




Сезон рождения оказывает заметное влияние на сроки наступления половой зрелости у кобыл, сук и особенно у овец. Так, если ярочка родится зимой или в начале весны и будет нормально развиваться, охота у нее проявится в этот же год осенью, хотя ей будет только 5-8 мес. от роду. Но если рождение произойдет поздней весной, то она не достигнет такого состояния, когда у нее может проявиться охота в этот половой сезон (осенью текущего года). Первая охота в таком случае наступит лишь в начале осени следующего года, т. е. на втором году жизни.
РЕПРОДУКТИВНЫЙ ПЕРИОД
С достижением половой зрелости у самок, проявляется инстинкт спаривания, а их половые железы вырабатывают способные к оплодотворению половые клетки. При этом их половой аппарат подчиняется определенному функциональному ритму. Процессы, которые лежат в основе этого ритма и обусловливают его, происходят в яичниках. Они заключаются в периодическом созревании одного или нескольких фолликулов, овуляции и образовании желтого тела (тел). Внешним проявлением этих изменений является течка (выделение слизи из половых органов) и половая охота (готовность самки к спариванию с самцом).
Определенная периодичность изменений в половых органах самки дала основание говорить о циклической природе половой функции. Впервые в литературу термин "половой цикл" ввел Маршалл. Под половым циклом следует понимать все многообразие структурных и функциональных изменений, происходящих в половом аппарате, эндокринной и других системах организма самки от одной половой охоты до другой (или от одной овуляции до другой). Период времени между двумя половыми охотами (или овуляциями) составляет продолжительность полового цикла.
Число половых циклов и их продолжительность не одинаковы у различных животных. Многие дикие животные на протяжении года имеют только один половой цикл. Это моноциклические животные. Время года, когда у них проявляется половая активность, называют половым сезоном. Собак и кошек также относят к моноциклическим животным, потому что у них промежутки между половыми циклами длительны. В течение года у сук течка проявляется обычно два раза: поздно зимой и в конце лета. Но нередко у них наблюдается четыре цикла в году.
Сезонность в размножении диких млекопитающих в большинстве случаев можно объяснить естественными причинами и прежде всего теми требованиями, которые предъявляет потомство к окружающей среде. Наличие тепла и света, водопоя, легкодоступного и усвояемого корма благоприятствует появлению потомства и облегчает ему дальнейшую жизнь. Тем самым эти факторы определяют время наступления полового сезона.
У сельскохозяйственных животных вследствие улучшения условий кормления и содержания и ослабления сезонных различий в их содержании, а также ослабление неблагоприятных климатических воздействий на них половой сезон удлинился, а некоторые виды (коровы, свиньи) могут проявлять половую цикличность на протяжении всего года. Таких животных относят к полициклическим. У овец и коз отмечается несколько половых циклов, следующих один за другим, после чего половая функция ослабевает. Это – полициклические животные с выраженным половым сезоном.
Учение о половом цикле впервые сформулировал Хипп. Он выделил в половом цикле следующие стадии (фазы).
Проэструс – подготовительный период, непосредственно предшествующий эструсу. Характеризуется началом быстрого роста отдельных фолликулов в яичниках. Растущие фолликулы вырабатывают эстрогены. Под их влиянием усиливается кровоснабжение половых органов, вульва набухает, слизистая влагалища приобретает красный цвет вследствие увеличения притока крови. Клетки ее подвергаются изменениям, которые у большинства видов приводят к ороговению. Секреция слизи клетками влагалища и шейки матки возрастает.
Матка увеличивается, слизистая оболочка ее становится кровенаполненной и отечной, а маточные железы – активными. У сук в этой стадии выделения из половых органов имеют кровянистый оттенок.
Эструс – кульминационный момент полового цикла. В этой фазе в центральной нервной системе животного появляется очаг повышенной возбудимости, который приводит к развитию половых ощущений. Половое возбуждение принимает господствующее положение. Животное стремится к спариванию с самцом, допускает на себя садку и стоит спокойно под ним (половая охота). Половые губы опухают еще более заметно, покраснение слизистых оболочек половых путей усиливается. Маточные железы и клетки шейки матки и влагалища секретируют много слизи жидкой консистенции. Канал шейки матки расслабляется, и у многих животных отмечается вытекание слизи (течка). Матка увеличена, хорошо проявляет ригидность. Быстро созревает фолликул (фолликулы) в яичниках, и в конечном итоге происходит разрыв его и выход яйцеклетки (овуляция).
Метэструс – стадия, следующая за эструсом (послетечковая). Эпителиальные клетки вскрывшегося фолликула (гранулеза) подвергаются гипертрофии и превращаются в лютеиновые; формируется желтое тело. В слизистой оболочке матки разрастаются кровеносные сосуды, маточные железы становятся очень активными. Выделение слизи из влагалища ослабевает, канал шейки матки закрывается. Уменьшается приток крови к наружным половым органам, вульва сморщивается. Прекращается половая охота.
Диэструс – период между следующими один за другим циклами. Характеризуется доминированием циклического желтого тела. Маточные железы активны, мускулатура матки расслаблена, шейка матки закрыта. Количество цервикальной слизи небольшое, консистенция ее вязкая. Слизистая оболочка влагалища бледная.
Анэструс – длительный период полового покоя, в течение которого функция яичников ослаблена. Развитие фолликулов не происходит или слабо выражено. Матка уменьшена и анемична, шейка плотно закрыта. Вагинальная слизь скудная, вязкой консистенции. Слизистая влагалища бледная.
А.П.Студенцов создал учение о половом цикле. В половом цикле он выделяет три стадии.
Стадия возбуждения – характеризуется ярким проявлением основных феноменов (явлений) полового цикла: течки, полового возбуждения, охоты, созревания фолликулов и овуляции. Каждый из этих четырех феноменов является специфическим и отражает одну какую-либо сторону полового цикла. Протекают они во взаимной связи, но возникают и проявляются неодновременно. В стадии возбуждения в половой и других системах организма преобладают пролиферативные процессы.
Течка – процесс выделения слизи из половых органов как следствие морфологических изменений полового аппарата самки. Характеризуется ярко выраженными пролиферативными процессами: гиперемией всех компонентов полового аппарата, новообразованием и разрастанием секретирующих клеток и желез слизистых оболочек яйцепроводов, рогов, тела и шейки матки. Число слоев эпителия во влагалище и преддверии влагалища увеличивается с 3-4 до 18-20. Одновременно отторгаются поверхностные эпителиальные клетки.
Диагностируют течку осмотром наружных половых органов, влагалища, шейки матки, исследованием выделяющейся из половых органов слизи, клиническими и лабораторными методами.
Половое возбуждение (общая реакция) – изменение в поведении самки, возникающее в связи с созреванием фолликулов во время стадии возбуждения. Проявляется половое возбуждение позднее течки и характеризуется ярко выраженной общей реакцией организма в виде беспокойства, отказа от корма, другими признаками. Самка проявляет "интерес" к самцу, может прыгать на него или других самок, но садку самца на себя не допускает.
Охота (половая охота) – положительная сексуальная реакция самки на самца. Во время охоты самка стремится к самцу, принимает позу для полового акта, часто производит акт мочеиспускания, допускает садку и коитус.
Стадия торможения – стадия ослабления и полного прекращения феноменов полового цикла – течки и полового возбуждения; вместо охоты ярко выраженный отбой (самка агрессивно настроена к самцу: стремится его укусить, ударить или убежать от него); на месте овулировавшего фолликула формируется желтое тело. Главная особенность этой стадии - преобладание инволюционных процессов.
Стадия уравновешивания - стадия отсутствия признаков течки, охоты и полового возбуждения. Характеризуется уравновешенным общим состоянием животного, наличием в яичниках и желтых тел, и фолликулов.
В настоящее время в связи с глубоким изучением гормональных изменений в организме и внедрением способов искусственного контроля воспроизводительной функции используют, как правило, терминологию стадий полового цикла по Хиппу.
СОЗРЕВАНИЕ ФОЛЛИКУЛОВ И ОВУЛЯЦИЯ
В период эмбрионального развития в яичниках женских особей закладывается большое количество примордиальных фолликулов. Каждый фолликул состоит из оогония и слоя фолликулярных клеток. Располагаются фолликулы в корковом слое. В яичниках 6-месячного эмбриона человека насчитывается до 6 млн. первичных фолликулов. К моменту рождения число их уменьшается до 200-300 тыс., но к 3 годам из них остается 70%, к 6 годам – 20%. Оставшиеся фолликулы сохраняются до зрелого возраста. После рождения новые ооциты и фолликулы не образуются. У крупного рогатого скота образование первичных фолликулов возможно и в постнатальный период (Г.А.Черемисинов).
В процессе роста фолликул глубже погружается в корковый слой яичника. Рост вначале заключается в увеличении всего фолликула и самой половой клетки. После увеличения в несколько раз оогоний превращается в первичный ооцит. Вокруг него образуется прозрачная оболочка. В это же время происходит рост и увеличение количества плоских эпителиальных (фолликулярных) клеток вокруг ооцита. Клетки эти приобретают кубическую, а затем цилиндрическую форму; количество слоев увеличивается до двух, затем -до трех и более, после чего клетки опять приобретают кубическую форму.
После окончания роста яйца фолликулярные клетки в нескольких местах расходятся и в образовавшихся полостях накапливается жидкость. Постепенно полости объединяются в одну большую полость. Яйцо оттесняется к стенке фолликула. Оно располагается в скоплении эпителиальных клеток, в яйценосном (яйцевом) бугорке (рис. 9). Те клетки, которые непосредственно окружают яйцо, образуют кольцо – лучистый венец. Ободок клеток, выстилающих полость фолликула, называют гранулезой. Содержащаяся в полости фолликулярная жидкость вязкая, богата эстрогенными веществами. В ней формируются структуры, состоящие из кристаллов микроскопической величины, каналов и сферических жировых капель диаметром около 500 мкм. В эструсе эти структуры становятся большими и на типичных кристаллах появляется нитевидный материал,
содержащий ДНК. В стадии диэструс происходит дезорганизация микрокристаллов. Фолликулярная жидкость способствует увеличению и растяжению
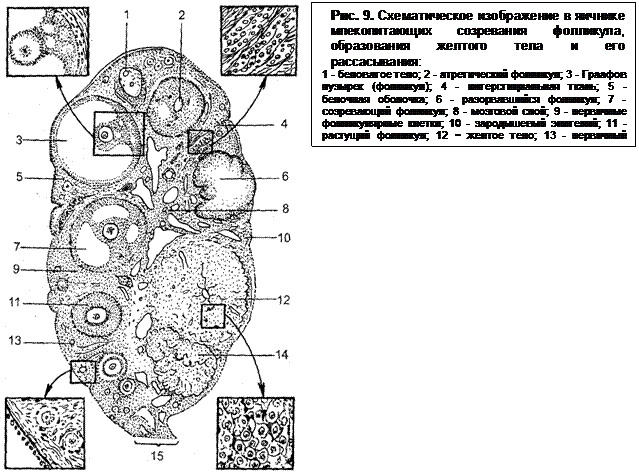 фолликула и выходу его из коркового слоя на поверхность яичника.
фолликула и выходу его из коркового слоя на поверхность яичника.
Гранулезные клетки окружены мембраной. Вокруг нее расположены два слоя клеток, которые образуют оболочку фолликула. Внутренний слой (theca interna) окружает фолликул с момента роста яйца до овуляции. Клетки внутреннего слоя играют большую роль в продуцировании половых гормонов (эстрогенов) и в образовании желтого тела. Их ядра округлые, в отличие от удлиненных, расположенных концентрическими слоями вокруг фолликула, ядер клеток наружной оболочки (theca externa). Состоит наружная оболочка из соединительной ткани и гладких мышечных волокон, обильно снабжена кровеносными сосудами.
Зрелый фолликул питает кровью артериола. У внутреннего полюса фолликула этот сосуд делится на 3-4 ветви, образующие в наружной оболочке широкопетлистую сеть из артериол, а во внутренней – узкопетлистую сеть из капилляров.
По мере роста фолликул становится округлым и набухшим и начинает возвышаться над поверхностью яичника. Внутренняя оболочка фолликула приближается, а затем соприкасается с оболочкой яичника. Обе они истончаются, появляется конусовидный выступ, на верхушке которого образуется разрыв. Яйценосный бугорок теряет связь со стенкой фолликула, и содержимое его, в том числе и яйцеклетка, медленно вытекает через разрыв. В некоторых случаях разрыв сопровождается выбросом части жидкости, а остальная часть ее затем продолжает медленно вытекать. Так происходит овуляция.
Перед овуляцией отмечается первое редукционное деление первичного ооцита и образование первого полярного тельца. Контролируется это, как и овуляция, гонадотропинами, которые стимулируют секрецию фолликулярными клетками особого вещества, специально предназначенного для индукции созревания.
У многих видов овуляция происходит спонтанно, т.е. независимо от наличия или отсутствия полового акта. Главную роль в механизме овуляции играют ЛГ (лютеинизирующий гормон) и окситоцин. Выделение ЛГ и увеличение синтеза простагландина Е приводит к расширению сосудов фолликула, гиперемии, повышению проницаемости капилляров и базальной мембраны. За несколько часов до овуляции преобладает действие простагландина Ф2α; сосуды суживаются, многие капилляры лопаются, соединительнотканные волокна стенки фолликула подвергаются дистрофии. Гранулеза истончается и на выступающей из яичника стенке фолликула перед овуляцией появляется конусовидный выступ.
Раздражение рецепторов полового тракта стимулирует выделение гонадотропных гормонов. На это указывает то, что у некоторых животных (крольчиха, кошка, хомяк) овуляция происходит только после полового акта. У животных со спонтанной овуляцией (у коровы) коитус ускоряет овуляцию. Удаление гипофиза делает овуляцию невозможной.
В процессе разрыва стенки фолликула определенную роль играет повышение давления жидкости в нем, сокращение гладких мышечных волокон яичника и стенки фолликула, а также разрушающее действие фермента гиалуронидазы.
На месте лопнувшего фолликула образуется желтое тело. Это происходит следующим образом. После разрыва стенка фолликула спадается, становится складчатой. В центре каждой складки, состоящей из гранулезы и внутренней оболочки, образуется петля из артериолы. От нее к внутренней оболочке отходит густая капиллярная сеть, через которую происходит васкуляризация желтого тела. Клетки желтого тела (лютеиновые) развиваются в основном из клеток гранулезы. Превращение это сопровождается значительным увеличением их в размере. Они располагаются рядами и имеют многоугольную форму. Между ними размещаются соединительнотканные клетки и кровеносные сосуды. Они образуют перегородки, направляющиеся к центру желтого тела. Благодаря перегородкам желтое тело имеет дольчатое строение. Клетки его продуцируют гормон прогестерон, имеют желтую окраску, что придает всему образованию желтый цвет.
Если в период охоты животное не было осеменено или же после осеменения не оплодотворилось, то желтое тело подвергается обратному развитию (инволюции). В этот период его называют желтым телом полового цикла. При оплодотворении и наступлении беременности у животного желтое тело называют желтым телом беременности.
Развитие фолликулов в яичниках самок происходит постоянно. Однако до наступления половой зрелости ни один из фолликулов не достигает состояния полной зрелости и подвергается обратному развитию без выхода яйцеклетки. Этот процесс называют атрезией фолликулов. Начинается атрезия с деструктивных изменений в ооците или самом фолликуле. В малых фолликулах сначала разрушается ооцит, затем дегенерирует и исчезает весь фолликул. В крупных фолликулах атрезия нередко сопровождается превращением клеток гранулезы в лютеальные и образованием структуры, сходной с желтым телом. У половозрелых животных многие фолликулы на различных стадиях развития также подвергаются атрезии.
НЕПОЛНОЦЕННЫЕ ПОЛОВЫЕ ЦИКЛЫ
Неполноценным называют такой половой цикл, в течение которого выпадает один или несколько феноменов. В связи с этим различают неполноценные циклы: анэстральный (отсутствие течки), алибидный (отсутствие охоты, "тихая овуляция"), ановуляторный (отсутствие овуляции), ареактивный (отсутствие признаков общей реакции). Могут быть смешанные неполноценные половые циклы (ареактивно-ановуляторные и др.).
Особый интерес представляют циклы ановуляторные, так как по внешним проявлениям они в большинстве случаев не отличаются от нормального, сопровождающегося овуляцией. У крупного рогатого скота и лошадей ановуляторные циклы являются одной из важных причин бесплодия. При этих циклах крупные фолликулы подвергаются атрезии, лютеинизации или дегенерации (кистозному перерождению). При атрезии вначале погибает яйцевая клетка, затем разрушается фолликулярный эпителий, сам фолликул уменьшается в размере, приобретает более упругую консистенцию и в течение 3-4 дней полностью исчезает. В процессе лютеинизации происходит постепенное заполнение полости неовулировавшего фолликула клеточными элементами его стенки. Обычно размножаются и разрастаются клетки гранулезы, которые затем превращаются в лютеальные клетки. Лютеинизация начинается от периферии фолликула и распространяется к центру. В разрастающихся клетках откладывается пигмент лютеин. К 5-6-му дню возникает атретическое желтое тело. Дегенерация – это гибель яйцеклетки и последующее увеличение фолликула в диаметре вследствие накопления жидкости и в конечном итоге превращение в кисту. Киста может образоваться уже на 5-7-й день после охоты. При ановуляторных циклах оплодотворение невозможно.
Алибидный половой цикл характеризуется отсутствием охоты у животного при наличии течки и созревания фолликулов и овуляции в яичниках. Особенно часто наблюдается у коров – в 10-30% случаев, а в первые 2 мес. после отела может достигать 65%. У телок первая овуляция, как правило, не сопровождается охотой. Оплодотворение в алибидный цикл возможно.
НЕЙРОГУМОРАЛЬНАЯ РЕГУЛЯЦИЯ ПОЛОВОЙ ФУНКЦИИ САМОК
Проблема обеспечения нормального проявления воспроизводительной функции у самок животных имеет исключительно важное значение. Решение ее в условиях современных технологий животноводства связано не только с созданием оптимальных условий кормления и содержания животных, но и с использованием биотехнологических приемов и способов управления процессами размножения. Эффективность этих способов зависит от того, насколько строго учитывается механизм естественного контроля этих процессов.
ФАКТОРЫ, ОБУСЛОВЛИВАЮЩИЕ ПОЛОВУЮ ЦИКЛИЧНОСТЬ
Регуляция половой функции осуществляется центральной нервной системой (гипоталамусом) во взаимодействии с гормонами гипофиза, половых и других эндокринных желез и рядом биологически Активных веществ.
В нейросекреторных клетках ядер гипоталамуса секретируется гонадотропин-рилизинг-гормон (ГнРГ, гонадолиберин). Это вещество (декапептид) из гипоталамуса перемещается в ножку гипофиза (medium eminence) и через соединения вилкообразных нервных окончаний с капиллярами (вазоневральные синапсы) попадает в кровь и благодаря особой системе кровообращения переносится в переднюю долю гипофиза (рис. 10). В гипофизе гормон связывается с мембраной секретирующей ткани и изменяет ее полярность, вследствие чего возрастает проницаемость для ионов кальция. По микроканалам кальций достигает секреторных гранул и вызывает освобождение их содержимого (гонадотропных гормонов).
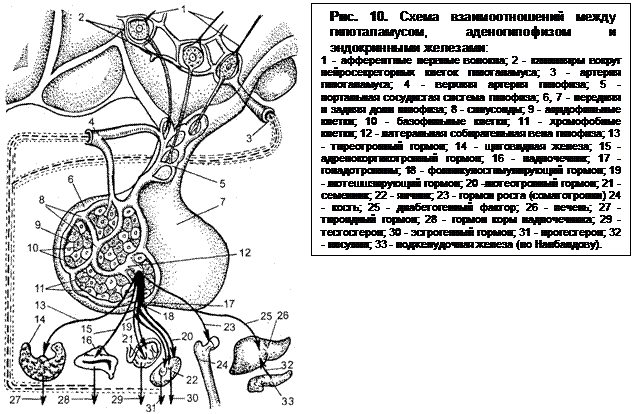 ГнРГ не имеет половой и видовой специфичности, но обладает строгой специфичностью биологического действия, т. е. стимулирует выделение только ЛГ и ФСГ. Он синтезирован.
ГнРГ не имеет половой и видовой специфичности, но обладает строгой специфичностью биологического действия, т. е. стимулирует выделение только ЛГ и ФСГ. Он синтезирован.
В регулировании выделения гонадотропинов принимают участие и другие специфические neurotransmitter субстанции. Норадреналин стимулирует выделение ФСГ и ЛГ; задержка превращения дофамина (dofamine) в норадреналин блокирует эстрадиолиндуцированное выделение ЛГ, которое необходимо для вызова овуляции. Серотонин угнетает базальную секрецию ЛГ и регулирует другие нейросекреторные системы. Дофамин играет также важную роль в контроле выделения пролактина.
Определенная роль в осуществлении контроля ЦНС за деятельностью гипофиза принадлежит и ряду эндогенных опиоидных пептидов, выделяемых мозговой тканью (р-эндорфин, метэнкефалин). Эти вещества обнаружены в высоких концентрациях в гипоталамусе и крови гипофизарной портальной системы в течение полового цикла и в другие фазы репродуктивного периода. Инъекция опиоидных пептидов угнетает секрецию ФСГ и ЛГ, но стимулирует выделение пролактина. Введение же налоксона (naloxone), специфического опиоидного ингибитора, вызывает повышение концентрации и частоты эпизодического выделения го-надотропных гормонов. Очевидно, что опиоидные пептиды влияют на выделение гонадотропинов путем предотвращения выделения ГнРГ из нервных окончаний в ножку гипофиза. Поэтому ГнРГ не попадает в кровь портальной системы и в переднюю долю гипофиза для стимулирования выделения ФСГ и ЛГ.
 У домашних животных секреция ФСГ и ЛГ контролируется двумя функционально различными, но тесно взаимодействующими гипоталамическими системами. Соответственно имеется два гипоталамических центра. Тоническая эпизодическая система обеспечивает продолжительную базальную секрецию гонадотропинов, тем самым стимулирует рост зародышевой и эндокринной ткани яичника. Пулъсаторная система контролирует непродолжительные массивные выделения гонадотропинов, особенно ЛГ, необходимые для овуляции.
У домашних животных секреция ФСГ и ЛГ контролируется двумя функционально различными, но тесно взаимодействующими гипоталамическими системами. Соответственно имеется два гипоталамических центра. Тоническая эпизодическая система обеспечивает продолжительную базальную секрецию гонадотропинов, тем самым стимулирует рост зародышевой и эндокринной ткани яичника. Пулъсаторная система контролирует непродолжительные массивные выделения гонадотропинов, особенно ЛГ, необходимые для овуляции.
В гипоталамусе синтезируется и один из гормонов гипофиза - окситоцин. Этот гормон продвигается вдоль аксонов гипоталамо-гипофизарного тракта в заднюю долю гипофиза (рис. 11) и скапливается в ее клетках — питуицитах, где гранулы окситоцина связываются со специфическим белком. Вытяжку из задней доли гипофиза, богатую окситоцином, называют питуитрином.
Окситоцин вызывает сокращение гладких мышц матки и тем са-
мым ускоряет роды. Быстро разрушается в крови ферментом окситоциназой, но в период родов содержится в повышенных количествах. Расширение влагалища в это время рефлекторно вызывает выделение гормонов. При коитусе окситоцин также выделяется в кровь под влиянием нервных импульсов и усиливает сокращение мускулатуры матки, что способствует продвижению спермиев в половых путях самки.
Чувствительность матки к окситоцину определяется концентрацией эндометриальных рецепторов к этому гормону. Количество же рецепторов зависит от соотношения прогестерона и эстрогенов. После формирования желтого тела и повышения содержания прогестерона количество рецепторов уменьшается, а в период лютеальной регрессии, напротив, возрастает (у овец 500-кратно, Flint et. al., 1992). Введение экзогенного эстрадиола индуцирует преждевременное увеличение рецепторов.
Окситоцнн стимулирует и сокращение миоэпителиальных клеток вымени, тем самым способствуя выведению молока из альвеолярного отдела в молочную цистерну. Кроме того, этот гормон играет роль в регрессии желтого тела у жвачных. Он продуцируется в обнаруживаемых количествах гранулезой фолликулов и желтым телом. Простагландин Ф2α стимулирует секрецию окситоцина желтым телом.
Передняя доля гипофиза сама секретирует гормоны. Базофильные клетки вырабатывают ФСГ и ЛГ, ацидофильные – пролактин. Это гонадотропные гормоны. Они являются гликопротеидами. ФСГ и ЛГ состоят из двух субъединиц, одна из которых общая для обоих гонадотропинов и тиреостимулирующего гормона. Часть, представленная углеводородной цепью, специфична и обусловливает биологическую активность гонадотропина; вторая, представленная полипептидной цепью, ответственна за иммунологические свойства. Пролактин представляет единую полипептидную цепь, в которой 211 аминокислотных остатков.
У самок ФСГ вызывает пролиферацию клеток гранулезы фолликулов в яичниках и способствует развитию их от полостной до преовуляторной стадии. Он активизирует также биохимические процессы в клетках гранулезы и оболочки фолликулов, обеспечивает синтез эстрогенов.
ЛГ стимулирует синтез половых гормонов всеми клетками яичника: желтого тела, гранулезы и оболочки фолликулов и интерстициальными. Определенная базальная концентрация его является обязательным условием для осуществления синтеза гормонов в яичниках. ЛГ вызывает также увеличение кровотока в яичниках и овуляцию фолликулов, подготовленных фолликул остимулирующим гормоном.
Следует отметить, что ФСГ и ЛГ проявляют свое действие в комплексе. Так, ФСГ оказывает слабое действие на рост фолликулов в яичниках, а ЛГ не влияет на их рост, но вместе при строго определенном соотношении эти гормоны обеспечивают полное созревание фолликулов и их овуляцию, а также секрецию эстрогенов. Под влиянием ЛГ-образуется желтое тело и начинается секреция прогестерона. Соотношения этих гормонов в гипофизе определяют особенности течки и охоты у разных видов животных. У кобыл высокое содержание ФСГ и низкое содержание ЛГ, поэтому половая охота у них наиболее продолжительная, а овуляция происходит задолго до окончания охоты. Нередко овуляция задерживается или совсем не происходит. У коров, наоборот, содержание ЛГ высокое, ФСГ – низкое. У них охота менее продолжительна, а нередко овуляция происходит без признаков охоты ("тихая овуляция").
Пролактин (ЛТГ) вместе с АКТГ и соматотропином образует гипофизарный лактогенный комплекс. Как и эстрогены, пролактин стимулирует рост протоков вымени и оказывает синергичное прогестерону действие на дольчато-альвеолярную систему вымени. Вместе с кортикостероидами пролактин вызывает и поддерживает секрецию молока в развитом вымени. ЛГ и пролактин образуют лютеотропный комплекс, необходимый для поддержания структуры и функции желтого тела у лабораторных животных. У коров, овец и свиней лютео-тропное действие оказывает лютеинизирующий гормон.
Гонадотропины различных животных отличаются друг от друга по химическому составу и антигенным свойствам. Но по биологическому действию они не обладают половой или видовой специфичностью.
В плаценте человека и некоторых обезьян образуется вещество, обладающее биологическим действием, сходным с действием ЛГ; выделяется с мочой. Это хорионический (плацентарный) гонадотропин (ХГТ). У женщин его удается обнаружить в моче уже на 9-й день после зачатия. Наибольшее количество его выделяется между 50 и 70-м днями беременности, после чего экскреция резко снижается.
ХГТ стимулирует образование половых гормонов в яичниках и семенниках, а также превращение андро-генов в эстрогены в плаценте, вызывает овуляцию зрелых фолликулов, а у неполовозрелых животных - рост фолликулов и образование желтых тел. Применяется животным для ускорения овуляции при ее задержке или при наличии фолликулярных кист, для синхронизации овуляции после предварительной обработки препаратами прогестерона, при вызове суперовуляции и др. Одна единица ХГТ (1 Е) равна 0,1 мг стандартного вещества.
У беременных кобыл в возвышениях слизистой оболочки матки в месте перехода тела в беременный рог образуется вещество, обладающее гонадотропным действием (ГСЖК). Благодаря большой молекулярной массе оно не выводится почками и циркулирует в крови с 6-й по 20-ю неделю жеребости. Максимальная концентрация его на третьем месяце. Активность гонадотропина измеряется в мышиных (ме) или интернациональных (ИЕ) единицах. 1 ИЕ равна 2,5 ме, или 0,25 мг активного вещества. Мышиной единицей принято называть то наименьшее количество СЖК, которое способно вызвать признаки полового созревания у инфантильного белого мышонка весом 7-8 г.
Биологическое действие гонадотропина СЖК похоже на комбинированное действие ФСГ и ЛГ с преобладанием ФСГ. У неполовозрелых животных ГСЖК вызывает увеличение массы яичников, рост фолликулов, их лютеинизацию, иногда овуляцию и образование желтых тел. Это зависит от используемой дозы. В небольших дозах СЖК вызывает реакцию половых проводящих путей и рост фолликулов (реакция А) и только при значительном увеличении дозы стимулирует образование желтых тел, т. е. реакцию Б (М. М. Завадовский). Широко применяется для вызова суперовуляции у доноров эмбрионов, повышения многоплодия у овец, а также для стимуляции и синхронизации полового цикла у самок.
Половые гормоны – гестагены, андрогены, эстрогены – секретируются половыми железами, надпочечниками, плацентой. Они относятся к группе стероидных гормонов и имеют общую основную структуру. Деление половых гормонов на мужские и женские условно и связано с тем, что яичники вырабатывают в основном прогестерон и эстрогены, а семенники - тестостерон.
Прогестерон - наиболее распространенный гестаген – секретируется желтым телом, в небольших количествах – гранулезой и текой фолликулов перед овуляцией, надпочечниками и плацентой. Этот гормон действует на матку, подготавливает ее к беременности и обеспечивает нормальное ее протекание. Подготовка к беременности начинается- под влиянием эстрогенов. Прогестерон затем превращает эндометрий из проли-феративного в секреторный. Железы эндометрия становятся извитыми, активными и начинают выделять густой секрет, называемый "маточным молоком". Это способствует прикреплению и питанию зародыша. Шейка матки и влагалище выделяют густую слизь, которая закупоривает цервикальный канал. Прогестерон тормозит самопроизвольные ритмические сокращения миометрия, понижает чувствительность его к гормонам, химическим факторам и механическим воздействиям, способствует его гипертрофии. Таким образом обеспечивается растяжение матки растущим плодом.
Прогестерон тормозит развитие фолликулов и овуляцию, а также способствует развитию дольчато-альве-олярной системы вымени.
Прегненолон и прогестерон превращаются в андрогены: дегидроэпиандростерон, андростендион, тестостерон. Образуются они главным образом в семенниках интерстициальными клетками Лейдига и частично клетками Сертоли. Первые два гормона, биологически более слабые, секретируются в значительных количествах надпочечниками и плацентой. Оболочка фолликулов секретирует андростендион и в небольших количествах – тестостерон. Дегидроэпиандростерон и андростендион могут превращаться в коже, печени и других тканях в тестостерон. Таким образом обеспечивается высокая концентрация андрогенов в отдельных тканях без высокой концентрации тестостерона в крови. Это предотвращает его нежелательное общее воздействие на организм самки. Наиболее активный тестостерон в специфических тканях (предстательной железе и др.) превращается в еще более сильный метаболит – дегидротестостерон.

