




ФИЗИОЛОГИЯ
И ПАТОЛОГИЯ
воспроизводства свиней
МОСКВА РОСАГРОПРОМИЗДАТ 1990
преддверия. На нижней стенке преддверия хорошо выражены две парные продольные складки. Они идут от мочеиспускательного отверстия назад. Складки охватывают спереди отверстие мочеиспускательного канала. Преддверие переходит в собственно влагалище. На их границе находится поперечная складка слизистой оболочки, называемая девственной плевой. Девственная плева в виде круговой складки хорошо выражена только у поросят. Длина влагалища колеблется от 13 до 18 см и более. Под слизистой оболочкой проходят два хорошо развитых слоя продольной и кольцевой
|
|

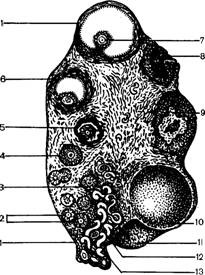
| Рис. 1. Половые органы свиноматки: 1 — тело матки; 2 — рог матки; 3 — яичник; 4 — яйцепровод; 5 — брюшное отверстие яйцепровода; 6 -- отверстие мочеиспускательного канала; 7, 8 — отверстия малых преддверных желез; 9 — головка клитора в ямке; 10 — вентральный угол половых губ; 11 — преддверие влагалища; 12 — средняя и боковые складки слизистой оболочки преддверия; 13 — влагалище; 14 — шейка матки; 15 — воронка яйцепровода |
мускулатуры. Суживаясь, влагалище без заметных границ переходит в шейку матки.
Матка. Длина шейки матки составляет 12—20 см. Внутренняя поверхность ее имеет 17—20 выступов, расположенных в основном с боковых сторон так, что каждый выступ одной стороны входит в пространство между двумя выступами противоположной стороны. В результате образуются так называемые замки шейки матки, среди которых остается спиральный проход. Извилистый ход шейки матки соответствует штопорообразной форме кончика пениса хряка, который получает раздражения, необходимые для нормального проявления совокупительного рефлекса и рефлекса эякуляции. Сужение просвета препятствует у подавляющего большинства свиноматок продвижению пениса или катетера через всю длину шейки матки, что имеет важное физиологическое значение (предотвращает попадание спермы только в один из рогов матки в случае продвижения пениса или катетера через шейку и тело матки).
Мускулатура шейки матки развита сильно. Шейка переходит в небольшое тело матки длиной 5—10 см. Мышечный слой ее тела меньше развит, чем в шейке. От тела в стороны отходят два рога матки и располагаются в брюшной полости в виде петель, похожих на петли тонкого отдела кишечника. Длина их у взрослых маток достигает 1 м, а масса обоих рогов — 600—800 г. Во время супо-росности длина и масса рогов матки значительно возрастают. Длина составляет 250 см и более, масса — 7 кг. Слизистая рогов формирует глубокие извилистые складки с многочисленными железками, секрет которых (маточное молочко) обеспечивает питание зародышей на ранних стадиях их развития. Трубчатые маточные железки имеются во всей толще слизистой оболочки. В глубокой ее части они расположены гуще. Диаметр их возрастает к отверстию железы. Помимо трубчатых желез в эпителии слизистой матки присутствуют железистые клетки, вырабатывающие слизисто-серозный секрет. В вершине каждого рога матки находится очень узкое отверстие, которое ведет в яйцепровод. На границе яйцепровода и рогов матки расположено маточно-трубное сочленение. Оно включает полиповидный отросток на слизистой матки и отверстие яйцепровода, выполняющее функцию сфинктера (рис. 2).
Маточно-трубное сочленение служит депо для сперми-ев, регулирует поступление спермиев в яйцепровод и переход яйцеклеток в матку. При удалении маточно-труб-ного сочленения количество спермиев, поступающих в яйцепровод, значительно возрастало, что приводило к полиспермии при оплодотворении яйцеклеток. В указанном сочленении отмечена наиболее высокая переживаемость спермиев. Возможно, что маточно-трубное сочленение осуществляет не только количественный, но и качественный контроль за спермиями, поступающими в яйцепровод. Так, при осеменении маток размороженной спермой оплодо-
|
|

| Рис. 2. Микрофотография сканирования маточно-трубного сочленения |
творения яйцеклеток не наступало, а при введении спермы хирургическим путем непосредственно в яйцепровод свиноматки были оплодотворены.
Яйцепровод. Яйцепровод имеет длину 20—25 см. Наружный диаметр его у маточного конца (истмуса) составляет 2—3 мм. В средней части (ампуле) он расширяется и достигает 6—8 мм, заканчиваясь отверстием, которое называется брюшным. Отсюда яйцепровод начинает резко расширяться в сторону яичника, в результате чего приобретает форму воронки. Ее конечная часть достигает 9 см в диаметре. Воронка охватывает яичник. Во время овуляции в результате отечности яичника и яйцепро-вода этот охват становится плотным, что препятствует попаданию яйцеклеток в брюшную полость. Наружный край воронки неровный и называется бахромкой яйцепровода.
Слизистая средней части яйцепровода имеет сильную многоступенчатую складчатость. В первичных складках проходят кровеносные и лимфатические сосуды, а также нервы, которые отдают веточки во вторичные, третичные и четвертичные складки. Складчатость значительно уменьшается в истмической части яйцепровода и полностью исчезает в воронке. Слизистая яйцепровода покрыта однослойным цилиндрическим эпителием из мерцательных ц секреторных клеток. В период полового цикла соотношение обоих типов клеток меняется. Во время эст-руса преобладают секреторные клетки, в остальные периоды — мерцательные.
Мышечная оболочка состоит из кольцевого слоя глад-комышечиых и небольшого количества косых мышечных клеток. К серозной оболочке примыкает слой продольной мускулатуры.
В яйцепроводах осуществляются процессы передвижения яйцеклеток к рогам матки, а спермиев — в обратном направлении. Ранее предполагали, что сокращение мускулатуры яйцепроводов и движения ресничек реснитчатого эпителия сориентированы таким образом, чтобы регулировать передвижение секретов и яйцеклетки в сторону рогов матки. Спермии же за счет так называемого явления реотаксиса (способности двигаться против слабого тока жидкости) продвигаются по яйцепроводу от его маточного конца по направлению к яичникам.
В последние годы исследователи пришли к выводу, что спермии продвигаются через яйцепроводы за счет сокращения их мышечных стенок. Иначе невозможно объяснить чрезвычайно быстрое продвижение спермиев через
яйцепровод. Так, если учитывать, что длина яйцепро-вода в среднем 20 см, а скорость движения спермиев составляет около 0,5 см в 1 мин, то, очевидно, спермии аа очст <-,обс,те<нж<эт'о дтажениА могут п.реодол.еть я_йце-провод примерно за 40 мин. Однако в 50-х годах было установлено, что у коров спермии попадают в яичниковые участки яйцепроводов уже через несколько минут после осеменения. Это происходило и в тех случаях, когда использовали сперму с погибшими спермиями. Подобные данные в дальнейшем получили и на свиноматках. При осеменении мертвыми и живыми спермиями сперма начинала выделяться из трех яйцепроводных канюль в начальной, средней и задней частях яйцепровода через 4,5 мин после осеменения. При изоляции яйцепроводов через 15 мин после осеменения свиноматок в них затем находили оплодотворенные яйцеклетки. В некоторых исследованиях спермии обнаруживали в яйцепроводах при убое свиноматок сразу после естественного осеменения.
В опытах с фистулами яйцепроводов наблюдали чередование выделения секрета из его яйцепроводного и маточного конца, что указывает на движение его в обоих направлениях. Это свидетельствует о том, что основную роль в движении секрета играют сокращения мышечных стенок яйцепровода.
В ряде исследований установлено, что все участки яйцепровода имеют спонтанную активность. В первый день цикла почти во всех его участках отмечали антиперистальтические сокращения. В предовуляторпый период число перистальтических и антиперистальтических сокращений примерно было одинаковым. Затем в течение 3 дней антиперистальтические движения отсутствовали. На 4-й день цикла, когда эмбрионы переходят в матку, в ампуло-истмусовом и истмусовом участках выявляли сильные перистальтические сокращения. В лютеальную фазу уровень сокращений был низким.
Появление спермиев в яйцепроводах вскоре после осеменения свидетельствует о том, что они быстро продвигаются через рога матки, достигающие у свиней длины 1—2 м. Естественно, это может происходить только за счет пассивного перемещения спермы через рога, благодаря их сильным антиперистальтическим (засасывающим) сокращениям.
Определены значительные колебания давления в полости матки и изменение потенциалов ее мускулатуры, особенно в период половой охоты. Исследования с по-
яйцепровод. Так, если учитывать, что длина яйцепро-вода в среднем 20 см, а скорость движения спермиев составляет около 0,5 см в 1 мин, то, очевидно, спермии за счет собственного движения могут преодолеть яйцепровод примерно за 40 мин. Однако в 50-х годах было установлено, что у коров спермии попадают в яичниковые участки яйцепроводов уже через несколько минут после осеменения. Это происходило и в тех случаях, когда использовали сперму с погибшими спермиями. Подобные данные в дальнейшем получили и на свиноматках. При осеменении мертвыми и живыми спермиями сперма начинала выделяться из трех яйцепроводных канюль в начальной, средней и задней частях яйцепровода через 4,5 мин после осеменения. При изоляции яйцепроводов через 15 мин после осеменения свиноматок в них затем находили оплодотворенные яйцеклетки. В некоторых исследованиях спермии обнаруживали в яйцепроводах при убое свиноматок сразу после естественного осеменения.
В опытах с фистулами яйцепроводов наблюдали чередование выделения секрета из его яйцепроводного и маточного конца, что указывает на движение его в обоих направлениях. Это свидетельствует о том, что основ ную роль в движении секрета играют сокращения мышечных стенок яйцепровода.
В ряде исследований установлено, что все участки яйцепровода имеют спонтанную активность. В первый день цикла почти во всех его участках отмечали антиперистальтические сокращения. В предовуляторный период число перистальтических и-антиперистальтических сокращений примерно было одинаковым. Затем в течение 3 дней антиперистальтические движения отсутствовали. На 4-й день цикла, когда эмбрионы переходят в матку, в ампуло-истмусовом и истмусовом участках выявляли сильные перистальтические сокращения. В лютеальную фазу уровень сокращений был низким.
Появление спермиев в яйцепроводах вскоре после осеменения свидетельствует о том, что они быстро продвигаются через рога матки, достигающие у свиней длины 1—2 м. Естественно, это может происходить только за счет пассивного перемещения спермы через рога, благодаря их сильным антиперистальтическим (засасывающим) сокращениям.
Определены значительные колебания давления в полости матки и изменение потенциалов ее мускулатуры, особенно в период половой охоты. Исследования с по-
мощью фистул показали, что рога матки сокращаются неодновременно, и сперма при осеменении попадает сначала в один, а затем в другой рог. Засасывающие движения рогов чередуются с выталкивающими (перистальтическими). Сперма вытекала из фистул рогов периодически через 1—3 мин обычно в течение 30 с. В результате асимметричного сокращения рогов матки в каждый рог во время естественного и искусственного осеменения попадает неодинаковое количество спермы.
Данные по скорости продвижения спермиев через рога матки чрезвычайно противоречивы. Спермии находили по всей длине матки при убое свиноматок сразу после их покрытия, их обнаруживали в верхушках рогов через 8 мин после осеменения, через 20 и 40 мин. Эти различия могут быть объяснены качеством проведения операции по наложению фистул, различным периодом половой охоты, в который проводилось осеменение, но в основном индивидуальными особенностями животных. При осеменении свиноматок мы наблюдали, что у некоторых животных до 30—40 мл.спермы засасывается в половые органы в течение 20—30 с после начала осеменения, а у других никаких засасывающих движений не происходит и в течение 20—25 мин после введения катетера в шейку матки. Как частота и продолжительность, так и сила засасывающих движений различны у отдельных животных. Изучение силы сокращений и их направления в рогах матки в различные периоды полового цикла с помощью тонометров показали, что сила сокращения рогов матки возрастает в предохотный период и достигает максимальных значений в период охоты. В послеохотный период, особенно в период полового покоя, уровень сокращений низкий. Волны сокращений на 2-й день охоты почти исключительно антиперистальтические, а по мере удаления от этого пункта как в сторону предохотного, так и в сторону послеохотного периода количество антиперистальтических волн сокращается, а число перистальтических — возрастает.
Исследования скорости поступления спермиев после осеменения в вершины яйцепроводов противоречат этим данным. Как при осеменении на 2-й день охоты, так и при осеменении на 7-й день после нее скорость поступления спермиев, их число и подвижность через 15 мин и 24 ч после осеменения оказались одинаковыми. Подобные результаты получены при осеменении свиноматок и в другие периоды охоты и лютеальной фазы цикла.
Сократительная функция рогов матки обеспечивает равномерное распределение в них поступающих из яйце-провода оплодотворенных яйцеклеток и выведение зрелых плодов во время родов. В рогах матки происходит питание зародышей вначале за счет маточного молочка, а в дальнейшем за счет плацентарного кровообращения. Образующиеся в них простагландины обусловливают лютеолиз желтого тела. Через тело матки зародыши перемещаются во время их миграции из одного рога в другой.
Сильная мускулатура шейки матки обеспечивает выведение плодов во время родов. Сокращение мускулатуры шейки матки во время охоты не совпадает с сокращениями мускулатуры рогов матки.
Трубчатые половые пути выстланы изнутри слизистой. Ее окружает мышечный слой, который внутри состоит из кольцевых, а снаружи из продольных мышечных волокон. Покрыты половые органы серозной оболочкой.
|
|
Яичники. Яичники (рис. 3) — парные органы. В них образуются половые клетки и гормоны, необходимые для осуществления процессов воспроизводства. У неполовозрастных свинок яичники имеют бобовидную форму и гладкую поверхность. Незадолго до и после наступления половой зрелости в результате выпячивания над поверхностью фолликулов и желтых тел яичники приобретают форму ежевики или тутовой ягоды. Их размеры и масса зависят в большей степени от стадии полового цикла, возраста и массы животных. Длина яичника колеблется от
| Рис. 3. Схема строения яичника свиньи: 1 — зачатковый эпителий; 2 — овоциты I порядка; 3 — первичный фолликул; 4 — растущий (вторичный) фолликул; 5 — появление полости в фолликуле; 6 — пузырчатый фолликул; 7 — яйцеклетка в зрелом фолликуле; 8 — недавно овулировавший фолликул, наполненный кровью (красное тело); 9 — м&лодое желтое тело; 10 — сформировавшееся желтое тело; 11 — атретический фолликул; 12 — сосуды мозгового слоя яичника; 13 — брыжейка |
10
2 до 7 см, ширина — от 1,5 до 4 см, а масса — от 2 до 15 г. У 8—9-месячных свинок средняя масса яичников в первой половине супоросности составляет 4,1 г, а в возрасте 3—4 лет (масса свиноматки — 250—280 кг) — 11,6 г. У половозрелых свиноматок с живой массой 100— 150 кг масса яичников на стадии лопнувших фолликулов достигает 3,9 г (с колебаниями от 2,1 до 5,9 г), в возрасте 1—2,5 года — 6,3 г (с колебаниями от 4,3 до 12,7 г).
Яичники имеют плотную эластичную консистенцию. Цвет яичников зависит от состояния половой функции животного. У неполовозрелых свинок они белые, перед овуляцией приобретают розово-красный, а затем кирпично-красный цвет. Во время супоросности они становятся фиолетовыми. Существуют различия в цвете обескровленных яичников (после убоя животного) и находящихся в физиологическом состоянии (при лапаротомии). В последнем случае они имеют более интенсивную светло-красную окраску.
Яичники покрыты зародышевыми клетками кубического эпителия. Под слоем зародышевого эпителия находится соединительнотканная белочная оболочка. Она покрывает основу яичника, состоящую из рыхлой соединительной ткани. В ней различают наружную (корковую) и внутреннюю (мозговую) зоны. Мозговая зона состоит из рыхлой, частично ретикулярной, соединительной ткани, а соединительная ткань корковой зоны содержит тонкие пучки коллагеновых волокон и фиброциты. В соединительной ткани яичника как в корковой, так и в мозговой зоне имеются интерстициальные клетки. Они мезенхимного происхождения и участвуют как в эндокринной функции, вырабатывая эстрогены, так и в трофической. В корковом слое расположено множество фолликулов, находящихся на разной стадии развития или атрезии (гибели), и желтых тел. В мозговой зоне находится большое количество нервных волокон, крупных кровеносных и лимфатических сосудов. В центре зоны они многократно ветвятся. Сосуды штопорообразно извиваются. Вены образуют венозное сплетение. Отсюда нервы и сосуды направляются в корковый слой, где достигают всех функциональных образований. Вокруг фолликулов, желтых тел, атретических фолликулов и интерстициаль-ных клеток формируются нервные сплетения. Нервы достигают яйцеклетки. В зависимости от стадии развития фолликулы располагаются в различных участках коркового слоя. Они состоят из половой и фолликулярных кле-
11
ток, последние выполняют по отношению к половой клетке трофическую (питательную) роль.

