




У неполовозрелых самок самые мелкие фолликулы (первичные, примордиальные) располагаются в корковом слое у его поверхности. Их количество у новорожденных свинок достигает сотни тысяч. Каждый из них состоит из половой клетки (овоцита первого порядка), окружающего ее одного слоя фолликулярных клеток и соединительнотканной оболочки—теки (рис. 4). Периодически часть первичных фолликулов начинает расти. По мере развития они перемещаются в корковом слое ближе к центру яичника. Во время развития эпителий фолликула из плоского становится сначала кубическим, а затем призматическим. Размножаясь, фолликулярные клетки окружают овоцит первого порядка сначала в два, а затем и в несколько слоев. Формируются так называемые вторичные фолликулы, которые еще больше углубляются в корковый слой.
|
|
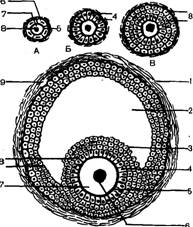
В дальнейшем в центре фолликула появляется наполненная жидкостью полость, образующаяся из слившихся небольших, наполненных жидкостью полостей между фолликулярными клетками. Фолликул превращается в пузырек (пузырчатый фолликул, Гра-афов пузырек).
В процессе развития фолликула происходит рост овоцитов. Они покрываются блестящей (прозрачной) оболоч-
| Р и с. 4. Строение фолликула: А — Б — первичный фолликул; В — вторичный фолликул; Г — пузырчатый фолликул; ] — соединительная оболочка пузырчатого фолликула; 2 — полость пузырчатого фолликула; 3 — яйценосный бугорок; 4 — лучистый венец; 5 — блестящая зока овоцита 1-го порядка; в — ядро овоцита 1-го порядка; 7 — овоцит 1-го порядка; 8 — фолликулярные клетки; 9 — кровеносный сосуд |
кои, которая состоит из мукополисахаридов и является продуктом обмена веществ овоцита. Вокруг овоцита фол-
12
ликулярный эпителии располагается в виде правильных радиальных лучей (лучистый венчик). Эти клетки выполняют по отношению к овоциту трофическую (питательную) роль. Их длинные отростки проникают через отверстия в прозрачной оболочке к ее цитоплазме. Остальной фолликулярный эпителий фолликула называется зернистым.
По мере роста пузырчатый фолликул занимает всю толщу коркового слоя и выпячивается над поверхностью яичника в виде прозрачного пузырька. Диаметр зрелого фолликула достигает 10—12 мм. Развитый фолликул представляет собой пузырек, центр которого заполнен жидкостью. Полость окружена зоной фолликулярных (гранулезных, зернистых) клеток, располагающихся в 10—12 слоев. В участке, где находится яйцеклетка, фолликулярные клетки образуют утолщение, которое вдается в полость фолликула в виде бугорка. На его вершине расположена окруженная лучистым венцом яйцеклетка. Снаружи фолликул покрыт соединительнотканной оболочкой, образованной из стромы яичника. В ней различают два слоя: внутренний, прилегающий к клеткам грану-лезы, и наружный — к строме яичника. Во внутреннем с;юе имеется много мелких кровеносных сосудов и клеток. Он отделен от клеток гранулезы нежной соединительнотканной мембраной. Клетки внутренней оболочки в строении сходны с клетками желтого тела, поэтому их иногда называют текалютеиновыми. Морфологически они очень изменчивы и участвуют в обмене веществ как в фолликуле, так и в желтом теле. Клетки определяют движение фолликула к поверхности яичника, их созревание и овуляцию, а также образование, функцию и регрессию желтого тела. Они находятся в тесной связи с клетками гранулезы.
Клетки внутреннего слоя служат основным местом выработки прогестерона, андрогенов и частично эстрогенов. Наружный слой оболочки образован пластинчатыми пучками и клетками. Он беден кровеносными сосудами. В том месте, где фолликул выступает над поверхностью яичника, он покрыт снаружи также белочной оболочкой яичника и слоем зародышевого эпителия.
Из растущих фолликулов только часть полностью развивается. Большинство их погибает на разных стадиях развития. Половые, а частично и фолликулярные клетки в таких фолликулах разрушаются, а оболочки и интерс-тициальные клетки достигают сильного развития.
13
Атретические фолликулы имеют вид мутновато-белых точечных структур. Учитывая, что атрезия фолликулов происходит постоянно, в последнее время большинство исследователей приходит к выводу, что это явление физиологическое, необходимое для нормального течения циклических процессов в яичниках. Возможно, что атрезия — следствие тонкого механизма, реагирующего на изменения в организме и обеспечивающего созревание и овуляцию оптимального количества фолликулов. При заболеваниях животных процесс атрезии фолликулов усиливается. Считается, что первичные изменения в атре-тичных фолликулах происходят в ооцитах, которые в дальнейшем вызывают перестройку в обмене веществ в клетках гранулезы. В клетках гранулезы снижается синтез эстрогенов из тестостерона, и в фолликулах образуется повышенное количество андрогенов.
Яичники неполовозрелых свинок содержат различное количество фолликулов диаметром 1—4 мм. Перед половым созреванием многие фолликулы увеличиваются до 3—7 мм в диаметре. Часть из них располагается в глубокой, но большинство — в поверхностной зоне коркового слоя. Их количество в одном яичнике достигает 50. Незадолго до полового созревания число таких фолликулов снижается до 5—6 в одном яичнике. У взрослых животных в период охоты происходит быстрый рост обычно 10—20 фолликулов, достигающих максимальной величины. После созревания все или большая часть созревших фолликулов лопается и находящаяся в них яйцеклетка вместе с фолликулярной жидкостью попадает в воронку яйцепровода. Процесс выделения яйцеклетки из фолликула называется овуляцией.
Причины, вызывающие овуляцию фолликулов, еще недостаточно раскрыты. Длительно существовавшая гипотеза о том, что разрыв фолликула вызывается давлением увеличивающегося в нем количества жидкости, не подтвердилась. Установлено, что перед овуляцией давление жидкости в фолликуле не только не возрастает, но даже уменьшается. Так, у коров при пальпации отмечается снижение упругости фолликула, а у собак перед овуляцией на стенке фолликула даже появляется складка. В фолликулярных кистах, несмотря на большое количество жидкости и более высокое давление, чем в фолликулах, разрыва стенок не происходит. Не удавалось вы-зйать овуляцию и введением в зрелые фолликулы дополнительного количества жидкости. Не подтвердилась
14
также гипотеза о том, что овуляцию вызывают сокращения гладкомышечных элементов яичника или стенок фолликула. В опытах было установлено, что овуляция происходит и в отделенных от яичника фолликулах, а в стенках фолликула не найдены гладкомышечные волокна. Овуляция отделенных от яичника фолликулов свидетельствует также об отсутствии связи ее с нервными импульсами.
В настоящее время считается, что овуляция связана с определенными биохимическими и ферментативными процессами в стенке фолликула. При созревании фолликула в лишенном кровеносных сосудов участке его стенки, где в дальнейшем происходит ее разрыв, она начинает истончаться. Сокращается и прилегающий к этому участку слой зернистых клеток. В результате истончения стенки в этом месте над поверхностью фолликула образуется круглое выпячивание, которое затем разрывается. С помощью лапаротомии и киносъемки удалось установить, что во время овуляции яйцеклетка с окружающими ее клетками медленно вытекает из фолликула с фолликулярной жидкостью. В момент овуляции фолликулярная жидкость имеет высокую ^вязкость, которая связана с со-.-р^лшимп серу мукополисахаридами. Значительная вязкость жидкости и медленное ее вытекание предотвращают попадание яйцеклетки в брюшную полость.
Возрастание в фолликулярной жидкости мукополиса-харндов связывается с деполимеризацией, вызванной, в свою очередь, появлением в ней гиалуронидазы. При -- 1м увеличивается онкотическое давление, вызывая приток в фолликул жидкости из окружающей среды. Синтез гиалуронидазы происходит под влиянием люте-инизирующего гормона (ЛГ). Перед овуляцией в фолликуле возрастает количество протеолитических ферментов, которые, вероятно, принимают существенное участие в лизисе оболочки фолликула. Так как у некоторых видов животных овуляция вызывается половым актом, считалось, что ее пусковым механизмом являются нервные импульсы. Однако в дальнейшем было установлено, что овуляцию вызывает ЛГ. Уже через 1 ч после полового акта у кролика выделяется ЛГ из передней доли гипофиза. Введение крольчихам ЛГ обусловливает овуляцию и без полового акта. У животных со спонтанной овуляцией выброс ЛГ из гипофиза отмечают в определенные периоды полового цикла. У свиней пик выделения ЛГ наблюдается незадолго до охоты или совпадает с ее началом. Овуляция
15
осуществляется примерно через 40 ч после этого. Процесс овуляции всех фолликулов длится около 2 ч. После разрыва оболочки грунулеза остается неповрежденной, за исключением яйценосного бугорка.
В исследованиях на фолликулах кроликов, у которых овуляция происходит через 11—12 ч после введения хорионического гонадотропина (ХГ), удалось установить, что наиболее ранней реакцией на ХГ является гиперемия яичников. При этом расширяются как кровеносные, так и лимфатические сосуды. Изменяются стенки сосудов, и клеточные элементы крови выходят за их пределы. Через 6 ч под базальной мембраной появляются свободно лежащие лейкоциты, а через 8 ч и эритроциты. Сосудистые изменения приводят к отеку ткани вокруг фолликулов. Размеры фолликулов увеличиваются. В течение 6 ч перед овуляцией их величина возрастает более чем в 2 раза. К моменту овуляции в месте разрыва полностью исчезает фолликулярный эпителий, который имел до этого 8— 12 слоев. Яйценосный бугорок удлиняется и истончается. Фолликулярный эпителий разрыхляется. Лучистый венец в первые часы после введения ХГ имеет 3—4 слоя. Через 4 ч здесь отмечают признаки отека, а через 8ч — дегенерацию клеток. Непосредственно перед разрывом фолликула яйцеклетка окружена одним слоем клеток лучистого венца. За 1—2 ч до разрыва фолликула яйценосный бугорок разрушается и яйцеклетка свободно плавает в фолликулярной жидкости. Через 4—6 ч после введения ХГ в результате отека соединительнотканная оболочка отделяется от фолликулярного эпителия. Через 8—10 ч в текальной и белочной оболочках наступает диссоциация клеток, которые оказываются плавающими в отечной жидкости. В месте будущего разрыва фолликула происходит дегенерация клеток, и его оболочка истончается. За 30— 60 мин до разрыва фолликула соединительнотканная оболочка в участке его разрыва практически отсутствует. По мере приближения овуляции в месте разрыва фолликула возрастает гибель всех слоев стенки. Перед разрывом фолликула в участке разрыва наблюдается только один слой фолликулярного эпителия, а иногда и клетки поверхностного эпителия. Участки теки и белочной оболочки фолликула отсутствуют. После разрыва стенки из фолликула медленно вытекает фолликулярная жидкость, а через 3—30 мин выделяется и яйцеклетка. Учитывая, что у свиней овуляция происходит примерно через 40 ч после выделения ЛГ, то все эти процессы протекают более медленно.
16
Образование и развитие желтого тела. После выделения яйцеклетки с окружающими ее клетками яйценосного бугорка полость фолликулов заполняется кровью из разорвавшихся сосудов. Клетки зернистого и внутреннего слоев оболочки фолликула начинают размножаться и постепенно замещают кровяной сгусток. В него проникают со стороны оболочки соединительная ткань и кровеносные сосуды. Они образуют радиальные перегородки, разделяющие желтое тело на дольки. Сосуды разветвляются, проникают внутрь дольки и окружают клетки. После овуляции клетки гранулезы гипертрофируются и превращаются в липидосодержащие лютеиновые клетки желтого тела. На 7-й день они увеличиваются в размерах и достигают 30—40 мкм в диаметре. Клетки желтого тела, образующиеся из клеток внутренней оболочки, имеют меньшие размеры (10—25 мкм) и содержат вакуоли и капельки жира.

