




Постепенно сперматида вытягивается, осевая нить удлиняется и в итоге формирует тело и хвост спермия. В области тела осевую нить обвивает спиральная нить из митохондрий. Последующая часть осевой нити покрыта слоем из плотных фибрилл. Они имеют вид полукружных ребрышек, своими концами соприкасаются и сливаются с двумя узкими продольными колонками, расположенными по противоположным сторонам хвостика.
На шейке полностью сформированного спермия остаются зернистые тельца Гольджи. Они исчезают вместе с невключенной в клетку цитоплазмой. Но в этом месте появляется цитоплазматическая капелька. Она обнаруживается до тех пор, пока спермий связан с сертолиевыми клетками, и исчезает в период прохождения его по каналу придатка семенника.
Образование спермиев у самцов после достижения половой зрелости происходит непрерывно. Активность сперматогенеза волнообразно перемещается по семенным канальцам. Между двумя последовательно появляющимися группами однотипных клеток в одном участке канальца проходит: у хряка – 8,6 дня, у барана – 10,3, у жеребца – 12,2, у быка – 13,5 дня. Это семенной эпителиальный цикл. В нем иногда различают 8 стадий (Ortavant, 1977). У быка для полного формирования спермиев из сперматогоний необходимо около 60 дней. Для продвижения по каналу придатка семенника требуется еще 8-14 дней. Головку и тело придатка спермий проходят за 5 дней, а хвост придатка – за 5-9 дней, в зависимости от интенсивности полового использования производителя.
Передвижение спермиев по внутренним протокам осуществляется за счет: положительного давления в канальцах вследствие постоянного вхождения в их просвет новых клеток; отрицательного давления в семявы-носящих канальцах и канале придатка семенника, которое создается в результате эякуляций и всасывания жидкости, поступающей в большом количестве из семенника; движения ресничек, выстилающих сёмявы-носящие канальцы и начальную часть канала придатка и, наконец, давления на семенники, возникающего в процессе движения животного. Попав в канал придатка, спермин продвигаются дальше за счет перистальтических движений его стенок.
В извитых канальцах семенника впервые спермин могут быть обнаружены в возрасте около 5 мес. – у баранов и хряков, 7,5 мес. – у быков и 13 мес. – у жеребцов. Способность же к воспроизведению у них появляется несколько позднее и связана с началом образования спермы, содержащей достаточное количество нормальных спермиев, а также с проявлением полового влечения. Факторы внешней среды (температура, продолжительность светового дня, кормление и содержание, возможные болезни) оказывают большое влияние на сроки достижения половой зрелости у самцов.
Образование и продвижение спермиев по половым протокам находится под влиянием гормонов гипофиза и семенников. Определенную роль играют также гормоны надпочечников и щитовидной железы.
Еще до наступления половой зрелости семенники подвергаются воздействию ФСГ и ЛГ. Лютеинизирую-щий гормон (гормон, стимулирующий интерстициалъные клетки) воздействует на рост и функцию клеток Лейдига, повышает секрецию и выделение андрогенов. Действие осуществляется через аденилатциклазу, путем сокращения до минимума периодов стероидогенеза. У быков интенсивность секреции и выделения половых гормонов повышается с 4-6-месячного возраста. Вскоре начинается и выделение фруктозы придаточными половыми железами.
У половозрелых животных выделение ЛГ происходит нерегулярно, эпизодически, с промежутками в 2-4 ч (3-7 раз в течение дня). Характер выделения ФСГ расшифрован недостаточно полно.
Примерно через 40 мин после каждого пика ЛГ следует пик тестостерона, затем через 40-80 мин содержание андрогена снижается до исходного уровня. Андрогены повышают чувствительность сперматогенного эпителия семенных канальцев к ФСГ. Этот гормон, вызывая размножение сперматогоний, дает начало сперматогенезу.
ФСГ воздействует и на клетки Сертоли, стимулирует секрецию андрогенсвязывающего протеина (АСП) и превращение тестостерона в эстрогены. Секреция ФСГ регулируется половыми гормонами иингибином, а также регуляторным протеином, секретируемыми клетками Сертоли.
Андрогены воздействуют на весь половой аппарат самца, стимулируют рост и функционирование придаточных половых желез, тем самым влияя на состав спермы, обеспечивают оптимальные условия для сперматогенеза, продвижения и созревания спермиев в канале придатка семенника. После превращения в мозгу в эстрогены тестостерон регулирует выделение ЛГ и влияет на поведение самца.
В семенных канальцах тестостерон превращается в 5-дигидротестостерон, который более устойчив к метаболическим превращениям. Здесь оба этих метаболита связываются с андрогенсвязывающим протеином. АСП обеспечивает высокую концентрацию тестостерона в извитых канальцах и канале придатка семенника (в 100-300 раз выше, чем в периферической крови), что важно для нормального сперматогенеза. Абсорбируется протеин в канале придатка семенника. Высокая концентрация тестостерона поддерживается также в сети семенника, семявыносящих канальцах и проксимальной части придатка.
Для поддержания постоянного нормального сперматогенеза требуется совместное воздействие ФСГ и андрогенов, вырабатываемых под влиянием ЛГ. В связи с этим должно быть строго определенное соотношение между этими гормонами. Уменьшение секреции гонадотропных гормонов тормозит продукцию андрогенов и ослабляет сперматогенез. Понижение уровня андрогенов стимулирует дополнительное выделение гонадотропинов, которые снова повышают секрецию андрогенов и образование спермиев.
У самцов, используемых в течение года, постоянный уровень секреции гонадотропинов. У сезонно используемых животных секреция ФСГ и ЛГ снижается вскоре после окончания случного сезона и остается низкой до следующего сезона. С наступлением его внешние факторы (продолжительность светового дня, температура и др.) дают толчок к выделению гипоталамусом ГнРГ, который в свою очередь стимулирует секрецию и выделение ФСГ и ЛГ гипофизом. У баранов в регуляции сперматогенеза важную роль играет щитовидная железа. Введение им тироксина летом, когда тормозится образование спермиев и увеличивается число их с различными аномалиями, способствует уменьшению таких клеток и повышению образования нормальных спермиев.
Образовавшиеся в процессе сперматогенеза спермин продвигаются по внутренним половым протокам и скапливаются в канале придатка семенника, который представляет собой значительную емкость. У быка в каждом придатке содержится до 4 мл сливочно-белого цвета вязкой жидкости с концентрацией спермиев от 2,5 до 5 млрд./мл. Половые клетки занимают примерно половину объема этой жидкости. Всего оба придатка у быка могут вмещать до 20 и даже до 40 млрд. спермиев, у барана - до 60 млрд.
Секреторная функция придатка семенника и передвижение спермиев по половым протокам регулируются окситоцином.
Поступающие в канал придатка семенника спермин имеют в месте соединения головки и тела цито-плазматическую капельку. По мере прохождения их по каналу капелька продвигается по направлению к хвосту и затем сбрасывается. С цитоплазматической капелькой связывают приобретение спермиями липопротеидного покрова (оболочки).
В канале придатка спермин приобретают подвижность и оплодотворяющую способность. У большинства видов половые клетки, взятые из семенников самца, не способны к движению, хотя они живые и могут совершать слабые колебательные движения хвостиком. У быка спермин из проксимальной и средней части головки также неподвижные, но подвижность появлялась в участках придатка вблизи семенников и уже хорошо проявлялась у спермиев из хвоста придатка. У быков нормальная подвижность и биологическая полноценность спермиев в канале сохраняется в течение 30-35 дней, а отдельные спермин могут сохранять подвижность 2-3 мес. Спермин кроликов сохраняют жизнеспособность 38 дней.
СПЕРМА: СОСТАВ И СВОЙСТВА
Семенная жидкость (сперма) состоит из двух основных частей: спермиев и жидкой части - плазмы. Семенники вырабатывают спермиев, а также являются источником различных клеточных фрагментов и жидкости, не всосавшейся при прохождении по внутренним половым протокам. Одним из компонентов этой жидкости является тестостерон. Жидкая часть плазмы является продуктом деятельности придаточных половых желез, канала придатка семенника и ампул спермиопроводов.
ВНЕШНИЕ СВОЙСТВА СПЕРМЫ
Количество и внешний вид спермы, соотношение жидкой части и спермиев и многие другие свойства варьируют в зависимости от вида животных и их индивидуальных особенностей. Жвачные выделяют небольшие эякуляты с высокой концентрацией спермиев. Жеребец, хряк и кобель дают большие эякуляты, но с низким содержанием половых клеток. У птиц объем эякулятов небольшой, а концентрация спермиев очень высокая (табл. 1).
1. Объем эякулята и концентрация спермиев
| Производитель | Объем эякулята, мл | Концентрация спермиев, млрд./мл |
| Бык | 4 (2-10) | 1,0 |
| Баран | 1 (0,6-2) | 3,0 |
| Хряк | 200 (100-500) | 0,15 |
| Жеребец | 75 (20-300) | 0,15 |
| Кобель | 3 (2-30) | 0,15 |
| Кролик | 0,5 (0,2-2) | 0,2 |
| Петух | 0,5 (0,2-0,6) | 3,8 |
| Индюк | 0,3 (0,2-0,4) | 7,5 |
| Гусак | 0,5 (0,2-1,3) | 0,5 |
Цвет и консистенция спермы зависят в основном от концентрации спермиев: чем выше их концентрация, тем более вязкая консистенция спермы, тем белее цвет. Сперма быка и барана обычно сливочно-белого цвета, но отдельные быки выделяют сперму желтого цвета. Сперма птиц молочно-белого или слегка желтоватого цвета, сливкообразной консистенции. Сперма жеребца и хряка серовато-белого цвета, водянистой, иногда вязкой консистенции.
СОСТАВ СПЕРМЫ
Соотношение отдельных составных частей спермы у различных видов животных различное. У барана объем эякулируемой жидкости из придатков семенников достигает 0,86 мл с концентрацией спермиев более 5 млрд./мл. Половые клетки составляют 30% объема всей спермы, секрет придатков - 50-60%, остальная часть спермы представлена секретами придаточных половых желез. Придатки семенников быка выделяют 0,6 мл жидкости с содержанием спермиев 4,9 млрд. /мл и общим количеством их около 3 млрд. Спермин с секретом придатков составляют 15% объема эякулята, секрет пузырьковидных желез - до 40%, предстательной железы - 5-6%, куперовых и уретральных желез - 35-40%. У хряка спермин с секретом каналов придатков семенников составляют 3% (2-5%) объема эякулята, причем основная доля приходится на половые клетки. Пузырьковидные железы выделяют до 26% объема семенной жидкости, куперовы – 19% (10-25%), предстательная железа - более 50%. Желеобразная фракция составляет в среднем 18% всего эякулята. У жеребца эякулят формируется в основном за счет секрета придаточных половых желез и в особенности – предстательной. У кобеля секрет семенников и придатков семенников оставляет 0,5-3,5 мл, уретральных желез – 0,25-5 мл и предстательной железы – 2-30 мл. Сперма содержит 85-98% воды и сухого вещества.
СОСТАВ СПЕРМЫ
Секрет придатка семенника имеет кислую реакцию (рН 5,8-6,2 у быка, 5,6-6,6 у хряка), высокое парциальное давление углекислого газа и высокое осмотическое давление, содержит мало электролитов, особенно ионов натрия и хлора. Фруктозы также содержится немного или она вообще отсутствует. Такой состав секрета благоприятствует длительному (до 30 дней или более) сохранению оплодотворяющей способности спермиев, находящихся в канале придатка в состоянии относительного покоя. Секрет придатка богат веществом липидного характера – глицерилфосфорилхолином (у кролика – фосфорилхолин). Особенно много его в плазме барана. Под влиянием фермента фосфатазы фосфорилхолин распадается с образованием свободной фосфорной кислоты и холина. Фосфорная кислота используется в процессе гликолиза. Глицерилфосфатилхолин – более стойкое соединение. Секрет придатка содержит также общий белок и белок, связанный с андрогеном, андрогены, фосфолипиды, значительное количество свободных аминокислот и аминосахаров, инозитол, прегнолон (у барана), в небольшом количестве – лимонную кислоту и эрготионин (у хряка).
Секрет ампул спермиопроводов не имеют ампулообразных расширений. Эякуляция у них происходит медленно и спермии поступают в уретру небольшими порциями непосредственно из канала придатка семенника.
Секрет пузырьковидных желез имеет слабокислую реакцию. У жвачных он водянистый и прозрачный, хотя у быков иногда приобретает желтоватый цвет; у хряка и жеребца секрет мутный и сильно слизистый, особенно - у жеребца. Главными компонентами секрета пузырьковидных желез являются лимонная кислота и фруктоза. Выделение их регулируется тестостероном. У хряка редуцирующие вещества (сахара) представлены не фруктозой, а серусодержащим основанием – эрготионином и инозитолом. В секрете содержатся также аскорбиновая кислота, липиды, белки и свободные аминокислоты – аланин, глицин, серии, тирозин. Из минеральных веществ в секрете много калия, меньше натрия, очень мало хлора. У барана в секрете содержится простагландин.
Секрет предстательной железы водянистый, имеет слабокислую (рН 6,5) или нейтральную реакцию. Содержит много протеолитических ферментов и свободных аминокислот, особенно глютаминовой кислоты; чрезвычайно богат электролитами - натрием, калием, хлором, кальцием, бикарбонатом, цитратом, причем преобладает хлористый натрий (у кобеля до 90%); содержит секрет и лимонную кислоту, простагландины и антагглютинин – вещество, предохраняющее спермиев от агглютинации. У кролика секрет предстательной железы насыщен форменными элементами и имеет вид мутновато-белой жидкости, похожей на сперму.
Куперовы железы выделяют щелочной секрет (рН 7,8-8), прозрачный и тягучий, альбуминоидной природы. Выделяется в начале эякуляции, промывает уретру. У хряка он выделяется в конце семяизвержения и, смешиваясь с секретом пузырьковидных желез, образует крупные, быстро набухающие зерна, препятствующие вытеканию спермы из половых путей свиньи. У барана секрет этих желез жидкий, богат фруктозой.
В целом секрет придаточных половых желез и канала придатка семенника (плазма спермы) имеет реакцию слабокислую (рН 6,5-6,7 у барана и быка) или слабощелочную (рН 7,2-7,3 у хряка и жеребца). Величина рН различных образцов спермы заметно отличается от средних показателей и при хранении неразбавленной спермы не является стабильной. Так, рН спермы быка и барана снижается в результате образования и накопления молочной кислоты.
В процессе эякуляции секрет придаточных половых желез смешивается с секретом придатка семенника, разбавляет его и активизирует движения спермиев.
Важными характеристиками плазмы спермы являются буферность и осмотическое давление.
Буферностъ (свойство устойчиво сохранять свою реакцию) спермы обусловлена в основном солями слабых кислот - карбонатами, цитратами и фосфатами. Эти соли могут передавать свой металл более сильным кислотам и таким образом заменять их более слабыми, которые дают во много раз меньше ионов водорода. Определенную роль в обеспечении буферности спермы играет и белок. Буферность спермы слабее буферности крови. Наиболее сильная она в сперме быка и барана.
Осмотическое давление спермы определяется количеством растворенных в плазме веществ: минеральных и органических. Причем у различных видов животных количество этих веществ различно. Так, у хряка основными элементами, создающими осмотическое давление, являются инозитол и калия цитрат; у кобеля -натрий хлористый. Выражается осмотическое давление в атмосферах; колеблется в пределах 6,7-8,7 атм. Определяется этот показатель на основании величины депрессии, т. е. точки замерзания, которая для спермы животных равна примерно -0,6°С, и, исходя из правила, что раствор, содержащий в 1 л грамм-молекулу неэлектролита, дает депрессию -1,86°С и имеет осмотическое давление 22,4 атм. Для получения равных сперме по осмотическому давлению растворов (изотонических) необходимо брать 1/3 грамм-молекулы неэлектролита на 1 л дистиллированной воды. Если берется электролит, то количество вещества уменьшается во столько раз, на сколько ионов диссоциирует его молекула. Например, для приготовления изотонического раствора хлористого натрия необходимо взять не 1/3, а 1/6 грамм-молекулы, т. е. 58,5:6.
МОРФОЛОГИЯ И ХИМИЧЕСКИЙ СОСТАВ СПЕРМИЕВ
Наиболее важной составной частью спермы являются спермин. От их качества зависит оплодотворяющая способность семенной жидкости, а также жизнеспособность и наследственные свойства получаемого потомства. Однако и плазма спермы может в значительной степени обусловливать качество спермы в целом и, главным образом, ее оплодотворяющую способность.
Впервые спермин обнаружил Иоган Гамм. О его наблюдениях сообщил Левенгук в 1677 г. С тех пор эти клетки стали предметом многих научных исследований, в результате которых были выяснены ультраструктура, химический состав и основные свойства их, хотя до настоящего времени знания о спермиях далеко не полные.
Спермин сельскохозяйственных животных представляют собой небольшие подвижные клетки длиной от 0,05 до 0,08 мм. Величина их не зависит от величины животных. Спермин состоят из головки, шейки, тела (средней части) и хвоста. У быка общая длина спермия 70 мкм, в том числе головки – 8-10 мкм, шейки – 1 мкм, тела – 8-10 мкм и хвоста – 50 мкм. Спермин барана имеют длину 69 мкм (66-72), жеребца – 59 мкм (58-60), хряка – 56 мкм (55-57) (рис. 25).
Головка плоская и при фиксации на предметном стекле обращена к нам одной из плоских сторон. Ширина  ее около 4 мкм, толщина – 1 мкм. Одна из плоских поверхностей головки слегка вогнута (имеет ложкообразное углубление) и это обусловливает поворачивание спермия вокруг своей оси при движении вперед. По происхождению головка представляет собой ядро зародышевой клетки и в основном заполнена хромати-новым материалом. Этот материал сконцентрирован в нижней части головки, почти отсутствует в средней части и как бы соединен в цепочки в передней части. В головке имеются нити, которые играют роль опорных ("скелетных") образований. Различают продольные и спиральные нити. Продольные обусловливают длину и изгиб головки, спиральные (две или три) – определяют размеры головки в поперечнике. Передняя часть головки на 2/3 покрыта акросомой, которая образуется из акросомного зернышка и вакуоли. Акросома плотно прилегает к ядерной оболочке и прочно держится на головке. Передний конец акросомы утолщен и загнут в одну сторону, как и сама головка. Задний край прикреплен в виде довольно широкой полосы – ядерного кольца. Если хранить сперму при температуре 16-37°С, то уже через несколько часов отмечаются дегенеративные изменения в акросоме. У некоторых спермиев она набухает, отделяется (кроме ядерного кольца) и ее можно рассмотреть как отдельное тело. При повреждении акросомы спермий теряет способность к оплодотворению.
ее около 4 мкм, толщина – 1 мкм. Одна из плоских поверхностей головки слегка вогнута (имеет ложкообразное углубление) и это обусловливает поворачивание спермия вокруг своей оси при движении вперед. По происхождению головка представляет собой ядро зародышевой клетки и в основном заполнена хромати-новым материалом. Этот материал сконцентрирован в нижней части головки, почти отсутствует в средней части и как бы соединен в цепочки в передней части. В головке имеются нити, которые играют роль опорных ("скелетных") образований. Различают продольные и спиральные нити. Продольные обусловливают длину и изгиб головки, спиральные (две или три) – определяют размеры головки в поперечнике. Передняя часть головки на 2/3 покрыта акросомой, которая образуется из акросомного зернышка и вакуоли. Акросома плотно прилегает к ядерной оболочке и прочно держится на головке. Передний конец акросомы утолщен и загнут в одну сторону, как и сама головка. Задний край прикреплен в виде довольно широкой полосы – ядерного кольца. Если хранить сперму при температуре 16-37°С, то уже через несколько часов отмечаются дегенеративные изменения в акросоме. У некоторых спермиев она набухает, отделяется (кроме ядерного кольца) и ее можно рассмотреть как отдельное тело. При повреждении акросомы спермий теряет способность к оплодотворению.
Шейка, тело и хвостик спермиев представляют собой единую двигательную систему. Внутри их находится осевая нить, которая образуется из дистальной центриоли. Состоит осевая нить из двух центральных фибрилл, окруженных тонкими двойными фибриллами. В теле спермия имеется еще девять грубых фибрилл. Тонкие и грубые фибриллы окружают две центральные в виде двух концентрических колец. Продольные фибриллы окружены спиральной нитью.
Тонкие фибриллы, состоящие из двух трубочек, по диаметру сходны с центральными, расположены на некотором расстоянии вокруг центральных. Каждая пара тесно сращена между собой, образует дуплет. Одна из пары – субфибрилла В – кажется прозрачной внутри так же, как и центральные, вторая -- субфибрилла А –заполнена непрозрачной массой и несет на наружной стороне, обращенной к другой паре, два выступа -ручки. Субфибриллы А соединяются с центральными фибриллами тонкими нитями - спицами. Пять субфибрилл А соединены с одной центральной и четыре - с другой центральной фибриллой. Стенки двух центральных фибрилл состоят из 13 рядов глобулярных белковых молекул. Периферические дуплеты из двух соединенных между собой трубочек имеют общий участок стенки. В них молекулы расположены в 23 ряда. Белок – спактин (тубулин), сходен по своему составу с актином. Отростки (ручки) состоят из молекулы белка спермозина (денеина), который, как и миозин, способен при определенной концентрации ионов кальция и магния действовать как фермент, расщепляющий АТФ, и высвобождать энергию.
В области шейки, внутри так называемой имплантационной ямки, девять грубых фибрилл соединяются одна с другой, образуя косо поставленную рамку с отверстием внутри. Несколько ниже в глубине их располагаются верхние концы тонких фибрилл. Грубые фибриллы – первая, пятая и шестая – в диаметре больше других. Положение их таково, что ось симметрии проходит от первой между двумя центральными и между пятой и шестой грубыми. Грубые фибриллы имеются только в области шейки и тела спермия и дальше постепенно истончаются и исчезают.
В области шейки и тела грубые фибриллы окружены митохондриальным слоем. Этот слой состоит из отдельных удлиненных митохондрий, соединенных между собой концами и закрученных по спирали вокруг продольных фибрилл. Число витков варьирует от 12 до 80. Внутри каждой митохондрии находятся 2-3 продольные перегородки – кристы, пространство между ними заполнено липидами. В митохондриях сосредоточены ферментные системы спермиев и в них вырабатывается энергия для движения. В хвостовой части спермия кольцо из 9 пар тонких фибрилл окружено тонкой спиральной нитью немитохондриального происхождения. Она связывает фибриллы, препятствует распаду их при движении. Только самый кончик хвоста не обвит этой нитью и в стареющем спермин нередко образует кисточку из разделившихся фибрилл. Снаружи спермий покрыт полупроницаемой тонкой оболочкой (мембраной), имеющей липидный слой. Появление его связано с продвижением вдоль осевой нити цитоплазматической капельки. Увеличение в эякуляте числа спермиев, имеющих в области шейки такую капельку, обусловливает понижение плодовитости самца.
Повреждения головки, которые проявляются неравномерностью формы ядра и многочисленными пальцевидными углублениями мембраны ядра у переднего края, а также наличие в акросоме кист и пузырьковидных структур и образований в виде выростов также могут быть причиной бесплодия отдельных особей. У спермиев кобелей грушевидная форма и другие дефекты спермиев нередко сопровождаются нарушениями акросомы и обусловливают бесплодие. У бесплодных жеребцов обнаруживали увеличение числа микротрубочек. В отличие от нормальных дефектные фибриллы отличались сильной извитостью, не были связаны между собой и на 20% были тоньше в диаметре.
Спермий содержат примерно 75% воды и 25% белков и жиров; химические компоненты: дезоксирибо-нуклеопротеид (в головке спермия), протеиносвязанный мукополисахарид (в акросоме), белки, липопротеиды и фосфолипиды (в теле и хвостике), кератиноподобный протеин (в мембране и фибриллах). Кроме того, в спер-мии содержатся минеральные вещества, разнообразные ферменты и коферменты.
Дезоксирибонуклеопротеид состоит из ДНК и протеида. ДНК половых клеток различных видов отличаются друг от друга соотношением главных составных частей: аденина, гуанина, цитозинаи тимина. Протеин (гистон) содержит 17 аминокислот, богат аргинином, но также имеет значительные видовые различия. Отклонения от нормального содержания и состава дезоксирибонуклеопротеида в клетках приводит к понижению оплодотворяющей способности спермы.
Мукополисахарид входит в состав сложного липо-гликопротеинового комплекса. Содержится главным образом в акросоме и играет важную роль в проникновении спермия через лучистый венец яйца. Причем такая способность половой клетки связана с гликопротеидами, которые появляются в акросоме в хвосте придатка. Главными компонентами мукополисахари-да являются сиаловая кислота, метилпентоза, галактоза, манноза, гексозамин. В акросоме содержатся и фруктоза, илипиды.
Шейка, тело и хвост спермиев состоят из простых белков, сложного белка липопротеида, свободных липи-дов и солей. Из простых белков, которые содержатся в фибриллах, можно выделить спактин и спермозин. Липопротеиды составляют значительную часть сухого вещества тела и хвоста спермия. Свободных жиров в этих частях клетки – 10-14%. Основным из них является плазмоген. Это альдегидной природы холиносо-держащий жир. Он может служить субстратом для эндогенного дыхания спермиев. Вторая часть свободных жиров приходится на лецитин. Помимо азотистого основания холина в лецитине и плазмогене содержится фосфорная кислота. Липиды находятся в основном в хвостике, меньше в средней части и головке. В спермиях содержится также небольшое количество обычных жиров (триглицеридов) и немного холестерина.
Цитоплазматическая мембрана спермиев состоит в основном из белковой субстанции, обладающей многими свойствами кератина. Она богата цистином, гистидином и аргинином и имеет высокое содержание серы.
Оболочка устойчива к разлагающему действию пепсина, трипсина, а также к щелочным и кислым растворам.
Ферменты и коферменты находятся в основном в головке и средней части и в меньших количествах в хвостике. В области головки (в акросоме) содержатся гиалуронидаза, которая может свободно выделяться в окружающую среду, и три формы (альфа, бета и гамма) акрозина, по своему действию напоминающего трипсин. Ферменты гидролитического и окислительного характера связаны со структурой спермиев и размещены на кристах или на поверхности митохондриальных нитей. Гликолитические ферменты (щелочная и кислая фосфатазы) участвуют в гликолитическом цикле. Они содержатся и в плазме спермы; вырабатываются в предстательной и пузырьковидных железах. Цитохромная система (цитохромоксидаза, аминокислотная оксидаза, различные типы дегидрогеназ и др.) играет важную роль в окислительном метаболизме (дыхании) и концентрируется в митохондриальной оболочке средней части спермиев.
Половые клетки содержат много фосфора и серы. В их головках фосфор связан с ДНК.
Спермин обладают подвижностью. Подвижность чрезвычайно важна и необходима для распределения их в половых путях самки, где находится яйцо. В результате активных движений спермиев происходит встреча их с яйцом и проникновение через его оболочки. Движение осуществляется за счет сокращений средней части и хвоста, в которых функционирует сократительный комплекс: спермозин – спактин, АТФ и фосфатаза. Поступательное движение спермиев обеспечивается в результате ритмичных сокращений хвостовой нити, происходящих в плоскости головки, и вследствие вращения головки вокруг своей продольной оси. Начало сокращения возникает в шейке и захватывает последовательно парные периферические осевые фибриллы по кругу с последующим их расслаблением. Две центральные фибриллы дают импульсы, координирующие такие последовательные сокращения и расслабления по всей длине каждой пары осевых фибрилл. Ввиду того что головка имеет с одной стороны небольшую вогнутость, при движении вперед она поворачивается вокруг своей продольной оси. Под микроскопом это проявляется вспышками света, отраженного головкой. Вследствие вращения головки хлыстообразные удары хвоста, происходящие в плоскости головки, приходятся фактически на разные стороны и это обеспечивает прямолинейно-поступательное движение. В свежей сперме благодаря наличию отрицательного заряда спермин не склеиваются, располагаются параллельно, и удары их хвостиков происходят ритмично и в такт. Это способствует одновременному направленному в одну сторону передвижению половых клеток и создает сильное вихревое движение жидкости, которое может уносить их самих, если скорость этого движения больше их собственной скорости. Вихревое движение заметно невооруженным глазом в густых эякулятах барана. Скорость движения спермиев в среднем составляет около 5 мм/мин. Зависит она от температуры, рН, вязкости среды и других факторов.
Важной особенностью спермиев является способность их к реотаксису, т.е. к ориентированному движению против тока жидкости. Медленное течение жидкости не только определяет направление их движения, но и стимулирует возрастание его скорости. В связи с этим многие полагают, что приобретение таких свойств спермиями обусловлено особенностями осеменения млекопитающих, в частности, характером движения жидкости (слизи) в половом тракте самок (В.К.Милованов).
ОБМЕН ВЕЩЕСТВ В СПЕРМИЯХ
Для совершения сократительных движений хвостиком спермиям необходима энергия. Ее они получают за счет распада аденозинтрифосфата. АТФ из митохондрий спиральной нити, обвивающей тело и шейку спермия, переходит на фибриллы и разлагается с высвобождением энергии.
АТФ состоит из аденина и рибозы (аденозина) и трех молекул фосфорной кислоты. Связи последних двух фосфатных групп богаты энергией и могут быть нарушены только путем затраты большого количества энергии. Под воздействием фосфатазы АТФ разлагается с образованием АДФ и фосфорной кислоты и высвобождением энергии:
 |
Связь второй молекулы фосфорной кислоты в АДФ также богата энергией и может быть разрушена с выделением энергии:
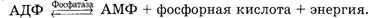 |
Высвобождаемая при разрыве этих связей энергия используется как механическая - для движения или как химическая - для биосинтеза, или же выделяется в виде тепла. Для того чтобы движение спермиев не прекращалось, требуется постоянное восстановление запасов АДФ и АТФ. Это и происходит в клетках, так как указанные реакции обратимы. Но для реосинтеза АТФ из АДФ или АДФ из АМФ путем присоединения фосфатных групп необходима энергия из внешнего источника. Таким источником в сперме являются углеводы, главным образом глюкоза, фруктоза и сорбит, а также жиры, аминокислоты, липиды. Использование этих веществ и дает энергию.

