




Слово гормон происходит от греческого и означает «двигаю», «возбуждаю», «раздражаю». Гормоны вырабатываются в определенных органах или тканях организма, выделяются в кровь и лимфу, разносятся по организму, усваиваются вне места их образования другими тканями, где вызывают существенные изменения в обмене веществ и функции органов. Гормоны оказывают выраженное действие в чрезвычайно малых концентрациях.
Физиологическая роль гормонов очень многообразна. Они регулируют все виды обмена веществ, вегетативные функции, ритм сердца, кровяное давление, работу почек, перистальтику кишечника, выделение пищеварительных соков, лактацию, морфогенез (рост и дифференциацию тканей), половое развитие организма, поддерживают постоянство внутренней среды организма (гомеостаз), воздействуют на развитие нервной ткани и т. д.
К настоящему времени открыто большое количество гормонов, число которых продолжает постоянно расти. Ввиду того, что железы, вырабатывающие гормоны, выделяют их в кровь, их называют эндокринными (секре-тирующими внутрь), а науку, изучающую гормоны,— эндокринологией. Гормоны имеют чрезвычайно различное химическое строение. Одни из них являются пептидами, содержащими от 3 до 200 аминокислотных остатков,
76
другие — аминами, третьи — стероидами. Одни из них оказывают действие на определенный орган, другие на различные органы и ткани. Одни из них существуют короткое время, всего несколько минут, другие значительно дольше. Одни гормоны вызывают в тканях-мишенях немедленный ответ. Ответная реакция на другие гормоны длится несколько часов и даже дней.
Кроме гормонов, вырабатываемых эндокринными железами, в тканях животного организма образуются биологически активные соединения, участвующие в регуляции различных функций. Такие соединения называются тканевыми гормонами, или парагормонами. К ним относятся гормоны плаценты, медиаторы нервного возбуждения, простагландины и др.
В клетках-мишенях гормоны связываются с так называемыми рецепторами гормонов, представляющими собой специфическую молекулу или группу молекул. Для гормонов, быстро проникающих в глубь клетки-мишени, рецепторы расположены в цитоплазме клеток. Для других гормонов рецепторы находятся на наружной поверхности клеточной оболочки. После присоединения гормона к рецептору в клетке образуется медиатор, который передает сигнал от рецептора к соответствующему ферменту или генам. В результате стимулируется их активность.
Во взаимодействии гормонов проявляется выраженная соподчиненность. На нижней ступени находятся периферические эндокринные железы, к которым относятся щитовидная, околощитовидная, тимус, надпочечники, поджелудочная и половые железы (яичники, семенники). Их функция регулируется центральной эндокринной железой — гипофизом. В свою очередь, гормональная функция гипофиза зависит от гормонов-регуляторов, выделяемых гипоталамусом. Взаимоотношения гормонов на различном уровне представляют собой очень сложную схему с многочисленными взаимосвязями. В упрощенном виде она представляет собой саморегулирующую систему, действующую по принципу обратной связи. Эту связь М. М. Завадовский (1941) определил как принцип плюс-минус взаимодействия. Она заключается в том, что при усилении выделения определенного гормона из гипофиза возрастает секреция соподчиненного ему гормона из периферической железы (плюс взаимодействие). Повышение же концентрации в крови гормона периферической железы приводит к торможению выделения гипофизарного гормона (минус взаимодействие). В результате такого взаимо-
77
действия основного и подчиненного гормонов в организме устанавливается определенный уровень обоих гормонов. Эта схема позволяет объяснить и предвидеть многие гормональные взаимоотношения в организме. Однако с позиций указанной теории находят объяснения не все известные факты. Так, в некоторых случаях наблюдается взаимодействие гормонов по принципу плюс-плюс, т. е. повышение концентрации гормона периферической железы вызывает усиление секреции гипофизом основного гормона. Подобные явления не всегда находят объяснение и свидетельствуют о недостаточном уровне наших знаний. Отклонения от взаимоотношений гормонов по принципу плюс-минус взаимодействия объясняются в настоящее время в основном тем, что функция самого гипофиза регулируется гипоталамусом, а на его активность оказывают влияние и гормоны периферических желез, и гормоны гипофиза, и нервные импульсы, поступающие из центральных и периферических отделов нервной системы.
ГОРМОНЫ СЕМЕННИКОВ
Андрогены. После удаления семенников у молодых петушков Бертхольд (1849) наблюдал уменьшение и по-бледнение гребня и сережек. Пересадка таким петушкам семенников (или их кусочков) устраняла послекастраци-онные изменения. Этим было доказано, что в семенниках вырабатывается вещество, оказывающее влияние на другие органы и ткани организма. Так как с этим веществом были связаны отличительные особенности самцов, то его назвали мужским половым гормоном.
Более поздние исследования показали, что у кастрированных в раннем возрасте самцов остаются неразвитыми половые органы, не проявляется половое влечение, меняется телосложение, отсутствует специфический запах, не образуются утолщения кожи и волосяной покров, не разрастаются клыки.
Признаки, которые зависят от деятельности семенника, были названы вторичными половыми признаками. У взрослых животных кастрация приводит к обратному развитию вторичных половых признаков. Помимо влияния на вторичные половые признаки половой гормон оказывает действие на обменные процессы в организме, ускоряет развитие молодых животных, усиливает рост скелетных мышц, способствует задержке и отложению в тканях белков и минеральных веществ. Мужской половой гормон
78
способствует переживаемости спермиев в придатках семенников. Половые гормоны регулируют выработку фруктозы и лимонной кислоты пузырьковидными железами.
Половой гормон оказывает значительное влияние на нервную систему, участвует в создании половой доминанты, т. е. господствующего очага возбуждения в центральной нервной системе. После кастрации животные становятся более спокойными, у них изменяются многие рефлексы. Если животных кастрируют в молодом возрасте, то половое возбуждение и половые рефлексы у них не проявляются, если удаление семенников проводят у взрослых животных, то половая активность у них вначале сохраняется или даже повышается, а затем постепенно снижается и исчезает. Введение таким животным тестостерона восстанавливает их половую активность и половые рефлексы, а у самки вызывает проявление половых рефлексов по типу самца. Это действие андрогенов особенно выражено, когда их вводят кастрированным самкам. Введение тестостерона беременным самкам не вызывает маскулинизации у них полового поведения. Это связано с высоким уровнем прогестерона, который тормозит влияние андрогенов на нервную систему.
Введение тестостерона неполовозрелым самцам приводит к их половому возбуждению и проявлению половых рефлексов. Половой гормон определяет развитие половых органов у эмбрионов по мужскому типу и воздействует на половую дифференцировку гипоталамуса.
Исследования с основным из андрогенных гормонов тестостероном показали, что он в больших количествах необходим для процессов сперматогенеза. Половой гормон поступает в клетки Сертоли и задерживается там с помощью тестостероисвязывающего белка. Отсюда он передается половым клеткам, у которых имеются рецепторы тестостерона. С помощью инъекций тестостерона у самцов можно поддерживать сперматогенез даже после удаления у них гипофиза.
Химические исследования показали, что свойствами полового гормона обладает целая группа стероидов, которые были названы андрогенами. К ним относятся андростерон, дегидроандростерон и тестостерон. Наиболее активен из адрогенов тестостерон. Он в 6 раз активнее андростерона, который, в свою очередь, в 3 раза активнее дегидроандро-стерона. Все указанные гормоны являются производными циклического углеводорода — циклопентанопергидрофе-нантрена и, следовательно, близки к холестерину (рис.
79
г» клетках-мишенях из тестостерона образуется более ,.-, —явный половой гормон — дегидротестостерон. Суточная::• нунция тестостерона увеличивается с возрастом хряков.
- ' месяца она составляет 0,3—0,5 мг, в 9 месяцев —
г: возрасте 5 лет — 10 мг. Концентрация тестостерона 1 г-.1азме крови семенной вены резко повышается с 4- до '--м-е-сячного возраста. В возрасте 2 месяцев она равняется
-римерно 20 нг/мл, в 4—80, в 12 месяцев — 400 нг/мл. В "^риферической крови взрослых хряков концентрация
--:т:и:терона составляет в среднем 2—5 нг/мл. У жирных хряков концентрация гормона в плазме крови оказывается '•"лее высокой, что связано с его накоплением в жировой ткани. В крови тестостерон находится в свободной или связанной форме. Он может быть связан с белками сыворотки крови (неактивная форма) или находиться в выделительной форме (глюкуронид, сульфатконъюгат).
Эстрогены. В семенниках хряков образуется чрезвычайно большое, несвойственное самцам других видов животных (за исключением жеребцов) количество эстрогенов. Известно, что эстрогены являются антагонистами эндро-генов и экзогенное введение их самцам нарушает воспроизводительную функцию производителей. Однако в небольших количествах эстрогены вырабатываются в организме самцов всех видов сельскохозяйственных животных. В то же время в организме самок помимо эстрогенов вырабатываются и небольшие количества андрогенов. У хряков эндрогенные эстрогены не только не тормозят половую функцию, но и участвуют в деятельности придаточных половых желез, в проявлении половых рефлексов и обладают анаболитическим действием. Эстрогены найдены в значительных количествах в плазме спермы.
В эякулятах хряка находится 5 мкг 17р-эстрадиола, 2 мкг эстрона и 4,5 мкг эстрон сульфата. Количество эстрогенов в сперме имеет существенные индивидуальные и сезонные различия. Осенью их концентрация в сперме в 10 раз выше, чем летом.
В настоящее время считается, что между этими группами гормонов существует не только антагонизм, но и синергизм, и взаимодействие между ними связано с их различной концентрацией в мужском и женском организме.
Эстрогены вырабатываются из тестостерона. Основными эстрогенами, синтезируемыми в семенниках, являются 17В-эстрадиол и эстрон. У взрослых хряков в 1 л мочи находят до 1—Змг эстрогенов, что значительно выше, чем у
81
несупоросных свиноматок. Концентрация эстрогенов в крови хряков также выше, чем у маток.
Ингибин. В семенниках также вырабатывается еще один гормон — ингибин. Его наличие установлено опытными исследованиями, в которых после определенных воздействий на половые железы резко возрастало выделение ФСГ гипофизом. Уровень тестостерона в организме при этом не нарушался. Например, это происходило при введении животным бисульфана, разрушающего клетки сперматогенного эпителия, при искусственном криптор-хизме, при искусственном перегревании семенников. Исследования ин витро с тканями клеток Сертоли показали, что ингибин вырабатывается именно этими клетками. Указанный гормон тормозит выделение ФСГ из гипофиза. Химическая природа его не установлена. Известно только, что он представляет термолабильную макромолекулу.
Феромон андростенон. Существенной субстанцией, вырабатываемой в семенниках хряка, является стероид, который относится не к гормонам, а к феромонам. По строению он очень сходен с тестостероном, но образуется независимо от него и не служит андрогеном. Его называют андростеноном. Химически он представляет собой 5а -анд-рост-16-ен-З-он. Феромоны служат у животных средствами связи между различными индивидуумами и вызывают у чувствительных к ним животных физиологические или психические процессы. У хряка феромон служит средством сигнализации о наличии самца для самок, находящихся в стадии половой активности. В результате самки находят самца на значительном расстоянии от них. Феромон благодаря своим липофильным свойствам накапливается в жировой ткани. При значительном его количестве в мясе оно становится непригодным в пищу из-за неприятного запаха. Этот запах особенно усиливается при варке мяса, что является одним из тестов его присутствия. В результате наличия андростенона мясо хряков (в отличие от самцов других видов животных) часто оказывается непригодным для использования.
Концентрация андростенона в организме находится в тесной связи с содержанием в нем тестостерона. Коэффициент корреляции между ними равняется 0,64. В плазме крови его количество измеряется в нанограммах, а в жировой ткани и в слюнных железах, куда феромон поступает из семенников, его концентрация выражается в миллиграммах на 1 г ткани. У 175-дневного хряка количество феромона в 1 г жировой ткани составляло
82
1,03 мг, в слюнной железе — 1,17 мг. У хряка в возрасте 217 дней эти показатели равнялись соответственно 7,49 и 11,43 мг, у хряка в возрасте 229 дней — 1,74 и 1,99 мг.
При низкой концентрации вещества в крови выделение феромона из жировой ткани происходит медленно. У хряков массой 100 кг запах в мясе исчезает через 3—4 недели после кастрации. С поступлением феромона в слюнные железы связано его участие в процессах воспроизводства. Особенно большое количество феромона вырабатывается в семенниках поздней осенью, когда у диких кабанов наступает половой сезон.
Ингибитор развития Мюллерова канала. В фетальных, а возможно, и в постнатальных семенниках образуется также субстанция, определяющая регрессию Мюллерова канала в период эмбрионального развития. Это установлено в опытах с кастрацией эмбрионов и инъекцией им тестостерона или при введении матерям веществ, обладающих антиандрогенным действием.
ГОРМОНЫ ЯИЧНИКОВ
Эстрогены. В яичниках вырабатывается несколько гормонов, которые необходимы для осуществления половой функции самок. Их называют женскими половыми гормонами. Ранее к ним относили только группу стероидных гормонов, с которыми было связано развитие половых признаков. Эта группа гормонов была названа эстрогенами. При кастрации самок введение им эстрогенов способствовало развитию вторичных женских половых признаков. Эстрогены образуются во всех клеточных элементах фолликула и в интерстициальной ткани яичников, в небольшом количестве — в коре надпочечников.
Эстрогены выполняют в организме многочисленные функции. В период полового созревания резко повышают у самок рост яйцепроводов, влагалища, молочных желез, пробуждают половой инстинкт. В зрелом организме они вызывают ороговение и отторжение эпителиальных клеток влагалища, стимулируют рост эндометрия и маточных желез, усиливают васкуляризацию матки и активность ее миометрия, способствуют развитию и дифференциации тканей в вымени, росту и развитию матки. Эстрогены влияют и на рост костей, с чем связано развитие телосложения по женскому типу. Они тормозят рост трубчатых костей и способствуют окостенению эпифизарных хрящей. Эстрогены стимулируют синтез белков, жиров,
83
гликогена, задерживают в организме глюкозу, натрий, кальций, фосфор и воду, снижают активность сальных желез. Эстрогены определяют половое поведение самок. Введение эстрогенов кастрированным самкам ведет к появлению у них признаков половой охоты.
Химические исследования показали, что свойствами эстрогенов обладает группа гормонов. В настоящее время из организма животных выделено и изучено более 12 гормонов, обладающих эстрогенным действием. Все они — стероиды. В основе их строения лежит углеводород цикло-пентанопергидрофенантрен и его производное — углеводород эстран с 18 углеродными атомами и метильной группой в положении 13. Эстрогены образуются из тестостерона. Наиболее часто в организме встречаются эстрон, а - и р -эстрадиол и эстриол. Эстрон также называют фолликулином. Указанные гормоны отличаются по структуре и активности. Эстрадиол существует в двух изомерах, отличающихся химическими и биологическими свойствами: 17р -эстрадиол в 40 раз активнее 17а -эстра-диола, в 8—10 раз — эстрона и в 100 раз — эстриола.
Для измерения активности эстрогена предложена мышиная единица. Под ней понимают минимальное количество эстрона, которое при шестикратном введении в течение 48 ч вызывает течку минимум у 75 % кастрированных самок мышей. За международную единицу принято считать 0,1 мкг стандартного препарата кристаллического эстрона.
В процессе биосинтеза эстрогенов наблюдается их взаимопревращение. Эстрадиол легко превращается в эстрон и наоборот. Инактивация эстрогенов осуществляется в печени. Здесь происходит их соединение с серной и глю-куроновой кислотой. Эстрогены и образующиеся из них соединения выводятся из организма.с мочой и калом.
Эстрогены широко распространены в природе. Их находят у насекомых и во многих растениях. Растительные эстрогены называются фитоэстрогенами.
Мишенями для эстрогенов служат матка и влагалище. Помимо этого они накапливаются также в печени, коре надпочечников, вымени, гипоталамусе, гипофизе и других органах. При введении эстрогенов увеличиваются масса матки и яичников в результате повышения в этих органах синтеза белка и нуклеиновых кислот. В свою очередь, клетками-мишенями для эстрогенов в матке являются эпителиальные клетки слизистой оболочки.
Стимуляция белкового синтеза матки происходит че-
84
рез 4 ч после введения эстрогенов. Максимума она достигает через 8 ч и совпадает с максимальной фиксацией эстрогенов. После этого синтез белков сохраняется на высоком уровне в течение 72 ч после введения гормонов.
Механизм действия эстрогенов в клетках-мишенях заключается в том, что они связываются в тканях специфическими внутриклеточными рецепторами. Эстрогенный рецептор называется эстрофилином I и имеет молекулярную массу около 200 000. При связывании молекулы эстрогена эстрофилин I претерпевает молекулярные изменения и превращается в эстрофилин II — вторичный посредник в действии эстрогена. Эстрофилин II проникает в клеточное ядро, где взаимодействует с хроматином, вызывая активизацию определенных генов и синтез специфических белков. Это приводит к характерным изменениям в тканях репродуктивной системы.
Ранее основной функцией эстрогенов считалось развитие вторичных половых признаков у самки и изменения в организме, обеспечивающие ее оплодотворение. В настоящее время установлено, что эстрогены выполняют чрезвычайно различные функции в организме в процессе всего воспроизводительного цикла. Эстрогены принимают участие в дифференцировке половых органов у зародышей, они необходимы для сохранения супорос-ности, осуществления родового процесса и т. д.
Прогестерон. Другой гормон, вырабатываемый в яичнике — прогестерон. Прогестерон вырабатывается в желтом теле яичника. По химическому строению он очень близок к андрогенам и является производным углеводорода прогнана. В его молекуле имеются две кетогруппы, три метильные группы и одна двойная связь. Его основное значение состоит в подготовке эндометрия матки к приему и имплантации оплодотворенной яйцеклетки, а также сохранению супоросности. Под его влиянием повышается секреторная функция эндометрия и слизистой влагалища. Он тормозит сокращения миометрия. Прогестерон усиливает развитие молочной железы. В основном прогестерон по своему действию антагонист эстрогенов. Однако в некоторых случаях в их действиях отмечается синергизм. Так, небольшие количества прогестерона резко активизируют действие эстрогенов в эротизации нервных центров и способствуют таким образом возбуждению самок. Прогестерон и эстрогены усиливают развитие молочной железы, секрецию желез матки. Прогестерон в организме может превращаться в неактивную
85
форму — прегнандиол, который соединяется с глюкуро-новой кислотой и выводится с мочой.
Релаксин. Релаксин — гормон, участвующий в подготовке и течении родового процесса. Релаксин — поли-пептид с молекулярной массой 6000. Молекула гормона состоит из двух неидентичных цепей, одна из них содержит 22, а другая 32 аминокислотных остатка. По строению он очень сходен с инсулином. Он вырабатывается в желтом теле в течение всей супоросности, но выделяется в кровь в больших количествах только с началом подготовки половых органов к родам. Релаксин расслабляет соединения тазовых костей, особенно лонного сочленения, матка и влагалище увеличиваются в размерах, содержание в них воды возрастает. Повышается растяжимость тканей и связок. Шейка матки расширяется и размягчается. Релаксин снижает тонус матки и ее сократительную деятельность. Действие его лучше проявляется на фоне эстрогенов, оно наступает быстро, через несколько часов. Известно, что прогестерон делает матку не чувствительной к окситоцину. Окситоцин в период супоросности не вызывает сокращения миометрия матки. Релаксин снимает это тормозящее действие прогестерона на матку.
Учитывая, что релаксин вырабатывается в желтом теле беременности и не образуется в циклическом желтом теле, его считали гормоном беременности. Однако в последующем релаксин был найден в крови небеременных самок и даже в крови самцов.
Ингибин. В фолликуле вырабатывается также гормон ингибин. Он оказывает тормозящее влияние на секрецию ФСГ гипофизом. Можно полагать, что он имеет такую же структуру, как и ингибин, вырабатываемый в семенниках.
Тестостерон. В фолликулах яичника вырабатывается тестостерон, который является непосредственным предшественником эстрогенов. В небольших фолликулах количество тестостерона превышает концентрацию эстрогенов. По мере созревания фолликулов содержание тестостерона в них снижается, а концентрация эстрогенов повышается, особенно в предовуляторный период. При повышенной концентрации тестостерона наступает атрезия фолликулов. При нормальном соотношении гормонов развитие фолликулов полностью завершается.
ГОРМОНЫ МАТКИ
Важная функция матки — выработка биологически активных веществ, которые вызывают регрессию желтого
86
тела в яичниках. При удалении матки желтое тело функционирует в течение продолжительного периода времени. У свиней оно сохраняется в течение 140 дней, т. е. дольше, чем продолжается вся супоросность (100— 130 дней). Минимальная часть матки, которая необходима для осуществления лизиса желтого тела, составляет '/4 часть одного рога матки. Перенос вещества, лизирую-щего желтое тело, в яичники осуществляется только при сохранении сосудистой связи между ними. Если яичник отделяли от матки и переносили в другой участок организма, то лютеолитическое действие матки не сохранялось. Если яичник переносили в другой участок тела вместе с маткой, то ее лютеолитическое действие проявлялось. Этим было доказано, что вещество, вызывающее лютеолиз желтого тела, поступает из матки не в общий кровоток, а только в сосуды, соединяющие матку с яичником. Этот путь оказался необычным. Лютеолизин, образующийся в матке, попадает в маточную вену, затем по анастомозам в артерию яичника. Это свидетельствует о том, что кровоток в яичниковой артерии возможен в обратном направлении. Исследования показали, что эта связь является локальной, т, е. перенос лютеолизина осуществляется только в яичник, расположенный на стороне соответствующего рога матки.
Химические исследования показали, что фактор, вырабатывающийся в матке и вызывающий лютеолиз желтого тела, является простагландином. Простагландины были известны давно. Они впервые были выделены из спермы человека и барана, в которой они содержатся в значительных количествах. Простагландины вырабатываются в предстательной железе (простата), откуда они и получили свое название. Определено, что Простагландины усиливают маточные сокращения. Лютеолитический эффект простагландинов в то время не был известен.
Химические исследования показали, что Простагландины представляют собой жирные кислоты с 20 углеродными атомами, содержащие циклопентановое кольцо. Они образуются из полиненасыщенных жирных кислот (линолевой, линоленовой, арахидоновой и др.). Имеются две основные группы простагландинов — Е и Г, различающихся по структуре и действию. Из простагландинов указанных групп образуется несколько вторичных простагландинов. Простагландины синтезируются в различных тканях и обладают многосторонним, иногда, в зависимости от типа, противоположным действием. Они
87
стимулируют или снижают активность аденилатциклазы, особенно в эндокринных железах, активизируют образование гормонов в надпочечниках, освобождение инсулина из поджелудочной железы, синтез и выделение гормона щитовидной железы, увеличивают или уменьшают кро-воток, усиливают ионный поток через эпителиальные мембраны, повышают температуру тела, обладают седа-тивным и транквилизирующим действием, модулируют действие адренергических факторов, тормозят желудочную секрецию, но усиливают секрецию в поджелудочной железе и кишечнике, воспалительную реакцию, моторику кишечника и матки, выделение окситоцина из гипофиза, вызывают расслабление или сокращение мышц бронхов и трахеи, тормозят или усиливают иммунитет, увеличивают внутриглазное давление.
Влияние на воспроизводство оказывают в основном простагландины группы Г. Ответственный за лютеолиз желтого тела простагландин Г 2а вызывает сужение сосудов яичников, в результате чего их кровоснабжение снижается на 30 %. У свиней при его введении наблюдаются такие побочные явления, как повышение кровяного давления, возбуждение кишечной перистальтики, спазм бронхов и внезапное выделение пота. Простагландины в крови быстро разрушаются. При прохождении крови через легкие и печень инактивируется до 90 % простагландинов. Период полураспада биологических простагландинов составляет 45 с. Простагландины способны вызвать лютеолиз желтого тела у свиней только с 11—12-го дня полового цикла.
Простагландины вырабатываются практически во всех органах и тканях организма. В отличие от простаглан-дина Г 2а простагландин Е способствует развитию и сохранению желтого тела.
ГОРМОНЫ ГИПОФИЗА
Гипофиз представляет собой маленькое плотное образование округло-уплощенной формы, бурого цвета, соединенное с дном промежуточного мозга тоненькой ножкой (рис. 11), покрыт твердой мозговой оболочкой и лежит в ямке турецкого седла у основания черепа. Его масса у свиней составляет 0,3—0,5 г. Гипофиз состоит из трех частей (долей), которые различаются по структуре, функции и происхождению. Передняя, наиболее развитая доля возникает из эктодермы глотки путем отшнуровыва-
88
ния и присоединения затем к основанию мозга. Она имеет железистую структуру. Задняя доля, или нейрогипофиз, несколько меньше передней, состоит из нервных структур, является производным дна третьего мозгового желудочка и продолжением нижнего отдела промежуточного мозга — гипоталамуса, соединение с которым представляет вид воронки. Средняя, или промежуточная, доля имеет общее происхождение с передней долей. Она эпителиальной структуры и в виде тонкой полоски рас-
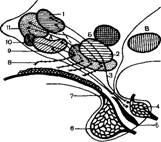
Р и с. 11. Структурная схема гипоталамо-гипофизарной системы:
А — ядра переднего гипоталамуса: / — супраоптические; 2 — вентромедиальные;
3 — аркуатные; 4 — задняя доля гиыо-физа; Б — ядра среднего гипоталамуса:
5 — средняя доля гипофиза; 6 — передняя доля гипофиза; В — ядро заднего гипоталамуса; 7 — воротные сосуды; 8 — первичная капиллярная сеть; 9 — паравен-трикуляряые ядра; 10 — супрахиаэмати-ческие ядра; 11 — преоптические ядра
полагается между передней и задней долями. Учитывая различное происхождение и структуру, гипофиз иногда подразделяют на аденогипофиз (передняя и средняя доли) и нейрогипофиз (задняя доля).
Несмотря на свой маленький размер, гипофиз вырабатывает более 10 различных гормонов, которые регулируют деятельность всех желез внутренней секреции и ряд важнейших функций организма. Количество обнаруженных в гипофизе гормонов постоянно растет и возможно открытие новых. Соединение в важнейшем регуляторе функций организма железистой и нервной секреторной тканей — еще одно свидетельство единства нервной и гормональной систем организма.
Из вырабатываемых гипофизом гормонов известны четыре, которые непосредственно влияют на воспроизводительную функцию животных. Три из них образуются в передней доле и ввиду своего влияния на развитие и функцию половых желез (гонад) называются гонадотроп-ными. К ним относятся: фолликулостимулирующий гормон (ФСГ), лютеинизирующий гормон (ЛГ) и лактоген-ный гормон.
Четвертый гормон — окситоцин — вырабатывается в задней доле гипофиза.
Из передней доли гипофиза были выделены два го-надотропных гормона. Один из них был назван фоллику-
89
лостимулирующим, так как стимулировал рост фолликулов в яичниках самок (продан А), а другой — лютеини-зирующим (продан Б), так как он способствовал овуляции фолликулов и их лютеинизации. Ввиду того, что этот гормон вызывает у самцов развитие интерстициаль-ной ткани семенника, его иногда называют также интер-стициальным.
ФСГ и ЛГ являются глюкопротеидами. Очищенные гормоны, полученные из гипофизов различных видов мле- | копитающих, сходны по биологическому действию, но > отличаются по некоторым физическим и химическим | характеристикам. Молекулярная масса ФСГ овцы достигает 67 000, свиньи 29 000. Очень богат ЛГ гипофиз овцы. Концентрация в нем ЛГ в 5 раз выше, чем в гипофизе свиньи, и в 12 раз выше. чем в гипофизе.коровы. Молекулярная масса ЛГ гипофиза овцы равна 40 000, свиньи — 100 000. ^
В последние годы в связи с выделением очищенных препаратов ФСГ и ЛГ были получены данные по их более тонкому действию на половые железы. Так, оказалось, что ФСГ в чистом виде неспособен вызывать рост и развитие фолликулов. Для проявления этого известного свойства ФСГ необходимо наличие небольших количеств ЛГ. В то же время и известное свойство ЛГ, заключающееся в его регуляции синтеза гормонов в семенниках и яичниках, не проявляется без участия ФСГ. ФСГ, с одной стороны, обеспечивает развитие клеток, которые в дальнейшем под влиянием ЛГ продуцируют гормоны, а с другой — ФСГ стимулирует активность ферментов, способствующих синтезу гормонов. Например, ФСГ стимулирует " действие ароматизирующих ферментов, осуществляющих последний этап в биосинтезе эстрогенов, их образовании из андрогенов. Таким образом, подтверждаются ранее известные данные о том, что все воспроизводительные процессы в половых органах происходят при совместном воздействии двух важнейших гонадотропных гормонов гипофиза ФСГ и ЛГ. Указанные гормоны никогда не выделяются из гипофиза изолированно. До сих пор остается также неясным, вызывают ли образование и выделение * обоих гормонов из гипофиза один или два рилизинг-фактора гипоталамуса.
Лактогенный гормон. Помимо указанных двух гормонов в передней доле гипофиза вырабатывается и третий гона-дотропный гормон — лактогенный (пролактин, лютеотроп-ный). Лактогенным он был назван потому, что гормон
90
оказывает влияние на развитие молочной железы и лактацию. Он также поддерживает функцию желтого тела у крыс и мышей. Из-за этого действия он был назван также лютеотропным. Гормон не содержит Сахаров и не является глюкопротеидом. Его молекулярная масса равна примерно 23 000—32 000, т. е. мало отличается от молекулярной массы ЛГ и ФСГ. У хряков лактогенный гормон в синергизме. с тестостероном и эстрогенами стимулирует функцию придаточных половых желез.
Окситоцин. В задней доле гипофиза содержится гормон окситоцин, в переводе с греческого языка означает «быстрые роды». Он действует на некоторые гладко-мышечные ткани, в особенности на мускулатуру матки, в меньшей степени стимулирует мускулатуру мочевого пузыря и кишечника.
В то время как действие окситоцина на мускулатуру матки давно и хорошо известно, имеются только отдельные сообщения о влиянии окситоцина на сокращение половых органов самцов.
При внутривенном введении окситоцина кроликам наблюдали ритмичные сокращения семенника, придатка и спермиопровода. Движения семенника и придатка были связаны с сокращением их оболочек, содержащих гладкую мускулатуру. При введении окситоцина усиливалось выделение спермы кроликами. Между содержанием окситоцина в крови быков во время полового возбуждения и качеством получаемого эякулята установлена прямая связь. Инъекции окситоцина быкам и хрякам перед или во время полового акта повышали количество выводимых в эякулят спермиев.
Окситоцин является полипептидом, состоящим из восьми остатков аминокислот: цистина, тирозина, пролина, глютаминовой и аспарагиновой кислот, глицина, лейцина и изолейцина. Молекулярная масса окситоцина составляет 1070. Окситоцин вызывает выделение молока сразу после его введения, благодаря сокращению миоэпители-альной ткани молочной железы.
Помимо окситоцина в задней доле гипофиза содержится другой гормон — вазопрессин, который оказывает антидиуретическое действие и повышает кровяное давление. Несмотря на чрезвычайно различное действие, они имеют очень сходное химическое строение, в результате чего их трудно было отделить друг от друга. Только в 1949 г. биохимику Дю Виньо удалось выделить из гипофиза животных чистый окситоцин. Через несколько лет была изу-
91
чена его структурная формула, а в 1953 г. ученый синтезировал окситоцин, который по действию не отличался от окситоцина из натуральных источников. Исследования показали, что окситоцин образуется не в задней доле гипофиза, а в паравентрикулярном ядре гипоталамуса, откуда он поступает в заднюю долю гипофиза. Здесь он накапливается и выделяется в кровь.
Окситоцин выводится из гипофиза в ответ на раздражение половых органов самки при коитусе или искусственном осеменении, а также при раздражении молочной железы при сосании или доении. Помимо влияния на гладкую мускулатуру воздействие окситоцина на воспроизводительную функцию заключается в его участии в регуляции функции передней доли гипофиза. ,
После введения окситоцина удавалось устранить нарушение гонадотропной активности гипофиза, связанное с повреждением гипоталамуса или перерезкой ножки «гипофиза. Установлено, что при раздражении вымени 1 усиливается выделение окситоцина, стимулирует выделение лактогенного гормона из передней доли гипофиза.
Путь, по которому воздействует окситоцин на перед- ' нюю долю гипофиза, недостаточно ясен. По мнению некоторых авторов, тесная сосудистая связь между передней и задней долями гипофиза имеет функциональное значение.
Помимо указанных действий окситоцин, вероятно, оказывает влияние и на некоторые биохимические процессы в организме. При его введении собакам наблюдалось увеличение выделения натрия и хлора с мочой. Введенный внутривенно окситоцин быстро разрушается;;
в крови. Гормональная активность крови крыс исчезала через 2 мин после его внутривенного введения. Действие окситоцина на мускулатуру половых органов самцов продолжалось в течение 7—8 мин после его внутривенного введения.
Роли средней доли гипофиза в половой функции до последнего времени не придавалось значения. Известно, что в ней вырабатывается меланоцитостимулирующий гормон, который регулирует пигментацию покровов жи- ^ вотных. Однако в последние годы установлено, что в средней доле гипофиза образуются также дериваты семейства (3 -липотропинов, к ним относятся эндогенные опиаты (эндорфи, энкефалин). Указанные соединения оказывают влияние на гонадотропную активность передней доли гипофиза.
92
Гонадотропин-рилизинг-гормон гипоталамуса. При нарушении связи между гипоталамусом и гипофизом, в случае перерезки ножки гипофиза, наблюдалось резкое изменение гонадотропной функции гипофиза и подавление секреции тестостерона. В аденогипофизе омертвлялась ткань, сохранившиеся клетки утрачивали дифференциров-ку, регенерации паренхимы не наблюдалось, промежуточная доля гипертрофировалась.
Гипоталамус (см. рис. 11) представляет собой часть промежуточного мозга, его подбугорный отдел. Условно он подразделяется на переднюю, среднюю и заднюю части. Он имеет тесную связь с ретикулярной формацией, через которую осуществляется взаимодействие с корой больших полушарий головного мозга. Подбугорье является ведущим центром вегетативной, или автономной, нервной системы. Помимо половой функции подбугорье регулирует также сердечную и сосудистую деятельность, сон, терморегуляцию, водный, углеводный и жировой обмены, деятельность желудочно-кишечного тракта, почек и т. д. В гипоталамусе имеется 32 пары нервных ядер. Связь гипоталамуса с гипофизом осуществляется за счет своеобразного кровоснабжения гипофиза.
Отходящие от артериального круга большого мозга артерии гипофиза разделяются на 20—25 артериол, которые направляются в гипоталамус. Здесь вблизи серого бугра они делятся на капилляры и образуют первичную капиллярную сеть. Капилляры этой сети образуют петли и клубочки. Затем капилляры первичной сети собираются в портальные вены, которые в количестве 10—20 идут вдоль гипофизарной ножки в паренхиму аденогипофиза, где вновь рассыпаются на вторичную сеть капилляров-синусоидов, ветвящихся между трабекулами передней доли.
Исследованиями установлено, что два крупных ядерных образования гипоталамуса — супраоптические и пара-вентрикулярные ядра — обладают процессами нейросекре-ции. Секреторные нейроны этих ядер направляются в заднюю долю гипофиза, формируя мощный гипоталамо-гипофизарный тракт. В него входит до 100 000 нервных волокон. На протяжении всего гипоталамо-гипофизарного тракта нейроны имеют тесный контакт с эндотелием кровеносных сосудов. Поток нейросекрета в нейронах направлен от гипоталамуса к гипофизу, а не наоборот, как это считали ранее некоторые исследователи. Здесь нейросекрет накапливается и выделяется в виде гормонов
93
задней доли гипофиза. Нейросекрет гипоталамуса, направляющийся по портальной системе сосудов в аденоги-пофиз и приводящий к выделению гонадотропных гормонов из гипофиза, отличается по своему строению от нейро-секрета, поступающего в заднюю долю гипофиза. Ввиду того, что он способствует выделению гонадотропных гормонов, был назван гонадотропин-рилизинг-гормоном Гн-РГ (рилизинг от английского высвобождать). Его называют также гонадолиберином, люлиберином. Помимо Гн-РГ в гипоталамусе был найден гормон, высвобождающий пролактин (пролактолиберин) и ингибирующий выделение пролактина (пролактин-ингибирующий гормон). Гонадолиберин представляет собой пептид с молекулярной массой около 2000. В свином Гн-РГ, выделенном из гипоталамуса, определена следующая последовательность аминокислот: пироглютаминовая кислота, гистидин, триптофан, серин, тирозин, глицин, лейцин, аргинин, пролин.
Синтезированный в 1971 г. рилизинг-гормон способствует выделению из гипофиза как ЛГ, так и ФСГ. Однако в основном он вызывает выделение из гипофиза ЛГ. Для выделения ФСГ требуются большие дозы гормона. Эти данные наводят на мысль о том, что или действие гона-долиберина зависит от различного фона в организме периферических гормонов, или из гипоталамуса был выделен и изучен фактор, вызывающий выделение из гипофиза ЛГ, а фактор, вызывающий ФСГ, остался неизвестным. О том, что в гипоталамусе, вероятно, образуются два различных гонадотропин-рилизинг-гормона, свидетельствуют опыты с повреждением различных зон гипоталамуса. Так, при повреждении супрахиазматических участков гипоталамуса у крыс наблюдалась длительная течка, в яичниках происходило кистозное перерождение фолликулов, желтые тела отсутствовали, в эндометрии отмечалась кистозная гиперплазия. При повреждении области паравентрикулярных ядер удлинялась стадия полового покоя, отмечались ги-перлютеальные процессы в яичниках, прогестационные изменения в эндометрии. Эти исследования свидетельствовали о том, что в первом случае тормозится секреция лютеинизирующего гормона и повышается выделение фол-ликулостимулирующего гормона, а во втором — повышается секреция ЛГ.
Указанные исследования свидетельствовали также о том, что помимо образования в разных участках гипоталамуса двух различных рилизинг-гормонов, в гипотала-
94
мусе имеются факторы, тормозящие их образование или выделение из нейронов. Разрушение таких зон приводило или к повышению уровня ЛГ-рилизинг-гормона или ФСГ-рилизинг-гормона.
В результате других исследований выявлено, что срединное возвышение и задний гипоталамус стимулируют выделение из гипофиза ФСГ и тормозят выделение ЛГ, а в переднем гипоталамусе стимулируется выделение из гипофиза ЛГ.
При изучении влияния на гипоталамическую активность некоторых веществ определено, что препараты меди (особенно глюконат меди, тестостерон, ацетилхолин, галлоиды, поваренная соль, бромистый натрий и другие стимулируют секреторную активность гипоталамуса, а йодистый натрий и допаминреактивные структуры вызывают застой нейросекрета в телах нервных клеток. Если нервные структуры гипоталамуса выключались гипотала-мическими анестетиками, то действие всех указанных соединений не проявлялось.
В гипоталамусе найдены различные области, ответственные за тоническое (у самцов) или циклическое (у самок) выделение гонадотропных гормонов из гипофиза. В первом случае ведущую роль играет медиобазальный гипоталамус и аркуатное ядро, во втором — преоптическая область, передний гипоталамус, супрахиазматическое ядро и другие ядра.
Гипоталамус ответственен за различие в гонадотроп-ной функции гипофиза у самцов и самок. О том, что эта функция зависит не от гипофиза или половых органов взрослых животных, свидетельствуют опыты, в которых гипофиз самца, трансплантированный гипофизэктомиро-ванным самкам, способствовал нормальному половому циклу.
Секреция оказалась одной из важнейших функций клеток ядер гипоталамуса. Нейросекреторные гранулы синтезируются в нейроплазме тела клеток, в их отростках и расширениях. Нейросекреторные гранулы представляют собой липопротеиды или липогликопротеиды, покрытые фосфолипидной мембраной. Образовавшиеся гранулы концентрируются в аппарате Гольджи и формируются в готовые гранулы. Они перемещаются по отросткам к окончаниям, где выводятся из тела клетки.
Некоторые исследователи считают, что в гипоталамусе есть высшие и низшие центры выработки нейросекретов. Высшие центры, связанные с гипоталамусом и корой
95
головного мозга (супрахиазматическое, преоптическое и другие ядра), вырабатывают нейросекрет, который оказывает влияние на выработку нейросекрета низшими центрами (вентромедиальное, аркуатное и др.). Низшие центры индуцируют рилизинг-гормоны, которые стимулируют функцию гипофиза.
На нейросекрецию клетками гипоталамуса оказывает влияние ряд веществ. К ним относят ацетилхолин, се-ротонин, допамин, простагландины, опиаты и другие соединения. Они вырабатываются как в гипоталамусе и гипофизе, так и в центральной нервной системе.
В регуляции тонического выделения Гн-РГ участвуют как адренергические, так и ацетилхолинореактивные структуры, стимулирующие выделение ЛГ из гипофиза. Серотонин оказывает тормозящее влияние на секрецию Гн-РГ.
Стимулирующее влияние на синтез и выделение Гн-РГ оказывают простагландины, особенно Еа. Под влиянием веществ, вырабатываемых в нервной системе, происходит снижение или увеличение синтеза Гн-РГ в клетках гипоталамуса, уменьшение или повышение его выделения из клеток. Одним из эффективных факторов снижения уровня гормона являются пептидазы. В наибольшей концентрации они содержатся в местах образования и действия Гн-РГ и обеспечивают в необходимых случаях быстрое снижение его концентрации.
Несмотря на многочисленные исследования, до настоящего времени нет единых взглядов по локализации в гипоталамусе центров, ответственных за производство отдельных гормонов, или парагипофизарной передаче специфических импульсов. Разные авторы называют противоположные зоны гипоталамуса как ответственные за одну из этих функций. Это возможно связано с тем, что у различных животных расположение зон, регулирующих гонадотропную деятельность гипофиза или передачу нервных импульсов, может быть различной. С другой стороны, трудно вообще представить, что одна из функций гипоталамуса может осуществляться только определенной гипоталамической структурой. Таким образом, маленький гипоталамус является вегетативным центром интеграции многочисленных импульсов экстеро- и интерорецептивной природы; имеет многочисленные связи с центрами коры и ствола головного мозга, с базальными ганглиями, тала-мусом, ретикулярной формацией и со всеми афферентными чувствительными путями; включает около 30 соб-
96
ственных ядер, тесно связанных друг с другом, поэтому понятно, что любая функция гипоталамуса является результатом взаимодействия и совместной деятельности нескольких гипоталамических образований.
Взаимоотношения между гипофизом и половыми железами. В случае удаления из организма семенников у самцов или яичников у самок гонадотропная активность гипофиза существенно повышается. ПриГэтом в передней доле гипофиза появляется большое количество крупных, интенсивно окрашиваемых клеток, содержащих большое количество окрашенных секреторных гранул. Эти клетки были названы клетками кастрации. Также установлено, что введение животным андрогенов или эстрогенов приводит к снижению содержания гонадотропных гормонов в гипофизе.
Кастрация оказывала влияние и на гормональную активность задней доли гипофиза. После кастрации уровень окситоцина в задней доле гипофиза увеличивался в 1,5 раза. Если животным начинали вводить тестостерон, то уровень окситоцина вновь снижался. Инъекциями эстрогенов содержание окситоцина удавалось снизить до уровня более низкого, чем до кастрации. Подобные исследования позволили М. М. Завадовскому (1941 г.) изложить теорию взаимоотношения между гипофизом и половыми железами, выраженную как принцип плюс-минус взаимодействия. Она отражала отношения не только между гипофизом и половыми Железами, но и между гипофизом и всеми остальными эндокринными железами. По этой теории гормоны гипофиза вызывают образование гормонов в семенниках и яичниках (плюс действие). Образовавшиеся половые гормоны тормозят выделение из гипофиза соответствующих гонадотропных гормонов (минус действие). Теория позволяла объяснить данные многочисленных исследований, наблюдавшихся при гипофизэк-томии, кастрации животных и введении им различных гонадотропных и половых гормонов. Так, было известно, что экзогенное введение животным половых гормонов тормозит образование в организме собственных половых гормонов. Механизм действия введенных животным гормонов объясняется следующим образом. Введенные гормоны тормозят образование в гипофизе гонадотропных гормонов. В результате уменьшения продукции гонадотропных гормонов снижается образование в гонадах половых гормонов.
При введении животным гонадотропных гормонов сни-
| 4-232 |
| 97 |
жалась выработка собственных гормонов в гипофизе, объясняемая повышением их уровня в организме. Это приводит к усилению образования в гонадах половых гормонов, что по принципу обратной связи тормозит продукцию в гипофизе гонадотропных гормонов. В дальнейшем была установлена и «пульсация» в выделении гормонов как из гипофиза, так и из половых желез. Это вновь подтверждало теорию М. М. Завадовского о взаимодействии между гипофизом и другими железами внутренней секреции. Оказалось, что пики высокой концентрации гормонов в крови появляются с интервалом в 6 ч, а между ними концентрация гормонов оказывается низкой.
В последующих исследованиях было установлено и взаимодействие между отдельными гормонами гипофиза и половых желез. При введении больших доз тестостерона наблюдали снижение содержания в гипофизе ЛГ. Количество ФСГ при этом не менялось. Это же регистрировали при введении прогестерона самкам. При длительном введении эстрадиола кастрированным крысам в гипофизе уменьшалось содержание как ЛГ, так и ФСГ. Гистологическая картина гипофиза полностью соответствовала его гормональному уровню. Количество и активность клеток, производящих ЛГ и ФСГ, находились в соответствии с количественными показателями этих гормонов в гипофизе.
Исследования последних лет показывают, что ЛГ вызывает образование в семенниках и яичниках андроге-нов, эстрогенов и прогестерона. Все указанные гормоны по принципу обратной связи оказывают тормозящее влияние на образование ЛГ. Саморегуляция системы ФСГ — половые органы осуществляется за счет образования в семенниках и яичниках пептидного гормона ингибина.
Простая схема гормональных взаимоотношений между гипофизом и половыми железами в ряде случаев не выдерживается в связи с тем, что функция гипофиза находится под влиянием гипоталамуса, который не всегда реагирует на концентрацию гормонов половых желез по принципу простой обратной связи. Во многом ответная реакция гипоталамуса на половые гормоны определяется его реактивностью, дозами гормонов. Так, известно, что как низкие, так и чрезмерно высокие дозы эстрогенов тормозят выделение ЛГ из гипофиза. В то же время образуемые во время полового цикла в яичниках в предохотный период сравнительно высокие дозы эстрогенов приводят к резкому усилению выделения из гипофиза ЛГ. Это свидетельствует о наличии в некоторых случаях положительного (плюс)
98
действия половых гормонов на образование гонадотроп-ных гормонов.
Половые гормоны, вводимые в гипоталамус, резко тормозят гонадотропную функцию гипофиза. При введении эстрогенов в передний гипоталамус их тормозящее влияние на гонадотропную функцию гипофиза было выражено в 125 раз сильнее, чем при его введении под кожу. При введении тестостерона в базальную область серого бугра и среднего возвышения гипоталамуса обнаруживалась атрофия яичников и семенников.
В норме у животных устанавливается определенный уровень, на котором функционирует саморегулирующаяся гипоталамо-гипофизарно-гонадальная система. От гормонального уровня этой системы зависит половая функция животных. Она имеет существенные индивидуальные особенности, связанные с генетически фиксированными количественными различиями в уровне активности этой системы. В то же время наблюдения свидетельствуют о том, что и у одного животного уровень саморегуляции между гонадами и гипоталамо-гипофизарным комплексом непостоянен. У молодых животных он сбалансирован на более низком уровне, у взрослых — на более высоком. Помимо генетической обусловленности на уровень этой системы оказывают влияние и факторы внешней среды.

