




Половой член, придаточные половые железы и мочеполовой канал питают артериальные ветви внутренней срамной артерии, берущей начало от внутренней подвздошной артерии. Половой член получает также веточки от запи-рательной и от наружной срамной артерий. Срамная артерия участвует в кровоснабжении мошонки и препуция. Семенник, его придаток, спермиопровод и оболочки получают артериальные ветви от проходящих в семенном канатике внутренней семенной артерии, артерии семявы-носящего протока и наружной семенной артерии. Наиболее мощная — внутренняя семенная артерия берет начало от аорты в поясничной области и по пути к паховому каналу проходит длинный извилистый путь. Это наряду со сравнительно небольшим диаметром артерии способствует охлаждению артериальной крови на пути к семеннику. Артерия семявыносящего протока тоньше внутренней семенной артерии и через семенной канатик направляется к спермиопроводу. Наиболее мелкая из трех артерий семенного канатика — наружная семенная артерия, отходящая от наружной подвздошной артерии. Она разветвляется во влагалищной оболочке семенника и семенном канатике. Внутренняя семенная артерия у сельскохозяйственных животных образует особую извилистость в области пахового канала и мошонки. При этом радиус и количество извилин артерии возрастают в направлении сверху вниз, в результате чего образуется тело конической формы, обращенное вершиной вверх, а основанием к семеннику. Извивы артерии находятся в тесном контакте с массивным венозным сплетением внутренней семенной вены, в результате чего создаются благоприятные условия для охлаждения артериальной крови. Ветви внутренней семенной артерии значительной длины формируют обширную поверхностную артериальную сеть из довольно крупных стволиков и петель в белочной оболочке и только после этого погружаются в паренхиму семенника.
Внутренняя семенная артерия делится на семенниковую и придатковую. Придатковая артерия входит в придаток из белочной оболочки. Все артерии семенного канатика связаны между собой анастомозами, поэтому повреждение даже внутренней семенной артерии часто не приводит к значительным нарушениям кровоснабжения семенника. В семеннике более крупные артерии проходят
49
в междольковых перегородках. Большая же часть артерий среднего калибра проникает в дольки и проходит между отдельными канальцами, широко анастомозируя между собой. Артериальное русло семенника имеет структуру сложной сети. Канальцы семенника густо оплетены кровеносными капиллярами, а каждый каналец по своему ходу получает питание от артерий, происходящих из различных источников. В семеннике ни одну из артерий нельзя назвать конечной.
Вены семенника образуют сетевидные сплетения венозного русла семенника. Внутри долек семенника обычно две вены сопровождают одну артерию. Венозная кровь выносится из половых органов в каудальную полую вену по одноименным с артериями венам.
Половые органы иннервируются ветвями поясничного и крестцового сплетений, а также симпатической цепочки. Веточки крестцовых нервов образуют сплетения с веточками симпатических нервов. Веточки этого семенного сплетения иннервируют простату, спермиопровод, а часть их, анастомозируя, проходит в семенники. После кастрации патологически измененные клетки появляются в межпозвоночных узлах, в подчревном и нижнем крестцовом симпатических узлах и спинном мозгу. В спинном мозгу патологические клетки сосредоточивались в 3-м, 4-м и 5-м поясничных сегментах и в 1-м, 2-м крестцовых. После перерезки тазовых нервов в семенниках наблюдается атрофия семенных канальцев, гипертрофия интерстици-альной ткани и значительное ослабление сперматогенеза. В иннервации семенников участвует также блуждающий нерв. Иннервация блуждающим нервом осуществляется как через солнечное сплетение, так и прямыми веточками к семенникам. При удалении спинномозговых чувствительных узлов от Дю до Ь.2 в семенниках отмечали сосудистые расстройства, отек соединительной ткани семенников и придатка, прекращение сперматогенеза, запусте-вание семенных канальцев и разделение полулунных узлов солнечного сплетения приводило к тяжелым изменениям в семенниках. Помимо расстройств, связанных с деафферентацией семенников, при этом происходило разрушение гладкой мускулатуры сосудов, связанное с нарушением их эфферентной симпатической иннервации. После субдиафрагмальной перерезки блуждающего нерва регистрировали уменьшение массы семенников, количества семенных канальцев, содержащих спермин, и снижение гормональной активности семенников. Удаление
50
пограничных симпатических стволов в пояснично-крест-цовом отделе у взрослых животных приводило к уменьшению массы семенников и снижению их гормональной активности. Это было связано как с перерезкой симпатических, так и с нарушением афферентных волокон, проходящих в составе этих стволов.
Основным источником иннервации семенника является внутреннее семенное сплетение. Оно в 27 % случаев формируется ветвями каудального брыжеечного сплетения, в 21 % — ветвями внутреннего семенного узла и в 52 % случаев в его формировании участвуют оба источника. В 15 % случаев в сплетение вступают волокна подчревного нерва. В ткань семенника нервные волокна проникают в основном с сосудами. Они состоят из мякот-ных и безмякотных волокон. От многочисленных сплетений мякотных волокон вокруг сосудов в мышечные стенки сосудов отходят двигательные окончания. В тесном контакте с нервными окончаниями находятся клетки Лейдига. Безмякотные нервные волокна образуют сплетения вокруг семенных канальцев. Нервные волокна этих сплетений проникают через стенки семенных канальцев и контактируют с их эпителием. В семеннике имеется также мощный рецепторный аппарат, центры которого находятся в грудном и поясничном отделах спинного мозга. Часть нервных волокон с сосудами разветвляется в оболочках семенника, образуя обильные сплетения, состоящие из мякотных и безмякотных волокон.
Оболочки семенника обеспечены хорошо развитым рецепторным аппаратом. Как париетальный, так и висцеральный листки оболочек семенника имеют гладкомы-шечные волокна, обильно снабженные безмякотными нервами в основном кабельного типа. При перерезке нервов симпатической и парасимпатической систем наблюдалась избирательная дегенерация отдельных волокон. Источником иннервации белочной оболочки служат внутренний семенной нерв, сплетение спермиопровода и наружный семенной нерв. Нервные окончания белочной оболочки являются в основном механорецепторами.
Спермиопровод иннервируется нервным сплетением, которое снабжается главным образом волокнами подчревного нерва, парасимпатическими нервными волокнами. В узел спермиопровода вступают также ветви тазового сплетения. В большинстве случаев нерв спермиопровода берет начало из каудального сплетения мочевого пузыря и пузырьковидных желез.
51
Мочеполовой канал иннервируется ветвями тазового сплетения и нервными ветвями спермиопровода. В мускулатуру мочеполового канала проникают ветви срамного нерва. Пузырьковидная железа иннервируется волокнами из подчревного сплетения и тазовыми нервами. В предстательную железу вступают волокна простатического сплетения. Симпатические волокна этого сплетения приходят из подчревного, а парасимпатические из 3- и 4-го крестцовых сегментов. В иннервации луковичной железы участвуют тазовое сплетение, срамной нерв и кожный каудальный нерв бедра.
Один из источников иннервации полового члена — тазовое сплетение, образованное подчревными нервами, тазовыми нервами, срамным нервом и ветвями пограничного симпатического ствола поясничной и крестцовой области. Симпатический срамной нерв начинается от 3—4-го крестцового сегмента. Он делится на промежуточный нерв, средний геморроидальный и дорзальный нервы полового члена. Дорзальный нерв полового члена иннервирует пещеристые тела, кончик и препуций. Средний геморроидальный нерв иннервирует вентральную поверхность полового члена. Промежуточный нерв отдает ветвь в ретракторную мышцу. В иннервации полового члена участвуют также соматические подвздошно-под-чревный нерв, подвздошно-паховый и наружный семенной нервы, которые иннервируют кожу боковой, вентральной поверхности полового члена, препуций и мошонку. Ветви тазового сплетения, иннервирующие половой член, состоят из симпатических, парасимпатических и чувствительных нервных волокон. Симпатические волокна происходят из каудального брыжеечного узла, двух каудальных поясничных и всех крестцовых узлов пограничного симпатического ствола. Чувствительные нервные волокна имеют своим источником два последних и все крестцовые спи-нальные ганглии. Парасимпатические волокна являются отростками нейронов, расположенных в узлах тазового сплетения. Таким образом, основные источники иннервации половых органов — парасимпатический и симпатический отделы вегетативной нервной системы. В иннервации участвуют также чувствительные веточки соматических поясничных и крестцовых нервов.
К источникам симпатической иннервации относятся полулунные узлы солнечного сплетения, каудальное брыжеечное сплетение, поясничные и крестцовые узлы пограничного симпатического ствола, подчревный нерв, а к ис-
52
точникам парасимпатической иннервации — блуждающий и тазовый нервы. Парасимпатическая и симпатическая системы отличаются по ряду анатомических и физиологических особенностей. Волокна симпатических нервов берут свое начало от нервных клеток боковых рогов гру-динно-поясничной части спинного мозга. Клетки волокон парасимпатической системы лежат в продолговатом, среднем мозгу и крестцовой части спинного мозга.
Симпатическая и парасимпатическая нервная системы оказывают на органы противоположное действие. Эффект парасимпатической системы обусловлен медиатором аце-тилхолином,.выделяемым окончаниями нервов, а эффект симпатической системы связан с выделением окончаниями нервов адреналина. Изучение физиологической роли отдельных нервов показало, что некоторые нервы, относящиеся по анатомическим признакам к симпатическим, на самом деле являются парасимпатическими. Например, на основании физиологического действия подчревный нерв стали относить к парасимпатическим нервам. Оказалось, что эзерин усиливает, а атропин снимает расширение сосудов, вызываемое раздражением подчревного нерва. Проведенные исследования по влиянию холин-эргнческих и адренэргических веществ на половую функцию показали, что холинэргическая иннервация — ведущая в кинетической деятельности гладкой мышечной ткани семенников, их придатков, спермиопроводов, мочевого канала и придаточных половых желез. При инъекции препаратов, стимулирующих активность парасимпатической нервной системы, удавалось повысить объем эякулята и общее число в нем спермиев. Однако введение атропина хрякам, выключающее влияние парасимпатической иннервации, почти не сказывалось на общем количестве спермиев, выводимых из половых органов в эякулят. Это свидетельствует о более сложном взаимодействии обеих систем, и, возможно, даже при определенных условиях в отдельных случаях симпатические нервные волокна могут перенимать функцию парасимпатических.
ОБРАЗОВАНИЕ ПОЛОВЫХ КЛЕТОК (ГАМЕТОГЕНЕЗ)
В процессе образования зрелых мужских и женских клеток родоначальные половые клетки проходят этапы
53
деления по типу митоза и мейоза. При этом структуры клеток претерпевают существенные изменения.
Строение клеток. Организм животных состоит из клеток, которые произошли в результате соединения двух половых клеток — спермия и яйцеклетки и последующего деления зиготы, образовавшейся при их слиянии. Клетка состоит из оболочки, представляющей собой мембрану. Оболочка отделяет клетку от внешней среды, защищает ее и одновременно обеспечивает связь и обмен веществ между клеткой и окружающей средой. Под оболочкой находится протоплазма, которая разделяется на кариоплазму (вещество ядра) и цитоплазму (остальная часть протоплазмы). В цитоплазме клеток имеются различные структуры, выполняющие специфические функции.
Р и б о с о м ы. Представляют собой мелкие частицы, рассеянные в цитоплазме, иногда в виде скоплений. Они осуществляют синтез белка. Цитоплазматическая (эндо-плазматическая) сеть представляет собой сложную систему трубок, пузырьков и цистерн, стенки которых образованы трехслойной липопротеидной мембраной. Цитоплазматическая сеть является транспортной системой клетки, обеспечивающей связь различных участков внутри клетки и их связь с окружающей средой в процессе синтеза и обмена веществ.
Митохондрии. Имеют форму нитей, палочек или мелких зерен. Осуществляют они клеточное дыхание, т. е. окисление углеводов и жиров, сопряженное с фосфо-рилированием. В них накапливается аденозинтрифосфор-ная кислота (АТФ), являющаяся источником энергии.
Л и з о с о м ы. Представляют собой маленькие мешочки (пузырьки), окруженные мембраной и заполненные жидкостью, содержащей до 30 различных гидролизирующих ферментов. Лизосомы участвуют в пищеварительной деятельности клеток.
Комплекс Гольджи (пластинчатый комплекс). Состоит из цистерн, микропузырьков и вакуолей. В нем происходят синтез веществ, изоляция и выведение из клеток вредных компонентов, попавших в клетку, и продуктов обмена веществ.
Клеточный центр (центросома). Располагается около ядра или ближе к периферии. Основная ее часть — центриоли, мельчайшие частицы, едва различимые в световом микроскопе. Как правило, в клетке имеются две центриоли.
Ядро (кариоплазма) —основной компонент клетки,
54
несущий генетическую информацию. Оно состоит из мембраны, белкового вещества, называемого хроматином, ядрышка и ядерного сока.
Ядерная мембрана отделяет ядро от цитоплазмы. Она представляет собой оболочку, состоящую из двух листков. Через нее осуществляется обмен веществ между кариоплазмой и цитоплазмой. Хроматин состоит из тонких нитей, называемых хромосомами. Именно в них сосредоточена основная генетическая информация, или наследственность. У домашней свиньи в ядре содержится 38 хромосом. В каждой хромосоме по две нити (хроматиды), располагаются они параллельно и соединяются в одной точке, называемой центромерой, или перетяжкой. У некоторых хромосом существуют две перетяжки. Участок хромосомы от конца до перетяжки называется плечом. Хромосомы бывают равноплечими и неравноплечими. Каждая хромосома в соматической клетке самки имеет сходную с ней по строению и размерам другую хромосому. Такие хромосомы называются гомологичными. Одна из них передается в родоначальную клетку организма (зиготу) спермием, а другая — яйцеклеткой. Каждая пара гомологичных хромосом отличается от других пар по строению, размерам и функциям. В организме самца одна пара хромосом не является гомологичной. Одна хромосома из этой пары называется Х (икс)-хромосомой и передается яйцеклеткой, а другая, передаваемая спермием, называется У (игрек)-хромосомой. У самок в этой паре имеются две гомологичные Х-хромосомы. Эти пары хромосом, единственные неодинаковые у самца и самки, называются половыми хромосомами, все остальные — аутосомами.
Количество ядрышек в ядре зависит от типа клетки. В ядрышках осуществляется синтез белков и рибосомной рибонуклеиновой кислоты. Под ядерным соком понимают остальную часть ядра, кажущуюся бесструктурной при использовании светового микроскопа. В нем сосредоточена большая часть ферментов ядра.
Деление клеток. Деление всех клеток в организме происходит по типу митоза (рис. 7). Только отдельные этапы деления половых клеток протекают по типу мейоза. При делении по типу митоза из одной материнской клетки образуются две дочерние, одинаковые с ней по генетическому материалу. В процессе деления клетки по типу митоза выделяют две фазы. Одну из них, заключающуюся в подготовке клетки к делению, называют интерфаз-
55
ной, другую — митозом. Деление занимает по времени только небольшую часть процесса. Интерфазу разделяют на три периода:
пресинтетический, синтетический и постсинтетический. В первом периоде в только что образовавшейся после деления молодой клетке проходят усиленный рост цитоплазмы и подготовка к следующему делению. При этом отмечают удвоение цент-риолей. Во втором периоде осуществляется синтез ДНК и удвоение хромосом. Во время третьего периода в клетке накапливается энергия и образуются крупные молекулы, необходимые для формирования структур, участвующих в делении клетки. В митозе различают четыре фазы: профазу, мета-фазу, анафазу и тело-фазу.
|
|
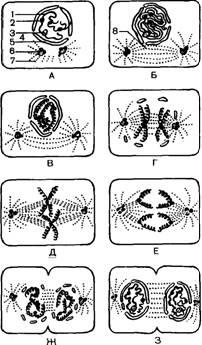
| Рис. 7. Схема митотического деления животной клетки с четырьмя хромосомами (гаплоидное число равно 2): А — интерфаза (стадия покоя); Б — ранняя профаза (центриоль разделилась); В и Г — более поздняя профаза; Д — метафаза; Е и Ж — ранняя и поздняя анафазы; 3 — телофаза (ядерная оболочка изчезла, началось деление цитоплазмы); / — Х-хромосома; 2 — центромера; 3 — ядрышко; 4 — ядро; 5 — ядерная оболочка; 6 — центриоль; 7 — звезда; 8 — веретено |
В профазе хромосомы вначале представляют собой клубок длинных тонких нитей. К концу фазы длина их резко уменьшается за счет спирализации. Ядрышко разрушается и
его содержимое участвует в образовании веретена деления. Центриоли разделяются и расходятся к противоположным полюсам клетки. Удлиняются прикрепленные к ним нити веретена. Ядерная оболочка распадается.
В метафазе утолщенные спиралеобразные хромосомы перемещаются в экваториальную плоскость клетки. К каждой из них с двух сторон прикрепляются нити полуверетена.
56
В анафазе клетка и веретено удлиняются, удвоенные хромосомы разделяются на хроматиды, которые затем движутся к противоположным полюсам.
Во время телофазы хромосомы расходятся к полюсам, в середине клетки образуется перетяжка, веретено исчезает и две дочерние клетки полностью отделяются друг от друга. Ввиду того, что ядерное вещество и цитоплазма материнской клетки в процессе митоза разделяются на две равные части, каждая дочерняя клетка полностью соответствует материнской на соответствующей стадии.
Мейотическое деление происходит в процессе образования половых клеток. При этом набор хромосом в сравнении с материнской клеткой уменьшается в 2 раза и из диплоидного становится гаплоидным, а хромосомы изменяют свой состав, так как каждая хромосома обменивается своими участками с гомологической хромосомой и в результате имеет часть генов отца и часть матери.
В процессе м е и о з а выделяются две стадии деления:
редукционное и уравнительное. В профазе редукционного деления выделяют пять стадий: лептонемы, зигонемы, пахинемы, диплонемы и диакинеза. В стадии лептонемы хромосомы деспирализованы. Они располагаются в ядре в виде тонких нитей и состоят из двух хроматид. В стадии зигонемы ядро увеличивается в размерах, гомологичные хромосомы притягиваются друг к Другу и сливаются. На стадии пахинемы хромосомы укорачиваются и утолщаются. В увеличенных по объему ядрах появляются ядрышки. В стадии диплонемы соединившиеся хромосомы начинают отталкиваться. Их расхождение от центромеры к концам возрастает. В точках соприкосновения хромосом происходят разрывы и обмен участками между гомологичными хромосомами. Это явление получило название кроссинговера. В диакинезе отмечают укорачивание хромосом, исчезает ядерная мембрана. В метафазе 1 тетрады располагаются в области экваториальной пластинки.
В анафазе 1 начинается расхождение гомологичных хромосом к противоположным полюсам. Распределение гомологичных хромосом в дочерние клетки носит случайный характер. В телофазс 1 хромосомы достигают полюсов клетки, восстанавливаются ядерные оболочки и ядрышки, происходит деление цитоплазмы и образуются две дочерние клетки с гаплоидным набором хромосом.
Отличительной особенностью мейоза от митоза является то, что при этом к каждому полюсу клетки отходит одна гомологичная хромосома, состоящая из двух хро-
57
матид. При митозе каждая хромосома делится на две хроматиды и к полюсам отходит по одной хроматиде из каждой хромосомы. После окончания первой стадии деления клетки существуют в непродолжительном периоде покоя, а затем переходят ко второму делению по типу митоза. При этом каждая хромосома разделяется на две хроматиды, одна из которых отходит в одну дочернюю клетку, а другая — во вторую. В результате двух мейоти-ческих делений из одной клетки образуются четыре с гап-лоидным набором хромосом, каждая из которых состоит из одной хроматиды.
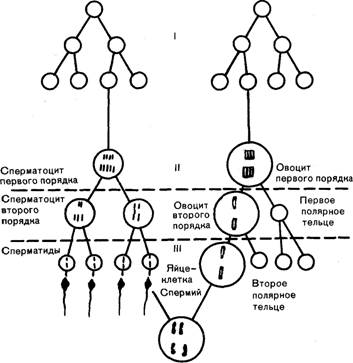
Сперматогенез. Сперматогенезом называется процесс образования спермиев из зародышевых клеток семенников (рис. 8). Родоначальниками зрелых половых клеток являются г о н о ц и т ы, поступающие из желточного
Сперматогенез Овогенез
Зигота
Рис. 8. Схема сперматогенеза и овогенеза у животных с гаплоидньм числом хромосом, равным 2
58
мешка в половую складку зародыша еще в ранний эмбриональный период. Отдельные авторы называют их также стволовыми клетками, или протогониями. Морфологически они не отличимы от ранних сперматогоний. В семенных канальцах семенника гоноциты лежат изолированно и делятся нерегулярно. Они не принимают участия в сперматогенезе. По неизвестным причинам отдельные гоноциты начинают делиться и вступают в сперматогенез. При этом клеточные деления полностью не завершаются и образующиеся клетки остаются соединенными цитоплазмати-ческими мостиками. Сперматогенез начинается у хрячков в возрасте 80—90 дней. После нескольких нерегулярных делений, при которых образуется клон соединенных друг с другом клеток, деления становятся регулярными. В процессе каждого деления клетки все более дифференцируются и достигают стадии поздних сперматогоний, которые называют сперматогониями типа В. Все менее дифференцированные клетки, начиная от стволовых, называются сперматогониями типа А. Для каждого вида животных число клеточных делений сперматогоний — постоянная величина и колеблется от 4 до 6. С каждым делением сперматогоний уменьшаются в размерах.
Сперматогоний типа В вступают в последнее деление, при котором образуются сперматоциты первого порядка. На этом завершаются первый этап сперматогенеза, размножение сперматогоний и начинается второй этап — мейоз. Сперматоциты первого порядка вступают в мейотическое деление, при котором образуется два спсрматоцита второго порядка с гаплоидным (одинарным) набором хромосом. Ввиду того, что у сперматогоний и спер-матоцитов первого порядка половая пара хромосом содержит Х- и У-хромосому, при делении сперматоцита первого порядка в одну из дочерних клеток переходит У-, а в Другую — Х-хромосома. После непродолжительного периода покоя сперматоциты второго порядка вступают во второе мейотическое деление, которое протекает по типу митоза. В результате этого деления получаются две спермати-д ы; при делении сперматоцита с Х-хромосомой — две сперматиды с У-хромосомой и т. д. Таким образом, половина образующихся в дальнейшем из сперматид спермиев имеет У-хромосому и называется мужской, так как после их слияния с яйцеклеткой образуется мужской организм, а другая половина — Х-хромосому и называется женской, так как из оплодотворенной ими яйцеклетки развивается самка.
Сперматиды появляются у хрячков в возрасте 4 месяцев. На этом заканчивается второй этап сперматогенеза и начинается третий — спермиогенез, при котором сперматиды превращаются в зрелые половые клетки — спермии. Они регистрируются в семенниках хрячков с 4-месячного возраста. В процессе спермиогенеза происходят сложные преобразования, затрагивающие все клеточные структуры. Из гранул комплекса Гольджи формируется акросома. Она представляет собой мембранную структуру, тесно прилегающую к переднему участку ядра спермия и покрывающую спереди его головку. Акросома (чехлик) содержит большое количество углеводов, ферменты гиалуронидазу, кислую фосфатазу и протеоли-тический фермент акрозин, который наряду с гиалуро-нидазой способствует проникновению спермия через оболочку яйцеклетки.
По преобразованиям в акросомной системе спермиогенез делят на этапы. Количество их у разных животных составляет 15—19. Во время ранних этапов начинается формирование акросомной системы на переднем полюсе ядра. Центриоли мигрируют к противоположному полюсу сперматиды. Дистальная центриоль дает начало жгутику, который растет в узкое щелевидное пространство между сперматидой и цитоплазмой клетки Сертоли. Ядро сперматиды удлиняется, хроматин в нем конденсируется, становится морфологически бесструктурным, а метаболически инертным. Из него образуется головка спермия. Цитоплазма перемещается к заднему полюсу ядра и окружает жгутик. Значительная часть ее сбрасывается. Митохондрии выстраиваются в спираль, она обвивает хвостик спермия в его средней части. В образовавшихся спермиях разрушаются связывающие их цитоплазматические мостики. Отростки клеток Сертоли освобождают их, и они попадают в просвет семенного канальца.
Вступившие в сперматогенез гоноциты еще не успевают достигнуть полного развития, когда в этом же участке канальца вступают в сперматогенез новые зародышевые клетки.
На поперечном срезе семенного канальца обычно видны четыре-пять генераций клеток сперматогенного эпителия, расположенных слоями и находящихся на разной стадии развития. Ближе к оболочке семенного канальца лежат клетки наиболее молодой генерации, а затем по направлению к просвету возраст каждой последующей генерации увеличивается. С течением времени, ввиду развития спер-
60
матогенных клеток, картина клеточных ассоциации в каждом участке семенного канальца меняется.
Процесс, при котором наблюдавшаяся в определенный момент ассоциация клеток в участке семенного канальца вновь повторяется, называется циклом спермато-генного эпителия. Часть цикла, характеризующаяся определенным сочетанием клеток, называется стадией цикла сперматогенного эпителия. Ранее в сперматогенезе выделяли еще один период роста. Считалось, что конечные сперматогонии типа В вступают в рост и при этом превращаются в сперматоциты первого порядка. Однако новейшие исследования показали, что, во-первых, сперматоциты лишь незначительно превышают размеры конечных сперматогонии; во-вторых, сперматоциты образуются не в результате роста, а в результате деления конечных сперматогонии, а рост их происходит на начальных стадиях мейоза.
Большое практическое значение имеет определение длительности сперматогенеза и времени продвижения образовавшихся спермиев через придатки семенников, так как это позволяет установить срок, когда те или иные факторы внешней среды скажутся на качестве эякулятов, выяснить причину ухудшения качества спермы, определить примерную длительность этого периода и сделать правильное заключение о возможности дальнейшего использования хряков в племенных целях. Ни один из использовавшихся ранее методов не позволял точно установить продолжительность указанных процессов. В результате считалось, что период, необходимый на образование спермиев, и поступление их в эякулят, равен 'примерно 1 месяцу и этот процесс можно ускорить соответствующим кормлением или другими приемами. В последнее время применение методов изотопов и Х-радиации позволило установить истинную продолжительность как сперматогенеза в целом, так и отдельных его стадий. Методы были разработаны на основании опытов, которые показали, что меченые атомы См и Рз2 при введении животным включаются в синтезируемую половыми клетками ДНК (от сперматогонии до сперматоцитов первого порядка в лептотене) и передаются в дальнейшем образующимся из них клеткам. По времени появления изотопов в клетках различных стадий сперматогенного цикла удалось установить продолжительность отдельных фаз сперматогенеза. Метод Х-радиации основан на том, что клетки отдельных стадий сперматогенеза имеют различную чувствительность к лу-
61
чевому воздействию. Определенными дозами излучения убивались сперматогонии — клетки наиболее чувствительные к ионизирующему излучению, а развитие оставшихся стадий прослеживалось. В некоторых опытах комбинировались оба метода.
Несмотря на то, что указанные приемы позволяют с высокой точностью определить продолжительность отдельных стадий, в оценке длительности сперматогенеза в целом имелись и существенные расхождения. Основная трудность при этом заключалась в определении длительности периода размножения сперматогонии, так как очень трудно установить время начала деления стволовых клеток, вступающих в сперматогенез. У хряков этот период ориентировочно оценивается в 7,6—8,6 дня. По данным Е. Е. Сви-ерстра (1968), длительность цикла сперматогенного эпителия у хряка составила 8,6+0,1 дня. Продолжительность 1—8 стадий цикла равнялась соответственно 0,9; 1,2; 0,3;
1,0; 0,8; 1,7; 1,6 и 1,0 дня. Длительность стадий сперма-тоцитов первого порядка составила 12,3 дня, сперматоци-тов второго порядка — 0,4, сперматид — 14 дней, в том числе с круглым ядром — 6,3, с удлиненным ядром — 1,5 и спермиев — 6,2 дня.
Длительность сперматогенеза в целом равна 35 дням. Таким образом, период сперматогенеза у хряка оказался значительно короче, чем у быка и барана (около 50 дней), но существенно больше, чем это считалось ранее. Исследования показали, что продолжительность сперматогенеза — постоянная величина. На нее не оказывают влияния гонадотропные гормоны, кормление и другие факторы внутренней и внешней среды. Это дает основание полагать, что внутри половых клеток существует какой-то точный механизм, регулирующий скорость процессов их преобразования. Синхронность процессов сперматогенеза, объясняемая связью группы клеток путем цитоплазматических мостиков, не подтверждается тем фактом, что синхронность процессов обеспечивается и в соседних, не связанных друг с другом синаптическими мостиками клонах клеток.
Устойчивость продолжительности сперматогенеза противоречит данным многих исследований, показывающих, что спермопродукцию у производителей можно сравнительно быстро повысить за счет улучшения их кормления, содержания, изменения режима использования и т. д. Так, на образование спермиев из сперматогонии необходимо около 35 дней, а на их продвижение через придатки
62
еще около 10 дней. Однако изменение условий кормления сказывается на увеличении спермопродукции производителей уже через 17—40 дней. Еще более быстрая ответная реакция наблюдается у производителей при введении им видонеспецифической сыворотки, окситоцина, парасимпа-тикотропных препаратов, гонадотропных гормонов. В некоторых случаях после применения препаратов спермо-продукция возрастала сразу, и в течение длительного времени от подопытных хряков получали на 55 % и более спермиев, чем в контроле. Эти исследования указывают на то, что в семенниках существует механизм, позволяющий быстро повысить уровень выделяемых в эякулят спермиев. С другой стороны, известно, что в случае снижения режима полового использования хряков уровень получаемых от них спермиев резко снижается. Следовательно, существует и механизм быстрого снижения количества спермиев, выводимых в эякулят, в случае уменьшения потребности в них. Как увеличение уровня спермопродукции хряков, так и ее снижение нельзя объяснить за счет процессов, происходящих в придатках семенников и связанных с изменением запаса в них спермиев. В опытах с применением хрякам окситоцина, прозерина, СЖК уменьшение спермопродукции после использования подвижного запаса спермиев из придатков семенников должно было наступать уже после двух-трех получений спермы, так как подвижный запас спермиев оценивается в 60— 70 млрд., а общий их запас в придатках семенников у хряков составляет 175—200 млрд. Однако нами в течение 2-месячного опытного периода получено стойкое увеличение спермопродукции у опытных хряков, которое в некоторых группах (введение СЖК) продолжалось и на протяжении 2-месячного послеопытного периода.
С позиций постоянства сперматогенеза не находят также объяснения факты, наблюдающиеся при длительном половом покое производителей. Если учесть, что в эякуляты хряков поступают спермии, образовавшиеся из сперматогоний, вступивших в сперматогенез за 45 дней до этого, то в случае отсутствия полового использования в придатки семенников будут ежедневно поступать спермин от уже прошедших процессов деления родоначальных сперматогоний. В таком случае при половом покое хряков в придатках должно дополнительно накапливаться около 700 млрд. спермиев, так как уровень суточного спермо-образования у взрослых хряков составляет 15—16 млрд. Однако даже после 6-месячного полового покоя у хряков
63
находят в придатках только 300 млрд. спермиев (обычный запас 175—200 млрд.). Указанное противоречие между постоянством продолжительности сперматогенеза и мобильностью подовых органов в выделении спермиев в большем или меньшем количестве объясняется процессами распада половых клеток на всех этапах сперматогенеза. Исходя из указанных теоретических предпосылок становится понятной возможность быстрого увеличения спер-мопродукции производителей. Она объясняется уменьшением распада половых клеток в семенниках и придатках семенников. Увеличение спермопродукции производителей через значительный промежуток времени после применения самцам стимулирующих приемов связано с усилением процессов деления родоначальных сперматогоний и снижением процессов дегенерации сперматогоний в течение дальнейших циклов их деления. При слабом половом использовании, особенно при длительном половом покое производителей, процессы дегенерации половых клеток усиливаются. В результате, с одной стороны, в придатки поступает значительно меньшее количество спермиев, а с другой — ив самих придатках усиливаются процессы их распада. Указанные явления особенно выражены в случае закупорки спермиопровода или его перевязки.
В зависимости от количества поступающих в придаток молодых спермиев и числа дегенерирующих старых запасы спермиев в придатках могут увеличиваться или уменьшаться. Дегенерацией старых спермиев в придатках и поступлением в них молодых объясняется и тот факт, что даже после длительного полового покоя производителей в первых эякулятах не наблюдается некроспермии, но спермии сохраняют подвижность в придатках около 40 дней. У животных с выраженным сезонным периодом полового покоя в первых эякулятах возможны некроспер- мия, олигоспермия и азаспермия. Однако это связано с тем, что у этих животных в указанный период процессы сперматогенеза затухают, и поступление спермиев в придатки прекращается.
Процесс дегенерации клеток сперматогенного эпителия — рациональное решение. С одной стороны, создаются значительные резервы в размножении животных, с Другой — в случае отсутствия условий для их реализации — энергия и питательные вещества остаются в организме и могут использоваться в нужном направлении.
Овогенез. Овогенезом называется процесс образования женских половых клеток—яйцеклеток (см. рис. 8).
64
Овогонии появляются в яичниках зародышей в возрасте около 45 дней. В 45 дней уже около 20 % из них находится в стадии митоза. Овогонии после ряда делений без перехода в интерфазу вступают впервое мейоти-ческое деление. Одновременно начинается их рост. В этот период овоциты очень бедны цитоплазмой, что связано с предшествовавшими делениями и отсутствием интерфазы на последней стадии. В период роста, связанного с увеличением объема цитоплазмы, отмечается активный синтез РНК. У 50-дневного эмбриона в яичниках появляются овоциты, у 100-дневного эмбриона их количество в 1 мм2 достигает 200, а при рождении — 270. С 90-дневного возраста в яичниках плодов образуются первичные фолликулы. У новорожденных в яичниках встречается небольшое количество делящихся овогоний, но в основном — овоциты, находящиеся в профазе первого мейотического деления. Иногда же у новорожденных свинок встречаются вторичные фолликулы. Во вторичных фолликулах овоциты находятся на диплотено-вой стадии деления. У поросят месячного возраста, все овоциты достигают этой стадии первого мейотического деления. В растущих фолликулах отмечается второй период роста овоцитов. При этом они значительно увеличиваются в размерах.
Указанный период роста характеризуется значительным накоплением в овоцитах белков, желтка и других соединений. Если во время первого периода роста синтез веществ происходит за счет функциональной деятельности самой половой клетки, то во втором периоде роста в процессе уже участвуют окружающие овоцит фолликулярные клетки. Овоциты по достижении диплотеновой стадии деления прекращают дальнейшее развитие, несмотря на рост и развитие фолликула. Последующие процессы деления овоцита восстанавливаются только в период созревания фолликула, т. е. совпадают со временем полового созревания животных. Инициатором продолжения деления яйцеклетки считается преовуляторное выделение ЛГ из гипофиза. При этом происходит быстрое дальнейшее преобразование овоцитов. Через 18—19 ч от начала охоты яйцеклетки в таких фолликулах находятся в метафазе первого деления созревания, а часть из них достигает анафазы. Перед овуляцией фолликулов в яйцеклетках видно полярное тельце и метафаза второго деления созревания.
Второе деление созревания завершается после
3-232 65
проникновения спермия в цитоплазму яйцеклетки, т. е. уже в яйцепроводе. В отличие от других видов животных первое направленное тельце при этом не делится, и в зрелых яйцеклетках свиней содержится не три, а два направительных тельца.
Особенность овогенеза в отличие от сперматогенеза — образование только по одной полноценной клетке после деления овоцита первого и второго порядка, так как в процессе мейоза после деления ядра на две части протоплазма не делится. Вся она остается только с одним из двух образовавшихся ядер. Второе ядро перемещается на периферию клетки и не участвует в дальнейших процессах деления или оплодотворения. Из-за периферического положения его называют полярным тельцем, а из-за его неполноценности — абортивным тельцем. Таким образом, из одного овоцита первого порядка образуется не четыре, как при сперматогенезе, а только одна зрелая половая клетка. Деление ядер, приводящее к образованию двух полярных телец, вероятно, необходимо только для уменьшения в 2 раза набора хромосом в яйцеклетке и осуществления процесса кроссинговера. Хотя у новорожденных поросят отмечают деление небольшого количества овогоний, считается, что новые овоциты при жизни животных не образуются. Весь запас половых клеток, которые достигают созревания за весь период жизни, содержится в организме уже при его рождении. Из большого количества яйцеклеток, имеющихся в яичниках свинок при рождении и оцениваемого в сотни тысяч, созревает только незначительная ее часть. Если считать, что в среднем у свиноматки в течение года проявляется около четырех половых циклов, в период одного цикла развивается около 18 фолликулов, а продолжительность жизни свиноматки составляет в среднем около трех лет, то у нее созревает в течение жизни около 200 яйцеклеток, т. е. минимальная часть из общего запаса. Значительное количество яйцеклеток погибает при жизни животного в атретичных фолликулах, но подавляющая их часть остается в яичниках и служит неиспользуемым резервом.

