




Яйцеклетка свиней имеет сходное строение с яйцами других млекопитающих. Снаружи она покрыта вторичной, прозрачной оболочкой, которая образуется в основном
66
за счет деятельности окружающих ее фолликулярных клеток. Оболочка сложного состава, включающая гиалу-роновую кислоту и муцин. Под ней находится тонкая первичная оболочка, которая представляет собой наружный, уплотненный слой овоплазмы яйца. Она называется желточной, или вителлиновой. Выделившаяся из фолликула яйцеклетка имеет одно полярное тельце и находится в метафазе второго деления созревания (рис. 9). Хромосомы компактные и укороченные.
Пузырчатое ядро с плотным крупным ядрышком, после выделения яйцеклетки из фолликула оно становится неоформленным.
В овоплазме, в отличие от яйцеклеток других видов животных, содержится большое число округлых включений, которые представляют собой желточные зерна диаметром 9,6 мкм. В поверхностном слое находится много шариков различных размеров липоидной природы. Яйцеклетка окружена лучистым венцом из фолликулярных клеток. После овуляции вителлиновая оболочка отслаивается от блестящей и между ними образуется прозрачная полость шириной около 9 мкм. В ней располагается полярное тельце.
|
|
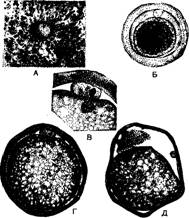
Яйцеклетки — обычно правильной шарообразной формы, встречаются также овальной, полушаровидной и др. Их количество достигает 7 %. Размеры яйцеклеток значительно колеблются как у разных животных, так и у одной особи в период одного цикла. Так, у одной свиноматки крупной белой породы были найдены яйцеклетки, объем которых (без прозрачной оболочки) колебался от 0,31 до 0,95 млн. мкм3.
| Рис. 9. Созревание яйцеклетки свиньи: А — яйцеклетка в сгустке фолликулярных клеток (X 80); Б — неоплодотворенная яйцеклетка из яйцепровода с одним полярным тыьцсм (X 400); В - участок г (X 1350); Г — яйцеклетка в стадии метафазы 2-го деления созревания (X 600); Д— изменения в неоплодотворенной яйцеклетке, вымытой из рога матки через 53 ч после случки свияо-матки с хряком (X 600) |
В среднем диаметр яйцеклеток составляет 165 мкм, а объем — 2,35 млн. мкм3. Толщина прозрач
оболочки равняется мкм. Максимальный
|
Ной 14,8 |
| 3м |
| 67 |
диаметр яйцеклеток составляет 188,8 мкм, минимальный — 140,4 мкм. Максимальный объем — 3,624 млн. мкм3, минимальный — 1,514 млн. мкм3. Овоплазма большей части яйцеклеток в основном также шаровидная или с некоторыми отклонениями от этой формы. Эти отклонения значительно меньше выражены, чем у яйцеклеток. Основной причиной отклонений служит то, что на конфигурацию яйцеклетки оказывает влияние наличие на ее полюсе полярного тельца. При дальнейшем развитии отклонения от шаровидной формы встречаются чаще и объясняются неравномерным расположением в ней бластомеров.
Выделившаяся из фолликула яйцеклетка окружена многочисленными фолликулярными клетками, представляющими собой удлиненный тяж. Все клетки его соединены вязким студнеобразным веществом, в состав которого входят содержащие серу мукополисахариды. Иногда в таком тяже находится несколько яйцеклеток.
Яйцеклетки быстро проходят верхнюю треть яйцепро-вода и попадают в ее среднюю часть (ампулу). Здесь они густо располагаются на расстоянии около 8 см. Иногда уже вскоре после овуляции яйцеклетки находят в месте перехода ампульной части яйцепровода в истмус. Такое быстрое продвижение яйцеклеток опровергает ранее существовавшее представление об их продвижении за счет реснитчатых клеток. В настоящее время установлено, что быстрое продвижение яйцеклеток по яйцепроводу к его истмической части осуществляется за счет своеобразных сокращений его мышечных стенок, создающих вакуум. Эти данные подтверждает также тот факт, что в слизистой передней части яйцепровода отсутствуют реснички.
В яицепроводе происходит быстрое (в течение 1—3 ч) отделение фолликулярных клеток, в том числе и клеток лучистого венца, от яйцеклетки. Наличие спермиев ускоряет этот процесс. Если спермин попадают в яйцепровод раньше яйцеклетки, то они внедряются в ее прозрачную оболочку, когда яйцеклетка еще не полностью отделена от окружающих ее фолликулярных клеток. Считается, что проникновение спермиев через оболочку происходит за счет разрушения входящей в ее структуру гиалуроновой кислоты выделяемым акрозомой спермиев ферментом гиалуронидазой. Вероятно, в этом процессе участвуют и протеолитические ферменты. Проникновение происходит под острым углом. Хвостик спермия совершает при этом интенсивные движения. Период проникновения спермия
68
через оболочку яйцеклетки составляет 3—4 ч. К этому времени в яйцеводах оказываются оплодотворенными до 24 % яйцеклеток. Через 5 ч после осеменения количество оплодотворенных яйцеклеток достигает 72 %, а через 8 ч — 100 %. Попав в овоплазму, спермии поворачивается на 180° и его передняя часть головки направляется в сторону оболочек, а задняя — к ядру яйцеклетки, занимающему периферическое положение. После внедрения спер-мия в овоплазму начинается второе деление созревания. До окончания созревания яйцеклетки в спермии не проходит заметных изменений. В результате деления ядра яйцеклетки образуется второе направительное тельце, оно несколько меньше первого. Диаметр первого направительного тельца достигает 22 мкм, в то время как у второго — 19 мкм. Оба тельца располагаются в перивителлиновом пространстве. После созревания яйцеклетки начинаются изменения в спермии. Его ядро сильно набухает и превращается в мужской пронуклеус. В нем виден хвост спермин. Сходные явления происходят и с ядром яйцеклетки, которое превращается в женский пронуклеус. По размерам он несколько меньше мужского. Структура обоих пронуклеусов незначительно различается. После образования пронуклеусы передвигаются к центру, сближаясь друг с другом. При этом они продолжают набухать и их размеры увеличиваются в 2—3 раза в сравнении с первоначальными. Если образование пронуклеусов происходит довольно быстро и уже через 6 ч после осеменения их находят в яйцеклетке, то период от их образования до слияния оказывается более продолжительным (12—20 ч). После слияния пронуклеусы уменьшаются в размерах, у них исчезают ядрышки и ядерные оболочки, гомологичные хромосомы движутся навстречу друг к другу. На этом процесс оплодотворения яйцеклетки заканчивается и образуется новая клетка — зигота, в ней ядро имеет диплоидный набор хромосом.
В яйцепровод после осеменения попадает большое количество спермиев. Масса их внедряется в прозрачную оболочку яйцеклетки. Известно, что в случае проникновения в яйцеклетку нескольких спермиев наступает так называемая полиспермия, что приводит к нарушению развития и гибели зиготы. Поэтому в яйцеклетках многих видов животных существует механизм, предотвращающий полиспермию. После проникновения первого спермин через прозрачную оболочку в ней быстро происходят тонкие биохимические изменения, в результате чего она становит-
69
ся труднопроницаемой для других спермиев. Однако у свиней процесс полиспермии в последнее время считается нормальным явлением. Через прозрачную оболочку обычно проникает несколько спермиев. Некоторые из них попадают в перивителлиновое (околожелточное) пространство, один или несколько спермиев проходят в ово-плазму. При этом может образоваться несколько мужских пронуклеусов. Один из них увеличивается в объеме, перемещается в центр и сливается с женским пронуклеусом. Остальные постепенно исчезают в протоплазме яйца в период дробления зиготы.
Исследования свидетельствуют о том, что для процесса оплодотворения яйца необходимо большое количество спермиев. Установлено, что чем больше спермиев проникает в прозрачную оболочку, тем быстрее в дальнейшем происходит дробление зиготы. Однако истинная роль большого числа спермиев, участвующих в оплодотворении яйцеклетки, остается недостаточно ясной. Одним из объяснений важности этого процесса могло служить явление избирательности оплодотворения, высказывавшееся многими исследователями. При этом считалось, что яйцеклетка пропускает через оболочку только определенного спермия, который по своей генетической структуре способствует образованию наиболее жизнеспособной зиготы. Мнение об избирательности оплодотворения подтверждалось тем, что в этом процессе, как правило, участвуют только гаметы, принадлежащие одному виду животных. При смешивании спермы самцов, один из которых принадлежал к той же породе, что и самка, а второй — к другой обычно больше потомков получали от последнего производителя. Это же наблюдали и при двойном спаривании самки с самцами. Однако точные исследования показали, что большее количество потомков в этом случае связано не с избирательной способностью яйцеклетки, а с более высокой выживаемостью и меньшей смертностью гетерозиготных эмбрионов. В опытах установлено, что сдвиг количества потомков в сторону одного из производителей связан не столько с разнокачественностью гамет, сколько с количеством спермиев от того или иного самца, поступивших в половые органы самки, и их оплодотворяющей способностью. Высказывается также мнение, что лишние спермии участвуют в передаче дополнительной генетической информации яйцеклетке, ее питании на ранних стадиях развития.
Образовавшаяся зигота имеет размеры 140—190 мкм
70
и шарообразную форму. Она состоит из ядра, протоплазмы, вителлиновой и блестящей оболочек. Двухслойная пористая оболочка покрывает ядро, содержащее коллоидную ядерную лимфу, линин-хроматиновые структуры, половой хроматин в форме хромосом и множество вакуолей. Ядро находится в тесной связи с протоплазмой, в которой имеются многочисленные видимые в оптический или
ЭЛеКТрОННыЙ МИКрОСКОЛ структуры В виде митохондрия,
микросом, центриолей, рибосом и т. д. Желток сосредоточен в виде гранул. Образовавшаяся зигота начинает быстро делиться. Через 20—24 ч некоторые яйцеклетки находятся в стадии 2—4 бластомеров,ав течение 2 суток продвижения по яйцепроводам оказываются на стадии 2—8 клеток (бластомеров). Ввиду того, что размеры зародышей при дроблении не изменяются, величина бластомеров с каждым делением уменьшается. Вначале обычно дробление идет синхронно и образующиеся бластомеры имеют одинаковые размеры. В то же время уже со стадии двух бластомеров может происходить асинхронное деление зародыша, в результате чего можно наблюдать зиготы с нечетным числом бластомеров (3, 5, 7 и т. д.). Число таких зародышей на стадии 2—4 бластомеров составляет около 8 %, на стадии 4—6 бластомеров — 23 %, а на стадии 6—8 бластомеров — превышает 70 %. Это указывает на то, что уже в самом начале дробления зародыша происходит дифференциация клеток.
Слишком значительные отклонения в форме, размерах бластомеров и скорости дробления зародышей указывают на их биологическую неполноценность. В продвижении яйцеклетки по яйцепроводу решающее значение придается сокращению его стенок. В истмической его части мышечные пучки проникают даже в складки слизистой. Реснитчатый эпителий, вероятно, не участвует в этом процессе, так как после овуляции в ампуле яйцевода остаются единичные реснитчатые клетки, которые быстро заменяются секреторными, а в истмической части реснитчатые клетки отсутствуют. Определенную роль в этом процессе играет и секреторная активность слизистой. Перед овуляцией в ампулярной части яйцепроводов резко усиливается секреторный процесс, в результате чего поверхность эпителия покрывается сплошной массой капелек. По мере продвижения яйцеклеток секреция распространяется в направлении истмуса. Секрет имеет определенное значение и в процессе оплодотворения яйцеклеток. Опыты с мечеными атомами указывают на то, что секрет яйцепровода
71
не играет роли в питании зародыша.
Для нормального продвижения яйцеклеток по яйцепро-воду необходим определенный гормональный статус. Его нарушение может вызвать полную задержку зиготы в яйцепроводе и привести к внематочной (трубной) беременности или нарушить распределение зародышей в матке. Установлено, что эстрогены способствуют более быстрому прохождению зародышей, а прогестерон, наоборот, замедляет этот процесс.
На 3-й сутки зиготы попадают в матку, имея 4—8 блас-томеров, округлую форму, а в хорошо выраженной прозрачной оболочке много разрушающихся спермиев.
На 4-е сутки бластомеры в зародыше становятся неразличимыми из-за их многочисленности, малого размера и нечеткости границ между ними. Эта стадия развития называется м о р у л о и. На разрезе зародыша хорошо видна клеточная масса (эмбриобласт), представляющая собой эмбрион. Он покрыт одним слоем более светлых клеток, образующих трофобласт, которые в дальнейшем не участвуют в развитии зародыша. Их функция заключается в обеспечении его питанием. До этого развитие зародыша происходит за счет питательных веществ яйцеклетки и внедрившихся в нее спермиев. После образования тро-фобласта зародыш начинает использовать питательные вещества из окружающих его секретов матки (маточное молочко). В результате накапливающихся в зародыше продуктов обмена веществ в нем появляется полость в центре.
На 5-е сутки зародыш приобретает вид пузырька (стадия бластулы). В результате поступления в зародыш веществ из полости матки внутреннее давление в нем возрастает, он увеличивается в размерах, а его прозрачная оболочка растягивается и истончается.
На 6-е сутки происходит ее разрыв и зародыш освобождается от оболочки. При этом он резко увеличивается в размерах. Его диаметр возрастает с 0,16 до 0,8 мм. Уже в этот период отмечаются существенные различия в величине отдельных зародышей.
На 9-й день бластоцисты теряют форму шара и несколько вытягиваются. В них закладываются и развиваются зародышевые листки. Эмбриобласт дифференцируется на наружный слой высоких клеток (эктодерму) и глубокий слой плоских клеток (энтодерму). Между этими слоями появляются клетки мезодермы. Происходит интенсивное разрастание трофобласта. Закладыва-
72
ется амнион — однослойная оболочка, окружающая зародыш. Клетки энтодермы быстро разрастаются по внутренней поверхности трофобласта, формируя стенки желточного мешка. В нем развиваются кровеносные сосуды. Желточный мешок обеспечивает избирательное всасывание необходимых зародышу питательных веществ. Зародыши становятся видимыми невооруженным глазом. Они представляют собой белые пленочки. В дальнейшем зародышевый пузырек сморщивается и удлиняется.
К 10-му дню зародыш имеет длину около 10—12, ширину — около 3 мм. На 13—14-й день происходит вытягивание зародыша, и он приобретает форму нити, достигающей длины 7 см. Диаметр его при этом уменьшается и составляет 104 мкм. Благодаря спиралеобразному расположению зародыш, несмотря на длину, занимает мало места. В дальнейшем -увеличение размеров зародыша происходит за счет его утолщения. Длина его при этом сокращается. Форма зародышей и их размеры сильно различаются даже у одной свиноматки.
После поступления в матку зародыши постепенно перемещаются по ее рогам. На 4-й день они продвигаются на 10—20 см, на 6—7-й день — на 40 см, на 8—9-й день — распределяются по всему рогу матки, а на 10-й день часть зародышей проникает в другой рог матки. На 11-й день наблюдаются Признаки прикрепления зародышей к эндометрию матки. На месте расположения зародышей в слизистой матки отмечается гиперемия. У трофобласта зародыша образуются выступы, которые входят в углубления слизистой матки. Имплантация заканчивается к 18-му дню.
К 14-му дню в зародыше появляются первые сомиты, из которых в дальнейшем образуются позвонки. Три зародышевых слоя к этому времени четко выражены. Продолжается клеточная специализация. Из эктодермы формируются наружный эпителий, млечные, сальные и потовые железы, щетинки, копыта, кишечный эпителий, эмаль на зубах и нервная система; из энтодермы — пищеварительный тракт, поджелудочная железа, печень, щитовидная железа, глотка, трахея, легкие, среднее ухо и околощитовидные железы; из мезодермы — скелет, скелетная мускулатура, соединительная ткань, кровеносные сосуды, сердце, соединительная ткань кровеносных сосудов, форменные элементы крови, кора надпочечников, плевра, брюшина, перикард, половые органы, почечные канальцы, мочеточники, гладкая мускулатура.
73
К 15-му дню желточный мешок достигает наибольшего развития. С этого времени усиленно развивается аллан-тоис, который выполняет функции желточного мешка. На 15—16-е сутки начинается интенсивное развитие эмбриобласта, и эмбрион быстро достигает длины 6—7 мм. На 20-й день его длина равна 10 мм. На 30-й день длина плодов равняется в среднем около 30 мм, а масса — около 3 г, на 45-й день эти показатели соответственно составляют 57 мм и 18 г, на 90-й день — 200 и 560 и на 105-й день — 220 мм и 850 г (табл. 5). Размеры и масса плодов имеют существенные индивидуальные различия. При рождении масса поросят колеблется от 400 до 2600 г. В период внутриутробного развития оболочки зародыша и эмбрион (плод) увеличиваются неравномерно. До 13-го дня трофо-бласт и амнион усиленно развиваются. В это же время увеличение эмбриобласта происходит сравнительно медленно. Его интенсивное развитие начинается с 15—16 суток. С этого времени масса эмбриона ежедневно возрастает в 2—3 раза. Средняя масса эмбрионов на 18-е сутки достигает 20 мг, а на 23-е — около 460 мг, т. е. в 13 раз больше. В дальнейшем интенсивность роста зародыша снижается. С 30-го по 45-й день его масса возрастает только в 6 раз, а с 60-го по 105-й день — меньше, чем в 8 раз. В то же время ежедневный абсолютный прирост массы возрастает с 45-го дня до конца супоросности.
Оболочки зародыша интенсивно развиваются в первой половине супоросности. На 17—19-е сутки длина плодных мешков достигает 40 см. К 40-му дню общая масса заро-
5. РАЗВИТИЕ ПЛОДОВ СВИНЕЙ
Возраст, дней Длина плода, см Масса плода, г
30 2,9 1,86
35 4,6 5,03
40 5,9 12,02
45 6,7 20,23
50 8,4 40,00
55 9,7 63,00
60 12,6 140,00
70 15,2 231,00
75 15,5 310,60
80 16,7 322,00
90 19,6 535,0
105 22,1 852,00
110 24,4 939,00
При рожде-
нии 25,8 1130,00
74
дышевых оболочек возрастает до 300—500 г, а к 60-му дню — до 2 кг и более. В дальнейшем рост зародышевых оболочек резко снижается и к 80-му дню супоросности их масса составляет в среднем 2,5 кг. После этого масса оболочек остается почти неизменной до опороса.
Количество аллантоисной жидкости на 20-й день супоросности — около 5 мл. К 45-му и 60-му дням ее объем возрастает соответственно до 113 и 350 мл, затем уменьшается. Количество амниотической жидкости возрастает с 26 мл на 45-й день до 62 мл на 60-й и достигает 250 мл на 80-й день. Затем ее объем снижается с возрастанием срока супоросности. Данные по массе оболочек, жидкости и матки в различные периоды супоросности приведены в таблице 6.
6. ОТНОСИТЕЛЬНАЯ МАССА ВНЕЗАРОДЫШЕВЫХ ОБОЛОЧЕК, ЖИДКОСТЕЙ И МАТКИ НА РАЗНЫХ СТАДИЯХ РАЗВИТИЯ ПЛОДА
|
| Оболочки
| Жидкость
| Матка
| ||||||||||
| День
| Число |
|
|
|
| процент от
| |||||||
| росно-
| плодов | г
| процент от массы
| г
| процент от массы
| г | массы у неоплодотво
| ||||||
|
|
| в 47 дней
|
| в 47 дней
| ренной сви
| ||||||||
|
|
|
|
|
| номатки
| ||||||||
| 47 | 12
| 800 |
| 1350
|
| 1300
| |||||||
| 63 | 11
| 2100 | 263
| 5050
| 374
| 2450
| 189 | ||||||
| 66 | 10
| 1570 |
| 4220
|
| 2450
| |||||||
| 81 | 12
| 2550 | 319
| 5650
| 419
| 2600
| 200 | ||||||
| 86 | 11
| 2150 |
| 2800
|
| 2550
| |||||||
| 96 | 10
| 2500 | 313
| 2250
| 207
| 3441
| 265 | ||||||
| 102 | 10
| 2500 |
| 1250
|
| 3200
| |||||||
| 108 | 9
| 2500 | 313
| 1890
| 140
| 3770
| 290 | ||||||
Масса матки во время супоросности постоянно возрастает. К 80-му дню она увеличивается в 2 раза, а к моменту опороса — в 3 раза. Масса матки во время супоросности существенно различается у разных свиней. Она зависит также от числа и размеров плодов. В свою очередь, размеры плода имеют положительную связь с массой плодных оболочек. Увеличение массы матки происходит за счет гипертрофии мышечных и эластичных волокон. В период супоросности отмечается и изменение состава матки. Содержание сухого вещества до 60-го дня снижается, а к 105-му дню вновь повышается. Содержание кальция возрастает в течение супоросности. Количество фосфора колеблется, увеличиваясь к концу супоросности. Содержание органических веществ, жира, азота, фосфора, нат-
75
рия,калия, магния к середине супоросности уменьшается, а затем возрастает до опороса.
Биохимический состав плодов отличается от состава материнского организма, что указывает на активное перемещение веществ через плаценту. С возрастом плода содержание в нем воды уменьшается. В 30 дней плоды содержат 94,7 % воды, а в 107 дней — 83,6 %. Уровень липидов повышается с возрастом зародышей от 0,5 % в 30 дней до 1,4 % — в 107 дней; соответственно уровень белка возрастает с 3,6 до 9,7 %. Химический состав плодов, полученных от одной свиноматки, мало отличается у разных поросят, несмотря на значительные различия в их массе. В то же время он существенно зависит от особенностей кормления свиноматок в период супоросности.

