 5 раз больше генов, чем у кишечной палочки, а весь его геном больше генома E.coli почти в 700 раз. При сравнении с эукариотической дрозофилой (плодовой мушкой) цифры также различаются существенно – у человека в 1,5 раз больше генов, а весь геном в 25 раз больше дрозофилиного. Даже у позвоночного – рыбы фугу, геном меньше человеческого почти в 10 раз при практически одинаковом количестве генов. Большинство генов рыбы и человека если не совсем одинаковы, то очень похожи – у них сходная последовательность белок-кодирующей части генов, одинаковое количество экзонов. Отличаются они размерами интронов. Главное отличие генома фугу от геномов других позвоночных – очень небольшое количество и протяженность различных некодирующих последовательностей – межгенных промежутков, мобильных генетических элементов и др.
Было установлено, что генов у человека около 23 тысяч. Из 3,2×109 пар нуклеотидов генома человека только 25% занято белок-кодирующими генами, и только 1,5% из них – собственно кодирующие белок последовательности (экзоны). Что же представляют собой остальные 75% генома человека (рис. 31, Б)? В основном эта часть генома занята так называемыми повторенными последовательностями. Они представляют собой многократно повторяющиеся участки ДНК с одинаковой первичной структурой. Примерно 2% генома человека составляют высокоповторенные (повторенные более 105 раз) последовательности, составленные повторяющимся блоком длиной в 171 п.н. Эти повторы находятся в центромерных участках каждой хромосомы. С этими участками связываются специальные белки, составляющие кинетохор (см. § 20). На концах хромосом находятся теломерные повторы (см. § 15). Эти повторенные последовательности генов не содержат.
Умеренно повторенные (менее 105 раз) последовательности включают в себя блоки генов «домашнего хозяйства», продукты которых клеткам нужны постоянно в большом количестве (гены гистонов, рРНК, тРНК, белков цитоскелета). Потребное количество этих продуктов один даже очень активно транскрибируемый и транслируемый ген обеспечить не может.
Не менее 50% генома человека занимают псевдогены и мобильные генетические элементы (МГЭ), которые иногда называют прыгающими генами. О них будет подробнее рассказано в § 20.
Геномика. Конец ХХ – начало XXI века охарактеризовался расшифровкой геномов не только человека, но и мыши, собаки, коровы и многих других животных. Возникло новое направление в науке – геномика. Сравнение нуклеотидных последовательностей в геномах разных видов организмов позволяет делать выводы об их родственных отношениях, эволюционной истории. Однако расшифровать геном – это только его прочитать, но не понять. Среди прочитанной последовательности надо выделить гены и некодирующие области и, самое главное, понять их функции. Компьютерный анализ позволяет находить области, похожие на гены (так называемые открытые рамки считывания), а также области, содержащие промоторы (ТАТА и ЦААТ-боксы), сигналы сплайсинга, сигналы полиаденилирования и т.д. Это может служить признаком того, что в этой области находится ген, кодирующий белок. Сравнение нуклеотидной последовательности такого участка с известными генами хорошо изученных видов позволяет делать предположение о том, что именно кодирует данный ген. Разработка методов «нокаутирования» генов с помощью антисмысловых или интерферирующих РНК (см. § 13) позволила создать так называемую «обратную генетику». Смысл ее заключается в том, что выключается ген с известной нуклеотидной последовательностью, но неизвестной функцией. После этого в таких «нокаутированных» клетках или организмах лабораторных животных определяют, какая функция нарушается, какого белка не хватает. Это и позволяет определить функцию данного гена. В настоящее время перед международным научным сообществом стоит задача определения функций всех 23000 генов человека.
§ 18. Компактизация ДНК.
Как мывидели, геномы даже самых мелких прокариот содержат миллионы пар нуклеотидов. Хотя размер одной нуклеотидной пары составляет всего 0,34 нм, длина геномной ДНК кишечной палочки составит 1,6 мм при длине самой бактерии 1-6 мкм. У эукариот геномы значительно больше. Суммарная длина всех 46 молекул ДНК человека составляет около 2 метров, а у некоторых лилейных – более 10 м! Каким же образом эти громадные молекулы умещаются в ядре диаметром менее 10 мкм? Для этого ДНК должна быть плотно упакована, при этом она не должна запутываться, и любой ее участок должен быть доступен для ферментов или транскрипционных факторов. Кроме того, молекулы ДНК такой длины не могут существовать сами по себе – они просто разорвутся под действием собственной тяжести. Поэтому ДНК никогда не бывает свободной от белков. Кроме ферментов и разнообразных регуляторных белков, это специальные упаковочные белки, обеспечивающие компактизацию (упаковку) ДНК в ядре клетки. Такой комплекс ДНК с упаковочными белками называют хроматином, а одна молекулы ДНК в комплексе с белками называется хромосомой.
Существует несколько уровней упаковки ДНК (обычно считается, что их 4, но некоторые ученые насчитывают 5). Два первых уровня компактизации эукариотического генома обеспечиваются белками, которые называются гистонами.
Гистоны – оснóвные белки. Все они обогащены лизином и аргинином – положительно заряженными аминокислотами. Благодаря этому они электростатически взаимодействуют с ДНК, являющейся полианионом. Выделяют пять видов гистонов, обозначаемых Н1, Н2А, Н2В, Н 3 и Н4. Синтезируется их очень много – 60 млн. молекул каждой фракции на клетку.
Все гистоны, кроме Н1, чрезвычайно консервативны в эволюционном отношении (у коровы и клевера гистоны Н2А отличаются всего одной аминокислотой!). Это значит, что эти белки выполняют важную функцию, которая у всех эукариот обеспечивается одинаково. Практически любая мутация в гистоновых генах летальна.
Н1 – очень вариабельная фракция. Этот гистон различен не только у видов, но даже у одного организма, в зависимости от стадий онтогенеза.
На первом уровне компактизации двойная спираль ДНК с диаметром 2 нм как на катушку наматывается на белковый комплекс, содержащий 8 молекул гистонов – гистоновые октамеры (рис. 35, А, Б). Образуется структура, напоминающая бусы на нитке, с диаметром 11 нм (рис. 35, В). В состав сердцевины каждой такой бусины – нуклеосомы, входит по две одинаковых молекулы четырех разных гистонов (Н2А, Н2В, Н 3 и Н4). Они образуют кор (сердцевину) нуклеосомы. На кор наматывается ДНК – 1,75 левых витка спирали. Устье этих витков замыкается молекулой гистона Н1. Непосредственно с октамером контактирует 145 пар нуклеотидов, и 20-40 пар нуклеотидов остается между нуклеосомными корами. Нуклеосомная нить короче неупакованной молекулы ДНК в 7 раз. Молекула диаметром 2 нм упаковывается в нить диаметром 11 нм.
На втором уровне компактизации нуклеосомы сближаются с помощью гистона Н1. Образуется фибрилла с диаметром 30 нм (рис. 35, Г-Ж), а общая длина уменьшается еще в 6-10 раз.
На третьем уровне упаковки формируются петли, содержащие от 20 до 80 тыс. пар нуклеотидов (рис. 35, З). Петли «подвешиваются» на ядерных белках (третий уровень компактизации), образующих так называемый «скэффолд» (англ. scaffold – опора). В «устье» каждой петли находятся белки, которые узнают определенные нуклеотидные последовательности и при этом имеют сродство друг к другу. Типичная хромосома млекопитающих может содержать до 2500 петель. Как правило, петли суперспирализованы. Частично расправляются те петли, в которых расположены активно транскрибирующиеся в данный момент гены. Негистоновые белки и гистоны вместе с ДНК 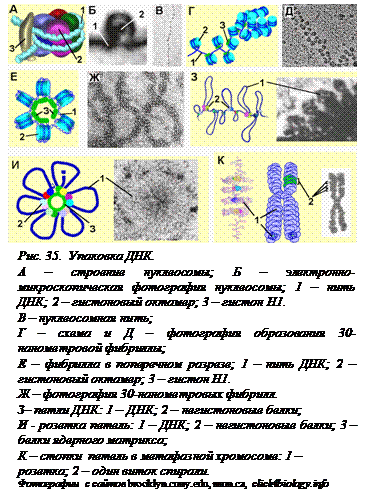 образуют хроматин, видимый лишь в электронный микроскоп. Среди негистоновых белков хроматина есть не только белки, участвующие в упаковке ДНК, но и ферменты, осуществляющие транскрипцию, репликацию и репарацию – восстановление структуры ДНК после повреждения.
образуют хроматин, видимый лишь в электронный микроскоп. Среди негистоновых белков хроматина есть не только белки, участвующие в упаковке ДНК, но и ферменты, осуществляющие транскрипцию, репликацию и репарацию – восстановление структуры ДНК после повреждения.
Перед делением клетки молекулы ДНК удваиваются, и в них возникает четвертый уровень упаковки, за счет чего хромосома утолщается и становится видимой в световой микроскоп. На этом уровнеупаковки структура, получившаяся на предыдущем уровне, свертывается в спираль, образуя розетки (рис. 35, И). Затем розетки петель складываются встопки, которые можно видеть в световой микроскоп (рис. 35, К). В целом «укорочение» ДНК достигает 104. Это соответствует тому, как если бы тонкую нитку длиной в полкилометра упаковали бы в спичечный коробок (5 см).
Если из такой компактизованной хромосомы удалить гистоны, белки скэффолда будут продолжать скреплять петли ДНК, но сами петли при этом расправятся и окружат «скелет» хромосомы плотным облаком, во много раз большим, чем метафазная хромосома.Это наглядно показывает, как плотно упакована ДНК в хромосомах (рис. 36, А).
 У бактерий гистонов нет, но ДНК связана с положительно заряженными полиаминами и ионами Mg 2+. Поэтому у бактерий нет нуклеосом. Однако их ДНК, по длине в 1000 превышающая размеры самих бактериальных клеток также компактизована. Вся кольцевая молекула организована в 20-100 (в зависимости от размера генома) суперспирализованных петель. Эти петли можно увидеть после обработки клеток растворами, удаляющими часть белков (рис. 33, Б). В обеспечении суперспирализации участвуют молекулы малых РНК.
У бактерий гистонов нет, но ДНК связана с положительно заряженными полиаминами и ионами Mg 2+. Поэтому у бактерий нет нуклеосом. Однако их ДНК, по длине в 1000 превышающая размеры самих бактериальных клеток также компактизована. Вся кольцевая молекула организована в 20-100 (в зависимости от размера генома) суперспирализованных петель. Эти петли можно увидеть после обработки клеток растворами, удаляющими часть белков (рис. 33, Б). В обеспечении суперспирализации участвуют молекулы малых РНК.
У архей, имеющих сходство с эукариотами в строении генов, обнаружены гистоны и гистоноподобные белки, связанные с ДНК, и, соответственно, нуклеосомы. Но в целом о компактизации ДНК архей известно мало.
§ 19. Хромосомы и кариотипы.
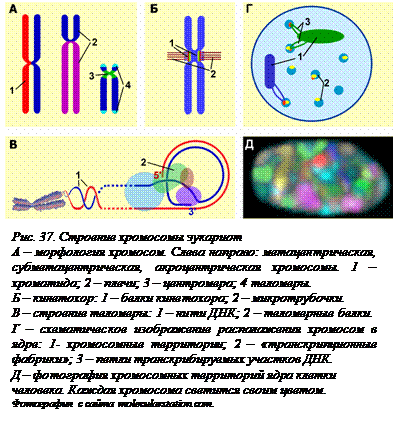 В ядрах клеток эукариот находится не одна кольцевая молекула ДНК, как у прокариот, а несколько линейных молекул, связанных с белками. Эти образования и представляют собой хромосомы. Декомпактизованные хромосомы обычно не видны в световой микроскоп, но во время деления клетки, когда они дополнительно компактизуются, становятся короче и толще, их можно видеть. Поскольку перед делением они обязательно удваиваются, то и в микроскоп они видны двойными, состоящими из двух хроматид. Хроматиды соединены в одном участке, но соединяются в нем не молекулы ДНК, а специальные белки – когезины.[O30] Эти белки по своей форме напоминают английскую булавку, которой хроматиды «пристегнуты» друг к другу. Этот участок соединения хроматид носит название центромеры. Разные хромосомы могут отличаться друг от друга положением центромеры. В метацентрических хромосомах центромера находится посередине, в субметацентрических она сдвинута к одному из концов, в акроцентрических находится почти на самом конце, рядом с теломерой (рис. 37, А). Часть хромосомы от центромеры до теломеры называют плечом. Хромосомы одной клетки различаются и по положению центромеры и по длине. У человека самая мелкая хромосома в 5 раз короче самой длинной.
В ядрах клеток эукариот находится не одна кольцевая молекула ДНК, как у прокариот, а несколько линейных молекул, связанных с белками. Эти образования и представляют собой хромосомы. Декомпактизованные хромосомы обычно не видны в световой микроскоп, но во время деления клетки, когда они дополнительно компактизуются, становятся короче и толще, их можно видеть. Поскольку перед делением они обязательно удваиваются, то и в микроскоп они видны двойными, состоящими из двух хроматид. Хроматиды соединены в одном участке, но соединяются в нем не молекулы ДНК, а специальные белки – когезины.[O30] Эти белки по своей форме напоминают английскую булавку, которой хроматиды «пристегнуты» друг к другу. Этот участок соединения хроматид носит название центромеры. Разные хромосомы могут отличаться друг от друга положением центромеры. В метацентрических хромосомах центромера находится посередине, в субметацентрических она сдвинута к одному из концов, в акроцентрических находится почти на самом конце, рядом с теломерой (рис. 37, А). Часть хромосомы от центромеры до теломеры называют плечом. Хромосомы одной клетки различаются и по положению центромеры и по длине. У человека самая мелкая хромосома в 5 раз короче самой длинной.
Во время деления клетки на центромере формируется особая белковая структура – кинетохор (рис. 37, Б). Это сложный комплекс из многих белков. К кинетохору присоединяются идущие от клеточного центра микротрубочки, служащие «рельсами», по которым моторные белки кинетохора двигают хромосомы к полюсам клетки во время ее деления.
На конце молекул ДНК находятся не содержащие генов теломерные повторы – многократно повторяющиеся последовательности нуклеотидов. При этом 3’-конец молекулы ДНК всегда выступает, то есть имеет однонитевой участок, образующийся в результате недорепликации 3’-конца матрицы (см. § 15). Эти участки защищаются от действия нуклеаз специальными теломерными белками. С помощью этих белков конец молекулы закручивается в кольцо (рис. 37, В). Самый кончик при этом образует триплекс – тройную нить. Теломеры защищают кодирующие последовательности ДНК – гены, от разрушения ферментами.
В неделящихся клетках, когда нет последнего уровня компактизации, ДНК, тем не менее, не просто заполняет внутреннее пространство ядра, хромосомы при этом занимают в ядре определенную компактную территорию. Предполагается, что те части хромосомы, где находятся неработающие гены или вообще нет генов, спрятаны внутрь этой хромосомной территории, а активно транскрибирующиеся районы декомпактизуются и в виде петель находятся в относительно свободных местах (рис. 37, Г, Д). При этом теломеры и некоторые другие участки хромосом закреплены на внутренней мембране ядра, а области хромосом, обычно соответствующие границам петель ДНК (третий уровень компактизации), закреплены на так называемом ядерном скелете. Определение белков, входящих в его состав, показало, что там находятся ферменты – ДНК-полимераза, топоизомераза, РНК- полимераза и некоторые другие. По-видимому, они представляют собой «транскрипционные фабрики», куда и протягиваются петли [O31] активно считываемой ДНК. По современным представлениям при транскрипции и репликации не ферменты движутся по нити ДНК, а ДНК протягивается через эти белковые комплексы.
|
Таблица 5. Числа хромосом (2 n) некоторых организмов | |||
| Вид | 2 n | Вид | 2 n |
| дрожжи | 32 | кошка | 38 |
| лошадиная аскарида | 2 | собака | 78 |
| дрозофила | 8 | носорог | 84 |
| окунь | 48 | человек | 46 |
| лягушка | 36 | лук | 16 |
| курица | 78 | бобы | 12 |
| мышь | 40 | кукуруза | 20 |
| барабинский хомячок | 20 | горох | 14 |
Полный упорядоченный набор хромосом клетки на стадии метафазы митоза называют кариотипом (греч. καριον – ядро, τυπος – форма). Количество хромосом и их морфология в пределах одного кариотипа постоянны. У организмов, размножающихся половым путем, соматические клетки обычно содержат два набора хромосом – один, полученный от отца, другой – от матери. Поэтому количество хромосом в кариотипе принято обозначать символом 2n (табл. 5). Кариотипы разных видов организмов могут различаться по числу, размерам и форме хромосом, и служат своеобразным паспортом вида. Организмы разных видов могут иметь одинаковое число хромосом, однако полностью идентичные кариотипы встречаются весьма редко и только у близких видов.
В некоторых случаях хромосомы одного кариотипа трудно отличить друг от друга по внешнему виду. На рис. 38 приведены фотографии кариотипа одного из видов полевок. Хромосомы №№ 8-24 у этой полевки одноплечие, и с помощью обычной окраски отличить, например, хромосому № 13 от хромосомы № 14 или № 16 невозможно. В этих случаях помогают специально разработанные методы дифференциальной окраски хромосом. Обработка препаратов хромосом с помощью протеолитического фермента трипсина приводит к удалению части хромосомных белков, в результате окраска хромосом становится полосатой. Распределение и толщина полос строго постоянны. Это не только позволяет «различать в лицо» хромосомы одного вида, но и  сравнивать между собой хромосомы, принадлежащие разным видам, определять их родственные связи, различать близкие виды. Дело в том, что рисунок «полосатости» определяется генетическим содержимым данного участка хромосомы и сохраняется при перестройках хромосом.[O32]
сравнивать между собой хромосомы, принадлежащие разным видам, определять их родственные связи, различать близкие виды. Дело в том, что рисунок «полосатости» определяется генетическим содержимым данного участка хромосомы и сохраняется при перестройках хромосом.[O32]
Кариотипы разных видов млекопитающих обычно сильно различаются между собой, но, поскольку предок у всех млекопитающих общий, то и хромосомы их между собой тоже «родственники». В ходе эволюции кариотипы подвергались многочисленным перестройкам, при этом менялось взаимное расположение разных участков хромосом. Но отдельные участки хромосом (а у близких видов и целые хромосомы) 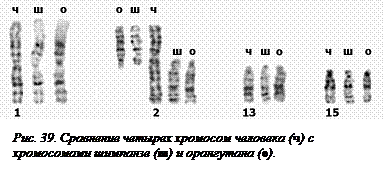 [O33] сохранили взаимное расположение и генов и рисунков окраски. На рис. 39 приведены фотографии хромосом №№ 1, 2, 13 и 15 человека в сравнении с соответствующими хромосомами близких видов – шимпанзе и орангутана. Видно, что хромосоме 2 человека соответствуют по две соединенных в области центромеры хромосомы обезьян. При этом общий рисунок окраски у них одинаков. Остальные три хромосомы одинаковы у всех трех видов.
[O33] сохранили взаимное расположение и генов и рисунков окраски. На рис. 39 приведены фотографии хромосом №№ 1, 2, 13 и 15 человека в сравнении с соответствующими хромосомами близких видов – шимпанзе и орангутана. Видно, что хромосоме 2 человека соответствуют по две соединенных в области центромеры хромосомы обезьян. При этом общий рисунок окраски у них одинаков. Остальные три хромосомы одинаковы у всех трех видов.
§ 20. Мобильные генетические элементы и псевдогены. Эволюция геномов.
Как было сказано в § 17, значительную часть геномов составляют мобильные генетические элементы (иногда называемые «прыгающими генами») и псевдогены. Долгое время они считались «генетическим мусором» или геномными паразитами. В настоящее время установлено, что, несмотря на то, что некоторые из них, действительно, можно назвать паразитами, они могут играть весьма важную роль в обеспечении некоторых функций организма и в эволюции.
Мобильные генетические элементы (или транспозоны) представляют собой небольшие участки ДНК, способные перемещаться по геному, встраиваясь в разные его участки. Они были открыты в 50-х годах ХХ века американским генетиком Барбарой Мак-Клинток, описавшей мозаичность окраски зерен у кукурузы, необъяснимую законами Менделя и мутационной теорией. Она предположила, что некоторые гены могут менять свое местоположение в геноме. Вначале к ее открытию отнеслись скептически, а через 50 лет присудили за него Нобелевскую премию. Сейчас МГЭ обнаружены у всех живых организмов – и у бактерий, и у архей, и у всех эукариот.
Все МГЭ делят на два класса – ретроэлементы и более малочисленные ДНК-траспозоны. Самый простые ДНК-транспозоны – так называемые ІЅ - элементы, их размер около 1000 п.н. На концах они содержат инвертированные (палиндромные) повторы (~20 п.н.) (рис. 40, А, I). IS -элементы содержат только один ген – ген транспозазы, фермента, обеспечивающего перемещение IS -элемента по геному. Транспозаза вырезает участок ДНК в одном месте и вставляет его в другое место генома. Перед геном в IS -элементе имеется промотор, за геном – слабый терминатор транскрипции. РНК-полимераза останавливается на нем не всегда, она может продолжать транскрибировать и расположенный рядом участок генома до сильного терминатора. Это может привести к тому, что в клетке «включится» «молчащий» ген. IS -элементы обнаруживаются в основном у прокариот и бактериофагов, но найдены и у некоторых растений.
Tn -транспозоны содержатся в геномах как прокариот, так и эукариот. Помимо гена транспозазы в них находится один или несколько структурных генов (рис. 40, А, II). Транспозоны прокариот чаще всего содержат так называемые гены лекарственной устойчивости. Эти гены кодируют ферменты, расщепляющие антибиотики. Количество копий транспозона в клетках бактерий возрастает при наличии в среде антибиотика, ген устойчивости к которому кодируется в данном транспозоне. Существуют транспозоны, не содержащие гена транспозазы. Такие транспозоны содержат по краям IS -элементы, за счет транспозазы которых они и перемещаются. Транспозоны могут вырезаться из основной геномной ДНК бактерии и встраиваться в плазмиды. Транспозоны эукариот могут содержать гены, кодирующие белки, облегчающие перемещение по геному (например, особые ДНК-связывающие белки). Активно перемещающиеся транспозоны обнаружены у многих растений, а также у животных от беспозвоночных до амфибий. В геноме человека также обнаружены ДНК-транспозоны, однако они неактивны, так как в них накопилось много мутаций. Полагают, что геномы млекопитающих вообще не содержат активных ДНК-транспозонов.
К классу ДНК-транспозонов прокариот относят плазмиды. Плазмида – это маленькая кольцевая молекулы ДНК у бактерий. Она меньше геномной ДНК в тысячи раз. Плазмиды способны к автономной репликации, независимой от репликации основной молекулы ДНК. Некоторые плазмиды, в которые встроены определенные транспозоны, способны встраиваться (интегрироваться) в геномную ДНК. Такие плазмиды называют эписомами.
После перехода из основной ДНК в плазмиду транспозон может попасть с ней в другую бактерию, придавая новому хозяину ранее отсутствовавшую лекарственную устойчивость. Кроме того, при вырезании транспозона из геномной бактериальной ДНК могут захватываться участки генома одной бактерии, которые вместе с транспозоном переносятся в другую бактерию. Плазмиды и эписомоподобные факторы есть и у некоторых эукариот, например, у дрожжей.
Некоторые вирусы животных и фаги прокариот фактически тоже являются мобильными элементами. ДНК так называемых умеренных фаг ов (вирусов бактерий, не вызывающих их гибели) может встраиваться в бактериальный геном, размножаться вместе с бактерией, а затем вырезаться, упаковываться в белковую оболочку и в виде фаговых частиц заражать другие бактерии. Например, λ-фаг может захватывать часть генетической информации одной бактерии и переносить ее в другую. У прокариот это происходит очень часто, поэтому иногда говорят о едином генофонде прокариот. Некоторые ДНК-содержащие вирусы животных также могут находиться в геноме клетки хозяина в форме провируса на протяжении нескольких поколений, а потом начать перемещаться.

