




У всех млекопитающих половая дифференцировка мужского индивидуума происходит хронологически раньше, чем женского. Развитие семенников определяет мужская У-хромосома. Это происходит даже тогда, когда в некоторых патологических случаях одной У-хромосоме сопутствуют две женских Х-хромосомы. Развитие яичников связано с двумя Х-хромосомами. При наличии только одной Х-хромосомы гонады остаются недифференцированными. Дальнейшее половое развитие в мужском организме определяется гормонами, вырабатываемыми в семенниках зародыша. При отсутствии гормонов, вне зависимости от генетической обусловленности, развитие идет по женскому типу.
Дифференциация мужских гонад начинается с образования семенных канальцев, в которых зародышевые клетки окружены клетками Сертоли. Мейоз наступает ко времени половой зрелости. Семенные канальцы появляются у плодов в возрасте 28 дней. В 30-дневном возрасте
4** 99
в семенниках образуются клетки Лейдига, в которых начинает вырабатываться стероидный гормон. В этот период уровень тестостерона в мужских плодах существенно повышается. Секреция гормона достигает максимума у плодов 35-дневного возраста. В то время как морфологическое дифференцирование клеток Лейдига не зависит от гонадо-тропных гормонов, продукция стероидных гормонов семенника связана с функцией фетального гипофиза. В женском зародыше задолго до образования структур яичника в 35 дней происходит значительное увеличение числа зародышевых клеток. Клетки вступают в мейотическую профазу прежде, чем они окружаются фолликулярными клетками, которые, вероятно, тормозят дальнейший мейоз. Только после этого начинается развитие яичника, в котором выделяются корковая и мозговая зоны. Ко времени недифференцированных гонад имеются парные Вольфов и Мюллеров каналы. У мужских организмов из Вольфова канала образуются придатки, спермиопровод и пузырьковидные железы. Мюллеров канал редуцируется. У женских плодов регрессирует Вольфов канал, а из Мюллерова канала формируются яйцепроводы и матка.
Установлено, что фетальные семенники вырабатывают две различные субстанции, из которых одна определяет регрессию Мюллерова канала, а другая ответственна за дифференциацию Вольфова канала. Ингибитор Мюллерова канала не является мужским половым гормоном, в то время как дифференциацию Вольфова канала определяют андрогены. Если в фазу половой дифференцировки в генетически женских плодах появляются существенные количества андрогенов, то образование яичников и преобразование Мюллерова канала в матку и яйцепроводы не нарушаются. В то же время из развивающегося Вольфова канала образуются придатки семенников, семяпроводы и пузырьковидные железы. Образование влагалища тормозится, и развитие наружных половых органов идет по мужскому типу, т. е. образуется пенис. Подобный половой тракт наблюдали у генетически женских индивидуумов в эксперименте, когда на 30-й день после случки в плоды вводили тестостерон-пропионат. Андрогены оказывают существенное влияние на половую дифференцировку только в определенный период развития плодов. Введение же тестостерона на 40-й день жизни приводит только к маскулинизации женских плодов, что выражается в переразвитии клитора. Половая дифференцировка сказывается не только на анатомии и морфологии гонад и развитии Воль-
100
фова и Мюллерова каналов, но и на деятельности гипо-таламо-гипофизарной системы и полового поведения. Известно, что циклическая деятельность половой системы самок определяется циклическим выделением гонадотроп-ных гормонов из гипофиза. У самцов эта цикличность отсутствует и выделение гонадотропных гормонов из гипофиза, а половых из семенника происходит постоянно.
Исследованиями установлено, что цикличность выделения половых гормонов у самок зависит не от гипофиза, а от гипоталамуса. Так, если пересадить гипофиз самца гипофизэктомированной самке, то половая цикличность у нее сохраняется. Если пересадить яичник кастрированному самцу, то в яичнике развиваются и постоянно присутствуют крупные фолликулы, которые однако не овули-руют. У самца при этом происходит разрастание молочных желез, характерное для лактирующей самки, что связано с постоянной продукцией эстрогенов. При удалении гипофиза и семенников у самцов и пересадке им гипофиза и яичников самки циклическая секреция гонадо-тропинов отсутствовала.
Половая дифференцировка гипоталамуса происходит после рождения, и проявляется она у самцов. Дифференцировка гипоталамуса по мужскому типу происходит у самцов в первые дни после рождения. Ответственными за дифференцировку являются вырабатываемые в семенниках андрогены. Если в первые дни после рождения в организме отсутствуют достаточные количества андрогенов, то дифференцировка гипоталамуса происходит по женскому типу, что проявляется в циклическом выделении гипофизом гонадотропных гормонов.
Эндокринология полового развития и созревания. Половое созревание — фаза развития организма, при которой он достигает способности к выделению зрелых половых клеток. У свиноматок это происходит при первой половой охоте, сопровождающейся овуляцией. Возраст наступления половой зрелости имеет существенные индивидуальные особенности и составляет у свиноматок 4—9, у хрячков 4—6 месяцев.
Примордиальные фолликулы появляются в яичнике уже в эмбриональный период. На протяжении жизни часть примордиальных фолликулов вступает в дальнейшее развитие. Факторы, вызывающие этот процесс, остаются неизвестными. Развитие фолликула до стадии пузырчатого, вероятно, не зависит от гонадотропных гормонов.
101
И только в дальнейшем для их развития необходимы гонадотропины. Уже в конце внутриутробного периода гонадотропин-рилизинг-гормон гипоталамуса стимулирует секрецию гипофизом ЛГ и ФСГ, а они, в свою очередь-развитие гонад. В последний месяц плода и первый после рождения уровень ЛГ и ФСГ в крови высокий. Затем уровень ЛГ снижается, а ФСГ остается высоким и увеличивается к 2-месячному возрасту. К 3-му месяцу ритм колебаний уровня ЛГ становится более выраженным, что приводит к усилению развития яичников и фолликулов. Усиление секреции эстрогенов приводит к снижению уровня ЛГ и ФСГ в крови у свинок в возрасте около 4 месяцев. Возможность вызывания стрессов полового созревания в этом возрасте у многих свинок свидетельствует о том, что гормональная система готова к запуску полового цикла задолго до появления первой охоты.
У неполовозрелых хряков в семенниках вырабатываются в основном тестостерон и андростенон. На 2-й неделе жизни уровень гормонов в семенниках достигает высших значений. В 1 г семенника содержится 962 нг андростенона и 1,7 нг тестостерона. До 63-го дня уровень андрогенов в семенниках постоянно снижается, а с 70-го дня постоянно возрастает до 280-го дня за счет резкого повышения продукции тестостерона. В 2-месячном возрасте уровень андростенона в семенной вене составляет 170 нг/мл, тестостерона — 8 нг/мл. В 4-месячном возрасте эти показатели равняются 90 и 70 нг/мл, а в 6-месячном возрасте — 60 и 140 нг/мл. Вероятно, до полового созревания тормозятся отдельные реакции биосинтеза тестостерона. Концентрация прогестерона в плазме крови хряков возрастает с 1,11 нг/мл в 65-дневном возрасте до 2,24 нг/мл в 150-дневном возрасте и снижается к возрасту в 178 дней до 1,99 нг/мл.
Эндокринологически половое созревание связано с усиленным образованием у хрячков тестостерона, а у свинок — эстрогенов, которые приводят к полному развитию половых органов и вторичных половых признаков. Гормональный механизм полового созревания недостаточно ясен. При половом созревании у свиней не установлено повышенного содержания в плазме крови гонадотропных гормонов, которые ответственны за образование половых гормонов в гонадах. Увеличение уровня тестостерона при половом созревании объясняется повышением в этот период реакции семенников на гипофизарный ЛГ. Это связывается с деятельностью центральной нервной системы и
102
гипоталамуса. Отмечено также повышение амплитуды пульсации выделения ЛГ гипофизом. В опытах не было установлено разницы в выделении ЛГ гипофизом в ответ на введение гонадотропин-рилизинг-гормона новорожденным, достигшим половой зрелости, и взрослым хрякам. В то же время при этом отмечена четкая возрастная реакция семенников на введение рилизинг-гормона и вызванное в результате этого увеличение секреции ЛГ гипофизом. У новорожденных хрячков это не приводило к увеличению секреции тестостерона, а у половозрелых и взрослых животных происходил четкий подъем уровня тестостерона в плазме крови. Другим фактором повышения уровня тестостерона у половозрелых животных является изменение его обмена в организме. С возрастом животных уменьшается степень его разрушения.
Подобный механизм полового созревания наблюдался и у самок. Ввиду повышения чувствительности яичников к ЛГ, а возможно, и в связи с изменением амплитуды пульсации ЛГ в яичниках происходит рост фолликулов и повышается уровень образующихся эстрогенов. В результате уменьшения степени их распада концентрация эстрогенов достигает уровня, необходимого для появления признаков охоты и преовуляторного выделения ЛГ гипофизом. Однако перед этим гипоталамус начинает реагировать на эстрогены по принципу обратной связи. Такая реакция отсутствует у новорожденных свинок. У свинок в возрасте 80—100 дней в яичниках имеются терциальные фолликулы. При инъекции гонадотропинов свинкам отмечаются созревание и овуляция фолликулов. Однако образующиеся желтые тела даже в случае супоросности преждевременно регрессируют, поэтому получить опорос от свинок удается только в более позднем возрасте. Предпосылкой этого, вероятно, служит тот факт, что нормальная секреторная активность эндометрия в количественном отношении достигается у свинок примерно в 5-месячном возрасте, а в качественном — несколько позднее.
В близкий к половозрелости период у свинок часто проявляются признаки охоты, однако она неполноценная и не сопровождается овуляцией. Это свидетельствует о том, что до наступления половой зрелости у маток несколько раз могут созревать группы фолликулов. Образующегося количества эстрогенов оказывается достаточно для наступления охоты, но недостаточно для того, чтобы вызвать предовуляторное выделение ЛГ из гипофиза. В результате фолликулы не достигают полного созревания.
103
Частично, вероятно, это также связано с еще недостаточной ответной реакцией гипоталамуса на эстрогены.
В гипофизе задолго до овуляции содержится достаточное для запуска полового цикла количество гонадотроп-ных гормонов. Однако до полового созревания они не поступают в кровь. Точный механизм пониженной чувствительности гипоталамуса к половым гормонам, а гонад к гонадотропным гормонам до полового созревания остается неясным. В последнее время большое внимание в этом отношении уделяют эпифизу, который оказывает на гипоталамус и половые органы указанное влияние.
Эндокринология полового цикла. В раннюю фолликулярную фазу (за 4—2 дня до охоты или 16—18-е дни цикла) отмечается быстрый рост группы фолликулов. Их размеры возрастают от 4 до 10—12 мм. Средний уровень ФСГ и ЛГ в этот период мало изменяется (рис. 12). Однако отмечается повышение частоты пульсирующего выделения ЛГ, снижение уровня прогестерона, который оказывает как прямое блокирующее влияние на развитие фолликула, так и непрямое за счет торможения выделения ЛГ.
Установлено, что не все крупные фолликулы достигают овуляторной фазы. Развивающиеся крупные фолликулы могут претерпеть обратное развитие, а овуляцион-ной стадии достигают более мелкие фолликулы, которые вырастают до крупных зрелых фолликулов менее чем за 2 суток.
После регрессии желтого тела в яичниках имеются фолликулы, часть из которых является эстрогенно активными, а часть — неактивными. Последние атрезируют-ся, и к моменту преовуляторного пика ЛГ в яичнике остаются только эстрогенно активные фолликулы. Концентрация эстрадиола в них превышает 1 мкг/мл. После преовуляторного выделения ЛГ концентрация эстрадиола в фолликуле начинает снижаться. В преовуляторный период повышается содержание эстрадиола в яичниковой вене и в периферической крови. Частота пульсации ЛГ повышается, а амплитуда — снижается.
В позднюю фолликулярную фазу (2—0 дня до охоты, 18—21-й дни цикла) происходит увеличение синтеза и секреции эстрадиола клетками гранулезы (рис. 13). Преовуляторное нарастание уровня эстрадиола угнетает пульсацию ЛГ и снижает среднюю концентрацию ФСГ. По достижении определенного уровня эстрогенов у свиноматок проявляются признаки половой охоты и осуществляет-
104
|
|
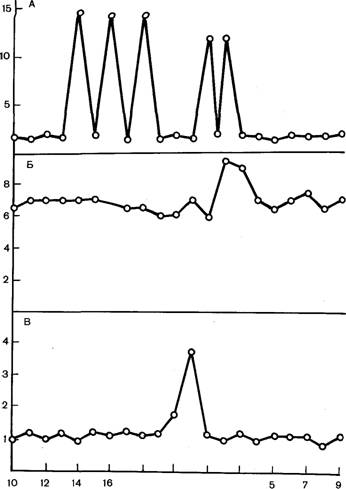
18 20) 3
Дни полового цикла
Рис. 12. Концентрация пролактина, ФСГ и ЛГ в плазме крови свиньи во время полового цикла:
А — концентрация пролактина, нг/мл; Б — концентрация ФСГ, нг/мл; В — концентрация ЛГ, нг/мл
ся преовуляторный выброс ЛГ из гипофиза. Это происходит через 2 суток после начала подъема уровня эстра-диола. В первый день цикла содержание в фолликулярной жидкости эстрадиола составляет 561 нг/мл, прогесте-
105

1 6 18 20 1 3
Дни полового цикла
Рис. 13. Уровни эстрогенов, простагландина и прогестерона в плазме крови свиньи, в период полового цикла:
А — концентрация эстрогенов, пг/мл; Б — концентрация простагландина Р 2а в маточной вене, нг/мл); В — концентрация прогестерона, нг/мл
рона — 236, андростендиона — 266 и тестостерона — 145 нг/мл. На 2-й день концентрация указанных гормонов составляет соответственно 38, 67, 76 и 19 нг/мл, а на 3-й — 27, 589, 18 и 12 нг/мл. Длительность повыше-
106
ния концентрации ЛГ составляет около 20 ч. Овуляция происходит через 36—40 ч после повышения уровня ЛГ. Увеличение концентрации ЛГ приводит к снижению уровня эстрадиола и в фолликулах, и в крови, способствуя лютеальным процессам в фолликуле. Обращает на себя внимание тот факт, что у свиней преовуляторный пик выделения ЛГ значительно менее выражен, чем у других видов животных. Максимальные значения циклического пика ЛГ только в 2 раза выше, чем амплитуды отдельных пульсаций ЛГ во время лютеальной фазы цикла. Вероятно, основным фактором, вызывающим овуляцию, является длительность повышенного уровня ЛГ.
Под влиянием выделяющегося ЛГ начинается лютеини-зация клеток гранулезы. При этом они теряют способность к ароматизации в них андрогенов и превращения их в эстрогены. В клетках гранулезы четко усиливается образование прогестерона. Однако этот прогестерон не поступает в кровь. Вероятно, его образование необходимо для внутренних процессов в яичнике и, в частности, для осуществления овуляции.
Помимо прогестерона в процессах овуляции принимают участие простагландины. Их концентрация в фолликуле возрастает в процессе его созревания и достигает максимума перед овуляцией. Овуляция происходит в первые 2 дня полового цикла. В этот период уровень ЛГ опускается до базисных значений. Концентрация ФСГ вновь повышается в связи со снижением уровня эстрогенов, а также с прекращением образования ингибина в фолликуле. Биологическое значение повышения уровня ФСГ в этот период заключается, вероятно, в росте новых фолликулов. Максимальной концентрации ФСГ достигает через 55 ч после пика ЛГ и продолжается 66 ч. Однако начало подъема совпадает с началом повышения преову-ляторной концентрации ЛГ. Между 8-м и 14-м днями цикла содержание ФСГ вторично повышается.
Концентрация прогестерона в крови возрастает со 2—3-го дня цикла и достигает максимума на 10—15-й день. Фактически выделение гормона желтым телом наивысшее на 10—12-й день, а в дальнейшем он поступает в кровь из жирового депо. До середины цикла секреция прогестерона не зависит от гипофизарных гормонов. Этот вывод подтвержается тем, что если после овуляции удалять гипофиз, то функция желтого тела не нарушалась. Введение же животным гонадотропных гормонов вызывало повышение секреции прогестерона только с середи-
107
ны полового цикла. Это указывает на то, что ЛГ необходим для сохранения желтого тела только в случае наличия супоросности. Как уровень ЛГ, так и ФСГ мало изменяется с 5-го по 18-й дни цикла. Содержание ЛГ низкое и постоянно колеблется с периодичностью 60—120 мин.
Лютеолиз желтого тела происходит примерно на 12— 16-й день цикла под влиянием образующихся в слизистой матки и поступающих оттуда в яичники простагландинов. В случае супоросности их образование тормозится эстрогенами, синтезируемыми в зародышах. Механизм действия простагландинов недостаточно ясен. Возможно, что он блокирует ЛГ-рецепторы. Причина выработки в матке простагландинов на 14—16-й день цикла неизвестна. Возможно, что в лютеолизе желтого тела принимает также участие пролактин. Его количество в этот период значительно повышается, а он ингибирует секрецию прогестерона из клеток гранулезы. Экзогенные простагландины, в отличие от других видов животных, не вызывают лютеолиза желтых тел у свиней до 12-го дня цикла. При введении свиноматкам в этот период простагландинов отмечается только кратковременное снижение уровня прогестерона. Функция же желтого тела и половой цикл в дальнейшем протекают нормально. На 16—18-й день цикла уровень прогестерона быстро снижается до базисных значений. В настоящее время возникают сомнения в том, что лю-теолиз желтого тела связан только с образующимися в матке простагландинами. В маточных венах содержание простагландинов в это время существенно возрастает, однако их количество в яичниковых артериях повышается незначительно. Кроме того, желтые тела сами способны вырабатывать простагландины.
Продуцируемые в яичниках в лютеальную фазу цикла эстрогены вырабатываются не в желтом теле, а в других структурах яичника. Высокий уровень прогестерона и определенная концентрация эстрогенов в лютеальную фазу цикла по принципу обратной связи тормозят выделение из гипофиза ФСГ и особенно ЛГ, уровень которого имеет в этот период только базисные значения. В результате в этот период происходит рост до определенной степени фолликулов, но не осуществляются их созревание и овуляция.
Концентрация пролактина в период полового цикла резко повышается дважды. Первое возрастание его уровня отмечается за 5—6 дней до охоты, а второе начинается незадолго до и кончается вскоре после преовуляторного
108
пика ЛГ. Значение пролактина в циклических процессах в этот период остается еще неясным.
Во многих исследованиях указывается, что случка свиноматок с хряком, особенно двойное спаривание, ускоряет процесс овуляции; Однако это противоречит тому факту, что пик ЛГ, запускающий механизм овуляции, отмечается незадолго до охоты и последующая случка уже не может оказать на него влияния. Возможность же воздействия нервных импульсов на процессы в фолликулах, приводящие к их овуляции, представляется маловероятной. Однако нельзя исключить возможность влияния нервных импульсов, возникающих при случке, на дополнительное выделение ЛГ из гипофиза, что может привести к некоторому ускорению овуляции.
Несмотря на хорошую изученность эндокринологии полового цикла, позволяющую успешно применять в производстве биотехнические приемы по стимуляции половой функции, синхронизации половой охоты и овуляции, необходимы более точные исследования в этом направлении. В свете современных представлений также не находят объяснения сравнительно частые случаи прихода в охоту свиноматок, оплодотворенных за 3 недели и менее. При этом, несмотря на наличие супоросности и достаточного числа эмбрионов, у свиноматок наблюдается полноценная охота, сопровождающаяся созреванием нормального количества фолликулов и их овуляцией. В результате в случае отсутствия эмбриональной смертности после обоих осеменений свиноматки вынашивают и рождают два помета с интервалом в 10 дней и более.
Эндокринология супоросности. При половом цикле с 10—12-го дня начинается регрессия желтого тела, и концентрация прогестерона в отходящих от яичников сосудах снижается. В случае же оплодотворения этого не происходит, что объясняется воздействием на процесс зародышей. Один из установленных факторов влияния — выработка ими значительных количеств эстрогенов на 12-й день после осеменения, т. е. непосредственно перед имплантацией. Результатом такого физиологического процесса является сохранение желтого тела. Если на 12-й день цикла ввести эстрогены неосеменявшимся свиноматкам, то это, как правило, приводит к ложной супоросности. На 3—5-й неделе супоросности желтые тела имеют такие же размеры, как и развитые циклические желтые тела. В то же время в них больше сосудов и окраска их до голубовато-красной. На разрезе они оранжевого цвета. Отлича-
109
ется также функциональная активность желтых тел супо-росности и полового цикла. В циклических желтых телах активность глютаматдегидрогеназы, гидроксиацилдегидро-геназы и конденсинг энзима ниже, а лактатдегидрогеназы выше, чем в желтых телах супоросности. Желтые тела су-поросности вырабатывают больше прогестерона, чем желтые тела полового цикла.
Концентрация прогестерона на 12—16-й день супоросности высокая и достигает 50 нг/мл плазмы. У свиней плацента продуцирует прогестерон, однако для сохранения супоросности в течение всего периода беременности необходим прогестерон, вырабатываемый желтыми телами. Специальные опыты показали, что для сохранения супоросности необходимо наличие минимум четырех-пяти желтых тел, так как концентрация прогестерона не должна быть ниже 4 нг/мл. Меньшее количество прогестерона приводит к эмбриональной смертности. В период ранней супоросности концентрация прогестерона находится в зависимости от количества желтых тел. В то же время выживаемость эмбрионов в этот период не зависит от уровня прогестерона, превышающего необходимый минимум. При экспериментальном снижении или повышении концентрации прогестерона в случае нормальной степени овуляции у свиноматок и оптимальном количестве плодов их выживаемость не изменялась.
Максимального развития желтые тела достигают к 75-му дню супоросности. Концентрация прогестерона в желтых телах повышается с 18-го по 40-й день быстро, снижается к 70-му дню, а затем остается примерно на одном уровне до 106-го дня. Содержание прогестерона в плазме крови снижается в последние дни супоросности.
У супоросных свиноматок, в сравнении с другими видами животных, значительно выше уровень образуемых эстрогенов. Основным среди них является эстронсульфат. На 4-й неделе супоросности их количество превышает в 10—15 раз уровень, наблюдающийся во время полового цикла. С 20-го дня супоросности эстронсульфат, который не находят у несупоросных свиноматок, с достаточной точностью можно определить в плазме крови. На основании установления эстронсульфата можно определять супоросность у свиноматок. Между 26-м и 30-м днями супоросности имеется прямая связь между концентрацией эстрогенов в моче и числом рождаемых поросят. Учитывая, что удаление яичников и надпочечников не приводит к исчезновению эстрогенов, вероятное место их
110
образования — плодная плацента. После снижения уровня эстрогенов к 50-му дню супоросности отмечается новый подъем их концентрации за 3 недели до опороса. К моменту родов содержание эстрогенов достигает высоких значений. Незадолго до опороса концентрация эстрогенов в плазме крови составляет 1—4 нг/мл, т. е. превышает более чем в 100 раз их концентрацию в период охоты.
В течение всей супоросности желтое тело вырабатывает пептидный гормон релаксин. Он считается гормоном беременности, так как не образуется в циклическом желтом теле. В течение супоросности он накапливается в лютеальных клетках желтого тела, а в предродовой период начинает выделяться в больших количествах в кровь. Его концентрация в плазме крови в начальный период супоросности составляет менее 1 нг/мл, повышается на 110-й день супоросности до 40 нг/мл, достигает за несколько часов до опороса максимальных значений в 145 нг/мл, а в первый день после родов вновь имеет значения ниже
1 нг/мл. Основная роль релаксина заключается в расслаблении связок сочленений подвздошных и крестцовых костей и расширении родового канала. В этом направлении он действует в синергизме с эстрогенами и простаглан-динами. Релаксин снимает твкжр тормозящее влияние прогестерона на сокращение мускулатуры матки в ответ на окситоцин.
Эндокринология опороса. Начиная с 4-й недели и до
2 дней до опороса концентрация прогестерона медленно снижается. Затем она начинает быстро падать. Однако низких значений прогестерон достигает только через несколько часов после завершения родов. Концентрация эстрогенов в последнюю неделю супоросности значительно возрастает. После опороса она быстро снижается до малых значений. Это также свидетельствует о том, что эстрогены в период супоросности вырабатываются плацентой. Концентрация кортикостероидов повышается в последние дни супоросности. При этом отмечаются существенные индивидуальные различия. Непосредственно перед опоросом и в его процессе уровень кортикостероидов еще более возрастает, а затем резко снижается. В крови плодов их содержание значительно выше, чем в крови матери. На основании современного уровня знаний взаимосвязь и роль гормонов в родовом процессе представляются следующими. В качестве важнейшего фактора, регулирующего родовой процесс, является повышение концентрации
111
глюкокортикостероидов в крови плодов. Их появление связано с усиленной секрецией АКТГ гипофизом и процессами созревания в надпочечниках плодов. Глюкокортикои-ды с кровью достигают плаценты и способствуют образованию там эстрогенов из вырабатываемых надпочечниками андрогенов. Повышенное количество эстрогенов сенсибилизирует матку к воздействию окситоцина. Одновременно они оказывают влияние на выработку простаглан-динов в плаценте и матке, которые обусловливают лютео-лиз желтых тел и снижение уровня прогестерона. Все это необходимо для лактации и стимуляции миометрия эстрогенами и окситоцином.
Окситоцин оказывает влияние на маточную мускулатуру только тогда, когда концентрация прогестерона снижается до 30 % от исходной. Это происходит незадолго до или к началу родов, поэтому окситоцин не может применяться для индукции родов. Его можно использовать только с целью усиления сократительной функции матки в родовой или послеродовой периоды. Максимальные количества окситоцина выделяются из гипофиза после выведения первого плода, что связано с нервными импульсами, вызванными механическим раздражением родовых путей.
Помимо указанных функций простагландины оказывают и прямое действие на стимуляцию мускулатуры матки. Они также способствуют выделению окситоцина из гипофиза матери и плодов, а он, в свою очередь, оказывает влияние на выделение простагландинов.
Простагландины способствуют секреции релаксина. Его повышенные количества появляются за 48 ч до выведения плодов.
Таким образом, пусковым механизмом родов являются кортикостероиды, а основным механизмом, осуществляющим процесс опороса,— простагландины. Кортикоиды в настоящее время используются для индукции опоросов. Широко применяются для индукции родов и прерывания супоросности простагландины.
Эндокринология лактационного периода. У свиней лактация оказывает тормозящее влияние на половую функцию. Подавляющее большинство свиноматок, за исключением первых дней после опороса, не приходят в охоту в период лактации. Причины блокады полового цикла в этот период находят только частичное объяснение. Основным фактором торможения полового цикла в период лактации является подсос поросят. При отъеме поросят в
112
любой период после опороса свиноматки через несколько дней приходят в охоту. Даже при временном отделении поросят от матки последние могут приходить в охоту. Другим необъяснимым фактором является то, что многие свиноматки приходят в охоту в течение первых 3 дней после опороса. Считается, что эта охота — следствие большого количества эстрогенов, образующихся в организме в родовой период, и она неполноценная, так как овуляции не происходит. Слученные в этот период свиноматки обычно не оплодотворяются. У убитых во время лактации свиноматок не обнаруживали желтых тел. В то же время имеются сообщения о том, что от слученных после родов свиноматок удавалось получить опорос. Отсутствие желтых тел в яичниках у лактирующих свиноматок свидетельствует о том, что если овуляция в этот период и происходит, то очень редко.
В период лактации отмечаются резкое повышение уровня пролактина в крови и низкое содержание ФСГ, особенно ЛГ. Это происходит, несмотря на незначительное содержание эстрогенов (менее 10 нг/мл), что по принципу обратной связи должно было усиливать выделение гонадотропных гормонов из гипофиза. В данном случае тормозящим фактором являются не яичники, о чем свидетельствуют опыты по их удалению. Удаление яичников в период цикла приводит к достоверному повышению уровня ЛГ, однако этого не отмечается в случае овариоэкто-мии у лактирующих свиноматок. Уровень ЛГ остается низким в течение всего периода лактации.
Инъекции окситоцина лактирующим свиноматкам не приводят к достоверным изменениям в гипофизе и яичниках. В фолликулах при этом уменьшается количество жидкости. Тормозящее влияние лактации на функцию яичников особенно выражено в, течение первой недели после опороса. К концу этого периода в яичниках достоверно уменьшаются число фолликулов и их размеры. Это явление связано не только с низкой гонадотропной активностью гипофиза, но и с пониженной чувствительностью к ним яичников. Экзогенное введение свиноматкам в период ранней лактации даже больших количеств гонадотропных гормонов оказывается, как правило, малоэффективным. С увеличением продолжительности лактации тормозящее влияние сосания на гипофиз и яичники медленно, но постоянно снижается. Это приводит к повышению уровня ФСГ и ЛГ в организме. В результате отмечается рост новой группы фолликулов, и у отдельных сви-
113
номаток возможна полноценная охота. После отъема поросят уровень лактогенного гормона в организме свиноматок резко снижается, а секреция ЛГ и ФСГ возрастает. Это вызывает созревание фолликулов и появление характерных циклических изменений в организме свиноматок. На 4—6-й день после отъема поросят в результате повышенной секреции эстрогенов яичниками свиноматки приходят в охоту. У них регистрируют преовуляторный пик ЛГ и овуляцию. При раннем отъеме поросят (3 не-, дели) преовуляторный пик ЛГ менее выражен. Однако это не сказывается отрицательно на степени овуляции. Установленное после раннего отъема поросят снижение многоплодия объясняется не уменьшением количества овулирующих фолликулов, а повышением роли эмбриональной смертности.

