




§ 22. Живые системы.
Во Введении уже говорилось, что живая материя представляет собой сложную систему с особыми свойствами. Можно повторить, что с истемой называют множество элементов, находящихся в определенных отношениях друг с другом и связанных прямыми и обратными связями, образуя целостность, единство. Наличие связей между элементами системы – это, говоря другими словами, обмен информацией.
Системы могут быть простыми и сложными. Элементами сложных систем являются другие системы (подсистемы), причем для работы системы высшего уровня, в общем, не важно, как устроены и как работают подсистемы, важна только информация, импульсы, получаемые от систем низшего уровня. Например, взаимодействие популяций разных видов организмов в пределах экосистемы не зависит от структуры макромолекул, из которых построены эти организмы, а только от их взаимоотношений – пищевых цепей, пространственного расположения, численности. Но главное отличие между простыми и сложными системами заключается в том, что поведение простых систем однозначно определяется тем или иным алгоритмом. Поведение сложной системы включает возможность выбора альтернативных путей функционирования. Это принципиально важно – для сложной системы невозможно однозначно предсказать ее состояние через какое-то время исходя только из начальный параметров.
Живые системы – одни из самых сложных в природе. Например, протеом (система всех белков и их взаимодействий) одного из самых простых эукариотических организмов – дрожжей включает в себя 1458 белков и 1948 взаимодействий между ними. Каждый белок кодируется не менее чем одним геном. Средний белок состоит примерно из 400 аминокислот, чему соответствует более 1200 пар нуклеотидов (рис. 43, А). Нуклеотид – сложная молекула, состоящая из 32-34 атомов. Сравните с относительно простой физической системой – молекулой воды, состоящей всего из трех атомов (рис. 43, Б). Именно для живой материи характерно возникновение, передача и реализация огромного количества информации.
Большинство из свойств, которыми обычно описывают живое (дискретность, иерархичность устройства, обмен веществ и энергии, обмен информацией, способность к развитию, адаптивность, раздражимость, поддержание гомеостаза, самовоспроизведение) являются на самом деле системными свойствами, характерными для самых разных сложных открытых систем. Системы изучают разные отрасли науки. Это могут быть физические, химические, биологические, технические (машины), социальные системы. Самое замечательное, что поведение этих систем зависит не от того, к какой сфере они относятся, а от их организации – количества и связей элементов. Поэтому свойства самых разных систем оказываются очень близкими. Эти так называемые системные свойства изучает наука общая теория систем, основоположником которой является австрийский биолог Людвиг фон Берталанфи.
Принципиальное свойство живых систем – то, что они являются открытыми. Именно на это указывают такие признаки живого, как обмен веществ, энергии, информации. Закрытые системы, которые изучала классическая термодинамика, находятся в равновесии или в состоянии, близком к равновесию. При этом происходит выравнивание термодинамических потенциалов в системе, то есть выравнивание температуры, концентраций и т.п. Сложные структуры, образованные с большой затратой энергии, то есть, неравновесные, при этом разрушаются. Другими словами, увеличивается энтропия. Можно это выразить и так – термодинамические изменения закрытой системы всегда идут в сторону наиболее вероятных ее состояний и, в конечном счете, ведут к состоянию хаоса – максимальной выравненности и симметрии.
В открытых системах, в частности, живых организмах – все наоборот. Выравнивания температуры не происходит (нагреваются не только теплокровные, но и все остальные, включая растения), вещества распределены крайне неравномерно, сложные структуры постоянно создаются, энтропия живой системы уменьшается. Знаменитый физик Шредингер выразил это афоризмом «Жизнь есть наиневероятнейшее состояние материи». Другими словами, живые системы находятся в состоянии, далеком от термодинамического равновесия, и эта неравновесность сохраняется, пока сохраняется жизнь. Одно из определений жизни, принадлежащее фон Берталанфи, гласит, что «жизнь – процесс поддержания неравновесного состояния извлечением энергии из внешней среды». Поскольку, эта неравновесность может поддерживаться только за счет притока энергии из внешней  среды, в целом энтропия Вселенной не уменьшается и второе начало термодинамики остается незыблемым. Это позволило дать Шредингеру «термодинамическое» определение жизни «Жизнь… – это работа специальным образом организованной системы, направленная на понижение собственной энтропии за счет повышения энтропии окружающей среды». Одно из наиболее полных определений живого принадлежит отечественному биофизику М. В. Волькенштейну: «Живые тела, существующие на Земле, представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров – белков и нуклеиновых кислот».
среды, в целом энтропия Вселенной не уменьшается и второе начало термодинамики остается незыблемым. Это позволило дать Шредингеру «термодинамическое» определение жизни «Жизнь… – это работа специальным образом организованной системы, направленная на понижение собственной энтропии за счет повышения энтропии окружающей среды». Одно из наиболее полных определений живого принадлежит отечественному биофизику М. В. Волькенштейну: «Живые тела, существующие на Земле, представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров – белков и нуклеиновых кислот».
Теорию поведения неравновесных систем (нелинейную термодинамику) разработал бельгийский ученый, нобелевский лауреат Илья Пригожин. Система, в которой что-либо происходит, естественно, меняет свои состояния. Такая последовательность состояний системы называется траекторией системы. Траектории закрытых систем предсказываются равновесной термодинамикой – они представляют собой монотонное возрастание энтропии и достижение равновесного состояния. Траектории неравновесных систем могут быть самыми разными – однозначно они не определяются. Однако среди них одни более вероятны, другие менее, то есть состояние системы через некоторое время после начала наблюдения определяется в терминах теории вероятностей. Состояние неравновесной системы совсем не обязательно должно меняться быстро. В наиболее вероятном состоянии система может находиться долго и устойчиво (то есть пребывать в стационарном состоянии, поддерживая его за счет притока энергии из внешней среды). Такие наиболее вероятные состояния и траектории системы в специальном разделе математики называются аттракторами (от англ. attract – притягивать). При незначительном случайном отклонении траектории система обычно возвращается в прежний аттрактор. Но траектории сильно неравновесных систем могут быть очень зависимы от таких флуктуаций – случайных отклонений какой-либо величины, характеризующей систему, от наиболее вероятного значения. В результате такой случайности система может скачком перейти в другой аттрактор – состояние, очень сильно отличающееся от прежнего. Следует иметь в виду, что аттракторы – это не состояния равновесия, а наиболее вероятные стационарные состояния неравновесной системы. Другими словами, одни неравновесные состояния системы предпочтительнее других. А это уже означает возникновение некоторой упорядоченности, то есть убывание энтропии. Это явление Пригожин назвал самоорганизацией в неравновесных системах.
Один из простейших случаев спонтанной самоорганизации в физической системе – неустойчивость Бенара, возникающая при нагревании снизу слоя вязкой жидкости, когда при определенной разности температур нижнего и верхнего слоев жидкости в ней образуются правильные структуры (конвективные ячейки правильной шестиугольной формы). Такие ячейки иногда возникают и в воздухе (приводя к появлению шестиугольных отпечатков на песке пустынь, арктических снегах и на правильно «расчесанных» полях злаков, что часто приписывают «пришельцам»), при таянии мерзлого грунта и в расплавленной лаве (рис. 43, В, Г).
Простым примером самоорганизующейся системы является сборка (часто так и говорят – самосборка) фосфолипидных мембран в водной среде. Молекулы фосфолипидов являются амфифильными – у них гидрофильная «голова», представляющая собой молекулу глицерина с остатком фосфорной кислоты, и длинные гидрофобные углеводородные «хвосты» – остатки жирных кислот. Если поместить фосфолипидные молекулы в воду, то гидрофильные «головы» будут растворяться в воде, а гидрофобные «хвосты» раствориться в воде не могут. Поэтому они будут препятствовать проникновению молекул в толщу воду. Результатом будет образование упорядоченной структуры – мономолекулярной пленки. Если молекулы фосфолипида окажутся в замкнутом объеме воды, где нет поверхности, то гидрофобные «хвосты», стремясь «спрятаться» от воды, образуют двухслойную пленку, замкнутую в пузырек – липосому (рис. 43, Д). В таком пузырьке с водой внутри и снаружи соприкасаются только гидрофильные «головы». Поэтому и клеточные мембраны, построенные из фосфолипидов, тоже могут «самособираться». Липосомы широко используют для доставки лекарственных веществ прямо в клетки. Лекарство при этом не попадает в кровь или другие жидкости тела – этому препятствует мембрана. А в клетки содержимое липосомы попадает легко, так как ее мембрана аналогична клеточной и легко сливается с ней.
Еще одним примером самопроизвольного образования упорядоченных структур является образование третичной структуры белка (см. § 1). Длинная полипептидная молекула в водной среде свертывается, так как гидрофобные радикалы аминокислот сближаются друг с другом, образуя плотную глобулу, максимально защищенную от контактов с гидрофильной средой (рис. 43, Е).
Другой пример самоорганизации, изученный Пригожиным – реакция Белоусова-Жаботинского (окисление малоновой кислоты бромноватистым калием, катализируемое солями церия), так называемые «химические часы», когда реакционная смесь внезапно через равные промежутки времени меняет цвет с ярко-красного на чисто-синий и наоборот. Как показали исследования, для этого в системе должны присутствовать автокаталитические циклы, то есть такие стадии, в которых продукт реакции катализирует синтез самого себя (рис. 44, А). [O34] Фактически это означает самовоспроизведение системы. Такие простые автокаталитические системы редки. Обычно это не одна реакция, а реакционный цикл, включающий большое количество промежуточных продуктов. Например, в реакции Белоусова-Жаботинского присутствует около 30 промежуточных реакций. [O35]
Детальный анализ ячеек Бенара и реакции Белоусова-Жаботинского, проведенный Пригожиным, показал, что по мере того, как система движется все дальше от равновесия, она достигает критической точки нестабильности, в которой скачком возникают организованные структуры. Другими словами, согласованное действие миллиардов молекул возникает, когда степень хаотичности достигает определенной критической величины. Такое поведение систем описывается нелинейными уравнениями, поэтому этот раздел физики называют нелинейной термодинамикой.
Подробно изучал неравновесные химические системы немецкий ученый Манфред Эйген, Нобелевский лауреат в области химии. Эйген исследовал ферментативные системы, в которых различные взаимосвязанные каталитические реакции образовывали сложные циклы и сети, содержавшие обратные связи. Он обнаружил, что автокаталитические циклы, подобные рассмотренным выше, могут взаимодействовать друг с другом, если продукты превращения субстрата катализируют не только реакции «своего» цикла, но и другие реакции. Несколько таких автокаталитических циклов, если в них участвуют общие промежуточные продукты, могут организовываться в систему более высокого уровня. Эйген назвал эти системы гиперциклами.
Простейшим примером природного гиперцикла является размножение РНКового вируса в клетке. С попавшей в клетку РНК вируса транслируется фермент – РНК-зависимая РНК-полимераза. Фермент реплицирует РНК, которая одновременно является матрицей и для информационной РНК этого же фермента, и геномной РНК для новых вирусных частиц. С новых молекул РНК снова транслируется РНК-полимераза, которая тоже реплицирует РНК. Результатом является экспоненциальное размножение вируса. Очень важно отметить, что подобные циклы из-за наличия  положительных обратных связей являются самоусиливающимися, они развиваются экспоненциально. Поэтому очень быстро возникает дефицит субстрата. Это создает отрицательную обратную связь – замедляет образование продуктов. В результате ускорение и замедление компенсируют друг друга. Другими словами, автокаталитические циклы обладают свойством саморегуляции. Важно понять, что слова «обратная связь» обозначают не что иное, как передачу информации. Таким образом, автокаталитические циклы способны создавать и передавать информацию, самовопроизводиться и саморегулироваться.
положительных обратных связей являются самоусиливающимися, они развиваются экспоненциально. Поэтому очень быстро возникает дефицит субстрата. Это создает отрицательную обратную связь – замедляет образование продуктов. В результате ускорение и замедление компенсируют друг друга. Другими словами, автокаталитические циклы обладают свойством саморегуляции. Важно понять, что слова «обратная связь» обозначают не что иное, как передачу информации. Таким образом, автокаталитические циклы способны создавать и передавать информацию, самовопроизводиться и саморегулироваться.
Одним из наиболее поразительных свойств таких гиперциклов является их способность, проходя через состояния нестабильности в ходе развития, спонтанно создавать более высокие уровни организации. Гиперциклы, не являясь живыми организмами, обладают некоторыми ключевыми свойствами живого. В частности, они способны к эволюционному поведению по Дарвину. При условии дефицита субстрата (а мы видели, что количество субстрата всегда ограничено в силу экспоненциального роста гиперцикла) разные гиперциклы, если они используют общие субстраты, начинают конкурировать, в результате чего возникает естественный отбор. Более «медленные» гиперциклы могут «вымереть». Здесь следует отметить, что дарвиновская эволюция с естественным отбором и наследственной изменчивостью – свойство определенного класса систем, которые мы рассматриваем, а именно сложных открытых систем с обратными связями и самовоспроизведением элементов. Главное, что эти системы не обязаны быть биологическими! Дарвин, сам того не подозревая, открыл универсальное свойство систем. Одним из проявлений этого свойства у только одного класса систем – биологического, и является эволюция (усложнение самовоспроизводящейся системы) путем естественного отбора. Более того, именно дарвиновская эволюция (случайные флуктуации, отбор, закрепление в памяти, или, другими словами, наследственная изменчивость и отбор) представляет собой универсальный способ возникновения новой информации.
Благодаря общности системных свойств оказывается, что работы в таких далеких друг от друга областях, как биология, теория лазеров, исследования автоматов, химия, термодинамика оказываются полезными друг для друга. Один из основоположников кибернетики, фон Нойман, занимался теоретическим исследованием технических систем – автоматов (сейчас мы называем их компьютерами). В частности, он решал проблему возможности самовоспроизведения автоматов. Его исследования показали, что способность к самовоспроизведению принципиально зависит от сложности организации системы. На низшем уровне сложности автоматы способны воспроизводить лишь более простые автоматы. Но на определенном критическом уровне сложность становится самоподдерживающейся или даже может расти.
Этот вывод фон Ноймана крайне важен, так как указывает на универсальное свойство открытых систем – для устойчивости системы критической является ее сложность. Такие биологические системы как экосистемы и биосфера в целом также являются сложными, отличающиеся большой устойчивостью, то есть способностью восстанавливать случайные нарушения. Но эта устойчивость зависит от сложности. Уменьшение биоразнообразия на Земле опасно не с точки зрения эмоций – вымер еще один вид цветочков или бабочек, а потому, что уменьшающаяся сложность может в конечном итоге привести к гибели всей системы.
§ 23. Современные представления о происхождении жизни.
Представления о возникновения жизни на Земле. С глубокой древности и до нашего времени было высказано бессчетное количество гипотез о происхождении жизни на Земле. Первая научная гипотеза о происхождении жизни принадлежит известному биохимику А. И. Опарину. В 1924 г. он предположил, что при мощных электрических разрядах в земной атмосфере, которая 4—4,5 млрд лет назад состояла из аммиака, метана, углекислого газа и паров воды, могли возникнуть простейшие органические соединения, необходимые для возникновения жизни. В 1955 г. американский исследователь С. Миллер экспериментально доказал, что это возможно. Пропуская электрические разряды напряжением до 60000 вольт через смесь СН4, NH3, H2 и паров Н2О под давлением в несколько паскалей при температуре 80°С, он получил простейшие жирные кислоты, уксусную и муравьиную кислоты, несколько аминокислот и другие органические соединения.
Однако другое предположение Опарина о том, что предшественниками первых клеток были коацерваты – сгустки белка, которые могут образовываться в растворах, в настоящее время большинством ученых не принимается. Причиной этого является отсутствие способности белков хранить и передавать наследственную информацию.
Невозможность точного восстановления первых этапов биогенеза. Исходя из того, что известно о свойствах сложных систем, об их способности к самоорганизации, можно попытаться представить себе возможный сценарий возникновения жизни, имея в виду, что проверить, как это было на самом деле, невозможно и никогда не будет возможно. Принципиальная невозможность установления «как это было» связана с тем, что система может двигаться по бесконечному количеству траекторий, даже если она в конечном итоге достигнет одного и того же аттрактора. Даже если удастся в лабораторных условиях создать искусственную жизнь абиогенным путем, это не будет доказательством того, что именно так возникала жизнь на Земле. Поэтому можно только представлять себе более или менее вероятные сценарии и по возможности экспериментально проверять, возможны ли в принципы те или иные этапы этого сценария.
Формирование планеты и первичной атмосферы. Примерно 7 миллиардов лет назад сформировалась Земля и другие планеты Солнечной системы. Считается, что первичная атмосфера Земли состояла из паров воды, метана, аммиака, диоксида углерода, водорода с примесью инертных газов (гелия, аргона, криптона, ксенона). При извержениях вулканов выделялись сероводород, фтористый водород, хлористый водород, цианистый водород, угарный газ (монооксид углерода). При подводных извержениях вулканов в результате пиролиза воды мог образовываться газообразный кислород. Легкие газы (водород и гелий) не могли удерживаться Землей, масса которой в то время была значительно меньше, чем сейчас, и улетучивались в космическое пространство, кислород немедленно использовался в реакциях окисления. В результате образовалась вторичная атмосфера Земли, состоявшая из метана, аммиака, углекислого газа и некоторого количества водорода. Кроме кислорода, во вторичной атмосфере отсутствовал азот.
Абиогенный синтез органических веществ и химическая эволюция. Следующий этап – абиогенный синтез органических веществ, достаточно хорошо исследован и воспроизведен в лабораторных условиях. В районах вулканических извержений, сопровождавшихся мощными грозовыми разрядами, могли синтезироваться некоторые аминокислоты, мочевина, формальдегид. Экспериментально в подобных условиях удалось получить и первые нуклеотиды – аденозинфосфат и уридинфосфат.
Далее следует химическая эволюция реакций, реакционных циклов, автокаталитических циклов и, наконец, гиперциклов. Какие это были реакции и циклы? Очень сомнительно, что самые первые циклы включали даже РНК. Дело в том, в результате абиогенного синтеза возникают самые разные органические молекулы (да еще в виде рацемической смеси), которые при этом еще и постоянно вступают в различные реакции. А для возникновения хотя бы самого простого автокаталитического цикла нужно много однотипных молекул, причем в одном месте и в одно время.
«Глиняные гены». Шотландский ученый А.Дж.Кернс-Смит предположил, что первые такие системы могли иметь неорганическую природу, например, это могли быть процессы кристаллизации глин разного состава – этим решается проблема одновременного наличия большого количества одинаковых молекул. Не углубляясь в детали этого сложного процесса, можно только отметить, что «глиняные элементы жизни» обладают необходимыми свойствами – способностью к самовоспроизведению (рис. 45) с присутствием случайных флуктуаций, которые также могут воспроизводиться и подвергаться действию естественного отбора. Вспомним, что область функционирования механизма дарвиновской эволюции значительно шире, чем это представлял себе Дарвин и чем представляют это себе очень многие люди и сейчас. Как уже 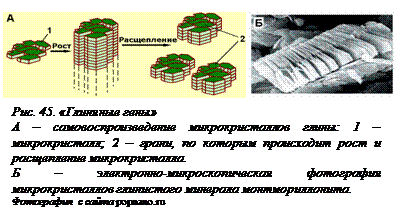 говорилось, это единственный понятный нам способ возникновения новой информации. Для дальнейшего возникновения органической жизни такие «глиняные гены» могли послужить первоначальной матрицей, «подпоркой», на которой возникавшие органические молекулы могли организовываться в новые циклы. Такой «симбиоз» минеральных и органических систем мог быть взаимно выгодным. Известно, что многие из тех органических молекул, которые тогда возникали, обладают определенным сродством к алюмосиликатам (глинам), могут стабилизировать и облегчать их кристаллизацию (элемент катализа), а алюмосиликаты, в свою очередь, являются (это тоже известно) катализаторами полимеризации органических молекул. Впоследствии, когда накопилось достаточное количество и разнообразие органических молекул, способных образовывать гиперциклы без участия глин, они, будучи более эффективными, вытеснили минеральные элементы. Можно это пояснить на образном примере строительства арки из камней. Наклонные камни в арке держатся за счет замкового камня, если его убрать, арка рухнет. Поэтому невозможно построить арку, последовательно складывая камни на камни – она рухнет до того, как мы дойдем до замкового камня. Получается, что арка существовать не может. Но она существует! Почему? Потому, что сначала была построена подпорка, матрица, совсем из другого материала (может быть, из досок или из мешков с песком), на матрице выложена арка до замкового камня, после чего подпорка была разобрана и исчезла бесследно. Роль такой подпорки и могли сыграть «глиняные циклы», в результате чего циклы, построенные из биополимеров, и смогли появиться сразу в достаточно сложном виде.
говорилось, это единственный понятный нам способ возникновения новой информации. Для дальнейшего возникновения органической жизни такие «глиняные гены» могли послужить первоначальной матрицей, «подпоркой», на которой возникавшие органические молекулы могли организовываться в новые циклы. Такой «симбиоз» минеральных и органических систем мог быть взаимно выгодным. Известно, что многие из тех органических молекул, которые тогда возникали, обладают определенным сродством к алюмосиликатам (глинам), могут стабилизировать и облегчать их кристаллизацию (элемент катализа), а алюмосиликаты, в свою очередь, являются (это тоже известно) катализаторами полимеризации органических молекул. Впоследствии, когда накопилось достаточное количество и разнообразие органических молекул, способных образовывать гиперциклы без участия глин, они, будучи более эффективными, вытеснили минеральные элементы. Можно это пояснить на образном примере строительства арки из камней. Наклонные камни в арке держатся за счет замкового камня, если его убрать, арка рухнет. Поэтому невозможно построить арку, последовательно складывая камни на камни – она рухнет до того, как мы дойдем до замкового камня. Получается, что арка существовать не может. Но она существует! Почему? Потому, что сначала была построена подпорка, матрица, совсем из другого материала (может быть, из досок или из мешков с песком), на матрице выложена арка до замкового камня, после чего подпорка была разобрана и исчезла бесследно. Роль такой подпорки и могли сыграть «глиняные циклы», в результате чего циклы, построенные из биополимеров, и смогли появиться сразу в достаточно сложном виде.
 Проблема хиральной чистоты биополимеров. Следует упомянуть еще раз о хиральной чистоте биополимеров. Дело в том, что при абиогенном синтезе органических веществ «правые» и «левые» молекулы возникают в одинаковом количестве, то есть получается рацемическая смесь (рис. 46). Но из рацемической смеси невозможно получить молекулы белка с одинаковой третичной структурой (из-за разного направления связей у зеркальных изомеров). По этой же причине из рацемической смеси нуклеотидов не идет синтез нуклеиновых кислот. Живые организмы умеют извлекать из питательной среды один из зеркальных изомеров, но для этого они должны уже существовать. Для того чтобы хиральная чистота возникла, должно было быть нарушено равновесие, то есть один из зеркальных изомеров должен был хотя бы на одном каком-то этапе возникать в большем количестве, чем другой. Известный физик В.И.Гольданский предположил, что на древней Земле с ее мутной атмосферой, содержавшей вулканическую пыль и газы, могла иметь место неоднородность поляризации солнечного света, что могло привести к нарушению равновесия состава изомеров синтезирующихся органических молекул.
Проблема хиральной чистоты биополимеров. Следует упомянуть еще раз о хиральной чистоте биополимеров. Дело в том, что при абиогенном синтезе органических веществ «правые» и «левые» молекулы возникают в одинаковом количестве, то есть получается рацемическая смесь (рис. 46). Но из рацемической смеси невозможно получить молекулы белка с одинаковой третичной структурой (из-за разного направления связей у зеркальных изомеров). По этой же причине из рацемической смеси нуклеотидов не идет синтез нуклеиновых кислот. Живые организмы умеют извлекать из питательной среды один из зеркальных изомеров, но для этого они должны уже существовать. Для того чтобы хиральная чистота возникла, должно было быть нарушено равновесие, то есть один из зеркальных изомеров должен был хотя бы на одном каком-то этапе возникать в большем количестве, чем другой. Известный физик В.И.Гольданский предположил, что на древней Земле с ее мутной атмосферой, содержавшей вулканическую пыль и газы, могла иметь место неоднородность поляризации солнечного света, что могло привести к нарушению равновесия состава изомеров синтезирующихся органических молекул.
В неживой природе оптическая активность свойственна некоторым кристаллам. Можно предположить, что геометрической форме микрокристаллов «глиняных генов» более точно соответствовал только один из двух зеркальных изомеров органических молекул, что и привело к нарушению равновесия.
«Мир РНК». По-видимому, первым биополимером была РНК. Она обладает принципиально нужными свойствами – способностью к самовоспроизведению с флуктуациями (мутациями), которые запоминаются и подвергаются естественному отбору. Причем с открытием рибозимов – ферментов не белковой, а РНКовой природы, стало ясно, что для самовоспроизведения ей даже не нужны были ферменты, она сама себе была ферментом (автокатализ), способным удлинять цепочки РНК. Другими словами, РНК могла выполнять основные функции живых систем – хранить информацию, самовоспроизводиться. Для этого ей не нужны были никакие другие макромолекулы. Поэтому ученые предполагают, что древнейшая жизнь представляла собой «мир РНК». Конечно, это была немного другая РНК, скорее всего она состояла только из двух нуклеотидов – аденозина и уридина. Это именно те нуклеотиды, которые удалось получить абиогенным путем. В пользу этого свидетельствует тот факт, что аденозинфосфаты выполняют в клетке самые различные функции – и функцию мономера всех нуклеиновых кислот, и универсальной энергетической валюты, и самых распространенных коферментов (НАД·Н, НАДФ·Н, ФАД·Н2, КоА). А все самые главные операции синтеза белков осуществляют РНК – именно они приносят информацию о последовательности аминокислот (мРНК) и сами аминокислоты (тРНК, которые присоединяют аминокислоты именно к своему концевому аденозину), в рибосоме сам акт катализа образования пептидной связи осуществляет рРНК (причем в активном центре главным «действующим лицом» является опять-таки аденозин!). Такими «реликтами РНК-мира», по-видимому, являются и малые РНК, которые, как оказалось, выполняют в клетке самые разнообразные и очень важные регуляторные функции. Сравнение последовательностей аминокислот в ферментах ДНК-зависимой РНК-полимеразе и РНК-зависимой РНК-полимеразе указывает на то, что именно РНК-зависимая РНК-полимераза была самым древним ферментом на Земле из ныне существующих. По-видимому, потом возникла трансляция (скорее всего, она возникала более или менее одновременно с возникновением репликации РНК), и оказалось, что с катализом лучше справляются белки. Поэтому ферментативная функция (как и структурная) была «передана» белкам. Потом функция сохранения генетической информации была передана ДНК. ДНК для этого более пригодна, чем РНК, так как в слабощелочных растворах она стабильна, кроме того, двуцепочечность молекулы повышает надежность хранения информации.
Гетеротрофы или автотрофы? Неясно, были действительно первые организмы, как принято считать сейчас, гетеротрофами. Дело в том, что процессы фото- и хемосинтеза очень сложны, для них нужны специализированные ферментативные системы, для возникновения которых нужна была достаточно долгая эволюция. То есть, сначала должны были существовать гетеротрофные организмы, у которых потом эти системы возникли. Но предположение о первичности гетеротрофов наталкивается на одно принципиальное затруднение – прежде чем в ходе эволюции возникнет автотрофность, запасы органики будут исчерпаны (так как размножение автокаталитических гиперциклов идет экспоненциально). Но пока органики в среде достаточно, нет нужды в ферментативных системах автотрофного питания (весьма и весьма сложных), а, значит, и нет отбора в направлении образования этих систем, другими словами, им незачем возникать. Понятно, что для «глиняных организмов» (для неорганических систем вообще) этот вопрос не имеет смысла. Кажется весьма вероятным, что элементы автотрофности возникали еще до вполне органической стадии (на смешанной «минерально-органической» стадии). Другими словами, автотрофность могла возникнуть до возникновения организмов, полностью построенных из белков и нуклеиновых кислот, то есть еще на доклеточной стадии. Если такая «минеральная» стадия биопоэза действительно имела место, она, скорее всего, была неклеточной, а в мембраны жизнь оделась лишь после «эмансипации» от глины.
Отечественный ученый В.П.Скулачев выдвинул достаточно обоснованную гипотезу о том, что первоначальные процессы улавливания энергии из внешней среды были значительно более простыми, чем сейчас. Он предположил и подтвердил расчетами, что мог возникнуть фотосинтез не на основе хлорофиллов, а на основе АТФ, которая улавливала энергию ультрафиолетового излучения Солнца. По мере того, как атмосфера становилась все более непрозрачной для ультрафиолета, фотосинтез был «передан» пигментам (хлорофиллу, каротиноидам), поглощающим более длинноволновой видимый свет. Однако, несмотря на то, что «АТФ-фотосинтез» значительно более прост, чем хлорофилльный, он все-таки предполагает существование достаточно развитого обмена веществ и наличие мембраны, на которой происходит разделение зарядов и перекачка протонов. Другими словами, он должен был возникнуть на более поздней стадии развития жизни.
Некоторые ученые считают, что жизнь вообще зародилась в тех местах, куда свет не попадает – в глубине океанов в районах выхода гидротермальных вод или в толще земной коры, пропитанной горячими растворами вулканического происхождения. Возможно, что именно хемоавтотрофы и были первыми живыми организмами на Земле.
Самоорганизация на разных уровнях организации жизни. Роль процессов самоорганизации не исчерпывается только возникновением живой материи. Самоорганизация, присущая неравновесным открытым системам, проявляется на разных уровнях организации жизни. В частности, наблюдаемое разнообразие, например, видов в роде, клеток в организме и т. п. также является результатом самоорганизации. Подсчитано, что для системы, состоящей из 25 тыс. элементов (приблизительно столько генов у млекопитающих), которые связаны друг с другом относительно небольшим количеством прямых и обратных связей, может существовать около 200 аттракторов (столько типов клеток в организме млекопитающих). По-видимому, именно поэтому наблюдается дискретность на всех уровнях организации жизни – дискретность клеток, дискретность тканей (типов клеток), дискретность организмов, видов, экосистем. Эти дискретные единицы жизни представляют собой аттракторы такой сложной системы, как живая материя.

