




Андрогены контролируют сперматогенез, развитие и проявление вторичных половых признаков, а также оказывают анаболическое действие. Контроль за сперматогенезом заключается во влиянии тестостерона совместно с ФСГ на извитые канальцы семенника и образование жизнеспособных сперматозоидов. Развитие и функционирование вторичных половых органов (придатка семенника, спермиопровода, мочеиспускательного канала, придаточных половых желез) почти целиком зависит от тестостерона. Развитие вторичных половых признаков (рост волос, характерные телосложение и развитие голосовых связок, обусловливающее более низкий голос) является в основном следствием анаболических процессов, стимулирующихся андрогенами. Эти процессы приводят к накоплению азота и синтезу белка, формированию мышечной массы. Половое влечение (libido) у самцов и самок и агрессивность у самцов также обусловлены андрогенами.
В эндокринных органах и коже андрогены превращаются в эстрогены. Основные эстрогены: Е: - эстрон, Е2а- 17а - эстрадиол, Е2р- 17р - эстрадиол, ЕЗ - эстриол. Секретируются эстрогены яичниками, главным образом клетками внутренней оболочки и гранулезой, в которой содержатся ферментные системы, обеспечивающие превращение андрогенов в эстрогены. В небольшом количестве эстрогены продуцируются надпочечниками и семенниками (семенники жеребца секрети-руют большое количество гормона).
Главным из эстрогенов является эстрадиол 17„. Он образуется клетками внутренней оболочки фолликула. Здесь вырабатывается и небольшое количество эстрона. Эстрогены и другие стероидные гормоны накапливаются в фолликулярной жидкости. Максимальное количество их установлено между 9-м и 50-м часами перед овуляцией. Из фолликулярной жидкости эстрогены попадают к трубчатым половым органам. Это происходит за 6-30 ч до овуляции.
Эстрогены вызывают васкуляризацию и усиление кровообращения тканей половых органов, разрастание слизистых оболочек матки и влагалища, увеличение секреции слизи клетками влагалища и шейки матки, повышают активность желез матки, способствуют разрастанию миометрия, повышают тонус и усиливают спонтанные сокращения матки. Матка под влиянием эстрогенов становится более устойчивой к инфекции. Эстрогены обусловливают поведение животного, характерное для течки и охоты. Они играют важную роль в наступлении предродовых изменений в половых органах и в начале проявления сократительного потенциала миометрия, стимулируют анаболизм белка и развитие молочных протоков вымени. Чем больше половых циклов у телок к моменту оплодотворения, тем полнее развитие в их вымени молочных протоков и выше молочная продуктивность в первую лактацию.
В крови стероидные гормоны находятся в связанном с белками состоянии. Концентрируются они в специфических тканях: эстрадиол – в цитоплазме и ядрах клеток эндометрия, тестостерон — в предстательной железе. В плазме клеток органов-мишеней имеются специфические рецепторные белки, которые избирательно связываются с гормонами. Например, эстрадиол, имеющий наибольшее сродство с белком эндометрия, связывается с ним сильнее, чем эстрон и эстриол. После взаимодействия гормона с рецепторным белком происходит изменение структуры белковой части комплекса. После этого комплекс гормон - белок проникает в ядро клеток. Взаимодействуя с определенными участками хромосом, гормон активизирует синтез РНК. Одновременно повышается синтез других белков и развиваются процессы, характеризующие биологическое действие гормона на орган-мишень.
В регуляции воспроизводительной функции самок большая роль принадлежит простагландинам. Впервые на существование таких веществ указали гинекологи из Нью-Йорка Курзрок и Лиеб в 1930 г. Они заметили, что матка женщины реагирует сокращением или расслаблением в ответ на свежую сперму. В 1935 г. шведский ученый Эйлер показал, что тонизирующим действием на гладкую мускулатуру обладает липидный экстракт спермы человека, обезьян, овец и коз. Он предполагал, что это вещество вырабатывается в предстательной железе и назвал его простагландином.
Простагландины – это биологически активные липиды, которые содержатся в небольших количествах практически в каждой ткани тела. Биосинтез всех наиболее известных групп простагландинов (А, Б, Е и Ф) происходит из ненасыщенных жирных кислот – арахидоновой и линоленовой. Эти кислоты входят в состав клеточных мембран. Здесь с помощью специфических ферментов и осуществляется синтез простагландинов. Действие каждого из простагландинов порой несходно или даже противоположно. Так, простагландины Ег и Е2 повышают тонус и увеличивают амплитуду сокращений матки и расположенных ближе к ней участков яйцепроводов и одновременно расслабляют дистальный конец их. Тем самым они облегчают доступ спермиев к яйцеклетке и задерживают продвижение ее в полость матки, пока не наступит оплодотворение. ПГ-Е3 угнетает сокращения яйцепроводов и матки, а ПГ-Ф2α стимулирует их. Простагландины группы Ф наиболее широко используются в животноводстве.
У самок ПГ-Ф2α проявляет два важных действия. Во-первых, он усиливает сокращения яйцепроводов и матки, способствуя передвижению спермиев, а также участвует в механизме передвижения зигот. Наряду с окситоцином он играет существенную роль в родовом процессе. В отличие от окситоцина, к которому беременная матка нечувствительна, ПГ-Ф2α вызывает сильные сокращения ее и прерывает беременность, если в этот период накапливается в избытке. Во-вторых, ПГ-Ф2α обладает сильным лютеолитическим действием. Он выделяется эндометрием в значительных количествах в конце полового цикла и обусловливает наступление очередной охоты.
У крупного рогатого скота простагландины попадают из матки в яичник по местному маточно-яичнико-вому пути. Этот путь включает маточно-яичниковую вену и тесно прилегающую к ее стенке сильно извилистую яичниковую артерию. Простагландины из вены проникают в артерию и достигают по ней яичника, связываются с высоко сродственными к ним рецепторны-ми местами в желтом теле и тем самым нарушают стимуляцию лютеинизирующим гормоном аденилциклазы и синтез прогестерона.
У жвачных животных регрессия желтого тела происходит в результате эпизодического выделения ПГ-Ф2α из матки с интервалом около 6 ч. Выделение стимулируется окситоцином, секретируемым желтым телом. Поэтому каждое эпизодическое выделение про-стагландина связано с эпизодическим выделением окситоцина. Более того, простагландин стимулирует секрецию окситоцина яичником.
МЕХАНИЗМ ЕСТЕСТВЕННОГО РЕГУЛИРОВАНИЯ ПОЛОВОГО ЦИКЛА
Основная роль в механизме естественного регулирования воспроизводительной функции отводится центральной нервной системе (гипоталамусу), гормонам аденогипофиза и половых желез, простагландинам и ряду гонадальных факторов.
На центральную нервную систему оказывают влияние внешние (свет, температура, присутствие самца и др.) и внутренние (гормоны яичника) факторы (рис. 12). Они стимулируют или тормозят активность гипоталамуса. В цитоплазме нервных клеток ядер гипоталамуса секретируется гонадатропин-рилизинг-гормон. Под влиянием адекватных раздражителей ГнРГ из гипоталамуса через портальную систему кровообращения попадает в переднюю долю гипофиза и вызывает выделение ФСГ и ЛГ.
Эти гормоны, воздействуя на яичник, стимулируют рост и созревание фолликулов и секрецию эстрогенов. Примерно между 16-м и 19-м днями полового цикла у коров, на 12-14-й день у кобыл и на 15-й день у овец и свиней отмечается резкое уменьшение содержания прогестерона в периферической крови и увеличение эстрогенов. У коров и овец содержание эстрогенов достигает пика в начале стадии эструса и затем снижается до базального уровня к моменту овуляции; у кобыл пик гормона отмечается в течение эструса, а у свиней за 48 ч до начала эструса.
Эстрогены усиливают кровообращение и рост тканей наружных половых органов и влагалища, стимулируют секрецию слизи клетками влагалища и шейки матки (вызывают течку). Под их влиянием увеличивается активность миометрия и происходят изменения в эндометрии, которые являются началом подготовки его к приему зародыша. В промежуточном мозгу возникает очаг возбуждения. При участии гипоталамических центров и лимбической системы формируется половая мотивация, а при участии центра в коре больших полушарий — половая доминанта, т.е. состояние половой охоты.
 В это время созревающие фолликулы (у крупного рогатого скота – доминантный фолликул) продуцируют возрастающие количества ингибина, фолликулярного регулирующего протеина (ФРП) и эстрадиола. Ингибин и эстрадиол, воздействуя на гипофиз, способствуют понижению секреции ФСГ и таким образом блокируют появление новых крупных фолликулов, а ФРП из преовуляторных фолликулов угнетает синтез эстрадиола второстепенными фолликулами, что приводит к их атрезии.
В это время созревающие фолликулы (у крупного рогатого скота – доминантный фолликул) продуцируют возрастающие количества ингибина, фолликулярного регулирующего протеина (ФРП) и эстрадиола. Ингибин и эстрадиол, воздействуя на гипофиз, способствуют понижению секреции ФСГ и таким образом блокируют появление новых крупных фолликулов, а ФРП из преовуляторных фолликулов угнетает синтез эстрадиола второстепенными фолликулами, что приводит к их атрезии.
Следует отметить, что ингибин секретируется всеми полостными фолликулами и имеет более продолжительный период полураспада, чем эстрадиол, который продуцируется только преовуляторными фолликулами. Поэтому действие ингибина на выделение ФСГ более заметное, чем эстрадиола. В то же время резкое увеличение содержания эстрадиола в преовуляторных фолликулах обусловливает повышение чувствительности их к гонадотропинам крови и обеспечивает дальнейшее развитие до овуляции. На выделение ФСГ оказывают влияние и два других пептидных гормона, которые выделены из фолликулярной жидкости – активин (стимулирует выделение) и фоллистатин (угнетает секрецию гонадотропина).
У овец не ингибин, а эстрадиол является основным фактором в механизме регулирования уровня гонадотропинов в периферической крови. Резкое увеличение эстрогенов (эстрадиола) в начале эструса приводит к увеличению содержания ЛГ, пик которого наблюдается примерно за 14 ч до овуляции. В соответствии с этим отмечается и пик ФСГ, а второе его повышение наблюдается через 2 дня после овуляции.
Повышение содержания эстрадиола активизирует гипоталамус, который стимулирует выделение из передней доли гипофиза большого (овуляторного) количества ЛГ. У кобыл содержание ЛГ повышается постепенно и удерживается на относительно высоком уровне в течение 5-6 дней до и после овуляции. В ответ на резкое возрастание количества ЛГ в плазме усиливается рост клеток гранулезы фолликулов, начинается секреция прогестерона и высвобождение коллагенолитического фермента, истончающего стенку фолликула. Это приводит к разрыву ее под влиянием гидростатического давления фолликулярной жидкости. Обычно овуляция происходит спустя 12 ч после окончания охоты у коров, а у овец, свиней и кобыл до окончания охоты примерно за 6, 10-15 и 24 ч соответственно.
На месте овулировавшего фолликула образуется желтое тело, которое секретирует прогестерон. Быстрое увеличение содержания этого гормона вскоре после охоты угнетает секрецию ГнРГ и препятствует дальнейшему освобождению ЛГ. Овуляция новых фолликулов не происходит. Под влиянием прогестерона эндометрий полностью подготавливается к приему и прикреплению зиготы и обеспечивает нормальное протекание беременности. Выделение прогестерона желтым телом поддерживается у ряда лабораторных животных лютеотропным гормоном, а у коров, овец и свиней - ЛГ.
При отсутствии в матке зародыша эндометрий выделяет в конце полового цикла лютеолитический фактор (ПГ-Ф2α), который, действуя на яичник, вызывает регрессию желтого тела. Исчезновение основного источника прогестерона происходит на фоне наличия в яичниках новых растущих фолликулов. Рост их начинается сразу же после овуляции вследствие уменьшения уровня ингибина, ФРП и эстрадиола и последующего периодического увеличения ФСГ. У крупного рогатого скота первая после овуляции волна роста фолликулов заканчивается атрезией вначале мелких, а затем и самого крупного из них (доминантного) фолликула. Атрезия, в свою очередь, дает начало новой волне роста фолликулов. У этого вида животных в течение полового цикла имеют место две или чаще три таких волны роста фолликулов. В результате последней волны появляется доминантный овуляторный фолликул и происходит овуляция, завершается предыдущий и начинается новый половой цикл, а в случае успешного осеменения животного развивается беременность. У беременных животных лютеолитический эффект эндометрия устраняется присутствием эмбриона.
 ПОЛОВОЙ ЦИКЛ КОРОВЫ
ПОЛОВОЙ ЦИКЛ КОРОВЫ
Половая зрелость у телок наступает чаще в возрасте 8-10 мес. Осеменяют их в 16-18-месячном возрасте при достижении живой массы 350 кг. По экономическим соображениям целесообразно более раннее осеменение (с 13-15 мес). При этом развитие телок должно быть хорошим, а последующее кормление во время беременности обеспечить достаточный прирост и достижение безопасной для первого отела живой массы.
Коровы - полициклические животные. После наступления первой овуляции у телок половая цикличность проявляется более или менее регулярно во все сезоны года. Продолжительность полового цикла у них составляет в среднем 20 (от 18 до 22) дней. В случае недокорма половая цикличность прекращается. Чаще это можно наблюдать осенью и зимой в холодную погоду.
У коров после родов с 7-10-го дня отмечается рост фолликулов в яичниках, и в конце второй недели может произойти овуляция. В среднем интервал от отела до первой овуляции составляет 20-38 дней (от 14 до 40 дней). У многих коров первая послеродовая волна роста фолликулов не завершается овуляцией. Но она сопровождается кратковременным повышением в крови (молоке) содержания прогестерона (рис. 13). Нередко отмечается два или даже три таких подъема гормона. Погрешности в кормлении и содержании, частое доение и подсос, а также различные заболевания в послеродовой период тормозят проявление охоты и овуляции. Первый половой цикл короче, чем второй, что связано с недостаточной функциональной активностью первого желтого тела. Наличие в матке инфекции может укорачивать цикл до 10 дней. В среднем длина полового цикла у коров составляет 21 день. Нормальные колебания находятся в пределах 18-24 дней. Длительность стадий полового цикла: проэструса – 3-4 дня, эструса – 12-18 ч, метэструса – 3-4 и диэструса – 10-14 дней.
Половая охота у коров и телок продолжается в среднем 16 ч (от 10 до 23 ч), у большинства животных она длится 13-17 ч (В.С.Шипилов). Возможны колебания продолжительности охоты от 2,5 до 36 ч. У телок с возрастом половая охота удлиняется и признаки ее у них выражены более отчетливо, чем у коров. Продолжительность первой послеродовой охоты короче, чем двух последующих. В жаркие месяцы у лактирующих коров отмечается значительное укорочение охоты. У многих животных она проявляется утром или в вечернее время.
Нередко у телок первая овуляция не сопровождается внешними признаками половой охоты ("тихая овуляция"). У коров в первый послеродовой цикл частота "тихой овуляции" достигает 70% и более. После установления регулярных циклов у телок и в последующие циклы у коров частота их с хорошо выраженной охотой повышается. У животных с "тихой овуляцией" отмечаются обычные морфологические и функциональные изменения в половых органах, но характерные для половой охоты поведенческие реакции отсутствуют. Тщательным наблюдением таких животных можно распознать.
На протяжении полового цикла в яичниках коров наблюдается две или три волны роста фолликулов (рис. 14). Поэтому в них постоянно находятся фолликулы диаметром около 0,5 см. В течение охоты доминантный фолликул быстро увеличивается в диаметре до 1,5-2 см и свободно пальпируется через прямую кишку. Окончательное созревание его происходит после охоты. У телок крупные фолликулы часто образуют сферическую выпуклость над поверхностью яичника. У коров они обычно не выпячиваются и дают ощущение большого диаметра, не вызывая больших изменений в очертаниях яичника. В момент овуляции диаметр фолликула составляет 16-19 мм, но некоторые фолликулы, имеющие меньший диаметр, также могут разрываться. Овуляция может происходить в любом месте свободной поверхности яичника. В последующем в связи с развитием желтого тела форма яичника будет зависеть в основном от места овуляции.
У 4-5 % коров овулирует два фолликула. Около 60 % всех овуляций происходит в правом яичнике. Овуляция наступает через 12,5 ± 0,8 ч после окончания охоты или через 28,9 ± 0,7 ч после ее начала (B.C. Шипилов). У коров время овуляции может колебаться от 2 ч до окончания охоты до 26 ч после окончания (в среднем через 12,5 ч), у телок - от 2,2 до 22 ч после окончания, в среднем через 11 ч (Г. У. Солсбери, Н. Л. ВанДемарк). В 85% случаев овуляция происходит в вечерне-ночное время; наличие самца и акта спаривания ускоряет ее.

 После разрыва фолликула полость его уменьшается, гранулеза и внутренний слой оболочки образуют складки и почти целиком заполняют полость. В ней остается лишь небольшое количество фолликулярной жидкости. Кровяных сгустков не бывает, но на месте разрыва иногда появляется небольшое кровоизлияние. Клетки гранулезы превращаются в лютеальные клетки, и через четыре дня после овуляции формирование желтого тела в основном завершается, секреция прогестерона резко возрастает. К этому времени оно имеет желтовато-коричневый цвет и его можно пальпировать через прямую кишку. Покровный эпителий полностью закрывает овуляционную ямку. К 10-му дню заканчивается васкуляризация желтого тела, оно окончательно сформировано, становится оранжевым или желтым. Диаметр его достигает 2-2,5 см, масса - 4,1-7,4 г. В период с 11-го по 17-й день цвет варьирует от светло-оранжевого до светло-желтого. В 16-25% случаев желтое тело внутри имеет небольшую полость, в которой содержится желтоватая жидкость (рис. 15, 16).
После разрыва фолликула полость его уменьшается, гранулеза и внутренний слой оболочки образуют складки и почти целиком заполняют полость. В ней остается лишь небольшое количество фолликулярной жидкости. Кровяных сгустков не бывает, но на месте разрыва иногда появляется небольшое кровоизлияние. Клетки гранулезы превращаются в лютеальные клетки, и через четыре дня после овуляции формирование желтого тела в основном завершается, секреция прогестерона резко возрастает. К этому времени оно имеет желтовато-коричневый цвет и его можно пальпировать через прямую кишку. Покровный эпителий полностью закрывает овуляционную ямку. К 10-му дню заканчивается васкуляризация желтого тела, оно окончательно сформировано, становится оранжевым или желтым. Диаметр его достигает 2-2,5 см, масса - 4,1-7,4 г. В период с 11-го по 17-й день цвет варьирует от светло-оранжевого до светло-желтого. В 16-25% случаев желтое тело внутри имеет небольшую полость, в которой содержится желтоватая жидкость (рис. 15, 16).
По мере формирования желтое тело выпячивается из яичника. У телок оно часто сильно выступает из стромы яичника и прикреплено к ней только основанием. У коров желтое тело нередко полностью погружено в ткань яичника, и его положение можно обнаружить по выступающей овуляционной точке. У большинства желтых тел к моменту их максимального роста эта точка расположена в виде углубления в возвышении диаметром около 1 см. Некоторые желтые тела возвышаются в виде сосочка, но иногда место возвышения их диффузно разлито (рис. 16).
Наличие желтого тела в яичнике изменяет его размеры. Как правило, этот яичник значительно больше по величине, чем другой. Хорошо заметна эта разница у телок.
 Уровень секреции прогестерона желтым телом достигает максимума через 7-8 дней после овуляции и быстро понижается с 18-го дня (см. рис. 13, 14). За 24-30 ч до следующей охоты желтое тело, как правило, начинает быстро уменьшаться в величине, цвет его становится желтовато-оранжевым. В период охоты диаметр желтого тела уменьшается до 1,5 см, а к середине полового цикла составляет около 0,5 см, и выступающая часть его немного больше булавочной головки.
Уровень секреции прогестерона желтым телом достигает максимума через 7-8 дней после овуляции и быстро понижается с 18-го дня (см. рис. 13, 14). За 24-30 ч до следующей охоты желтое тело, как правило, начинает быстро уменьшаться в величине, цвет его становится желтовато-оранжевым. В период охоты диаметр желтого тела уменьшается до 1,5 см, а к середине полового цикла составляет около 0,5 см, и выступающая часть его немного больше булавочной головки.
 В течение полового цикла заметным изменениям подвергаются эпителиальные клетки передней части влагалища и секретирующие клетки шейки матки. В середине цикла клетки влагалища варьируют от уплощенных до низкоцилиндрических. В стадии проэструса происходит размножение и рост высоких цилиндрических слизеобразующих поверхностных клеток. В эструс этот процесс усиливается, что приводит к сильному утолщению эпителия передней части влагалища.
В течение полового цикла заметным изменениям подвергаются эпителиальные клетки передней части влагалища и секретирующие клетки шейки матки. В середине цикла клетки влагалища варьируют от уплощенных до низкоцилиндрических. В стадии проэструса происходит размножение и рост высоких цилиндрических слизеобразующих поверхностных клеток. В эструс этот процесс усиливается, что приводит к сильному утолщению эпителия передней части влагалища.
За 12-24 ч до появления первых признаков охоты начинается обильная секреция слизи из передней части влагалища и шейки матки. Количество слизи увеличивается в течение охоты и затем постепенно уменьшается к 4-му дню после охоты. В это время слизь содержит хлопья из лейкоцитов. Лейкоцитарная инвазия слизистой оболочки влагалища максимальная через 2-5 дней после охоты. Эластичность слизи варьирует, она максимальная во время охоты. В связи с эластичностью изменяется и способность слизи к кристаллизации. Наиболее выражена кристаллизация во время охоты и в течение 2 последующих дней, а в другое время отсутствует. Под микроскопом в мазках кристаллы слизи расположены в виде листка папоротника. Этот феномен (арборизация), так же как количество и свойства слизи, зависит от уровня эстрогенов. В начале охоты слизь клейкая, прозрачная, светлая, а к концу охоты становится более вязкой. Время наименьшей вязкости слизи совпадает с временем максимального ослизнения слизистой оболочки преддверия влагалища и минимального электрического сопротивления покрывающей эту оболочку слизи. Наряду с изменением внешних свойств изменяется структура и химический состав, а также рН слизи. В передней части влагалища за день до охоты показатель рН составляет 6,7-7,0, но затем уменьшается в начале охоты и к моменту овуляции опускается до минимального значения (6,5-6,6), затем быстро увеличивается до 7,0 и достигает 7,2-7,4 на 4-5-й день и 7,3-7,5 на 11-12-й день.
Циклические изменения в матке и во влагалище контролируются гормонами. Под влиянием эстрогенов в период течки и охоты прогрессирующе усиливается кровообращение половых органов, увеличивается сеть кровеносных сосудов, т. е. происходит васкуляризация. Матка кровенаполнёна, ее слизистая оболочка блестящая и покрыта отечной жидкостью. После овуляции уровень эстрогенов быстро падает и оказывается недостаточным, чтобы поддержать такое состояние сосудов. Наполненные кровью, они разрываются (главным образом в области карункулов), и небольшое количество крови выходит в матку. Кровь смешивается со слизью и выделяется из половых органов. У некоторых животных через 40-48 ч после начала охоты наблюдается выделение ярко-красной слизи. Спустя 3-5 дней после окончания охоты резко снижается интенсивность кровообращения в половых органах, слизистая оболочка влагалища и матки становится бледной, канал шейки матки закрыт, слизи немного, она вязкая, светло-желтая или коричневатая.
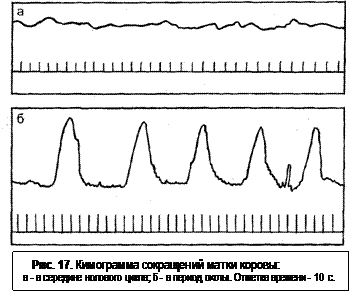 С началом созревания фолликула мышечные волокна матки растут в длину. Матка становится большей по величине. Происходят изменения и в характере ее сокращений. В стадии эструса сокращения с большей амплитудой, не частые (рис. 17). В ответ на пальпацию через прямую кишку матка реагирует сильным сокращением. Способность к сокращениям (ригидность) повышена за день до охоты, в период охоты и на следующий день после охоты; в середине цикла сокращения более слабые и частые.
С началом созревания фолликула мышечные волокна матки растут в длину. Матка становится большей по величине. Происходят изменения и в характере ее сокращений. В стадии эструса сокращения с большей амплитудой, не частые (рис. 17). В ответ на пальпацию через прямую кишку матка реагирует сильным сокращением. Способность к сокращениям (ригидность) повышена за день до охоты, в период охоты и на следующий день после охоты; в середине цикла сокращения более слабые и частые.
Половая охота - строго специфическая реакция самки на самца и ее можно точно установить только рефлексологическим методом, т. е. используя самца-пробника. Однако у коров и телок при контакте с другими самками проявляются такие характерные для охоты признаки, которые позволяют практически безошибочно распознать ее. Знание этих признаков и умение их использовать позволяют животноводам успешно организовать искусственное осеменение коров и телок без использования быков-пробников. Наиболее важными являются следующие признаки.
1. Еще до начала охоты корова обнюхивает и облизывает соседних коров, особенно их половые органы, а также подошедших близко людей; при этом спина спереди от.таза выгибается, а затем сильно прогибается, хвост поднимается и опускается. Такое движение можно вызвать поглаживанием поясницы или дотра-гиванием до вульвы. Этот признак выражен у большинства животных. Иногда корова сильно изгибается и пытается облизать свои половые органы.
2. Слизистая оболочка преддверия влагалища припухшая и гиперемирована, влажная и покрыта прозрачным секретом, который часто вытекает из половой щели, поэтому пучок волос в нижнем углу вульвы всегда бывает влажным.
3. Постоянно переступает с ноги на ногу, а находясь в группе коров, пытается приблизиться к другим животным в охоте (или не в охоте) и вспрыгнуть на них, иногда даже на человека.
4. Корова в охоте проявляет признаки общего возбуждения, чаще стоит. Если кто-нибудь проходит сзади, она поднимает голову и провожает проходящего взглядом.
5. Глаза более живые, часто неподвижные и блестящие зрачки расширены.
6 у некоторых коров охота сопровождается громким
мычанием в течение нескольких часов или всего дня.
7 У многих животных наблюдается снижение удоя, но оно может пройти незамеченным; более обычным является затрудненное доение, так как корова задерживает молоко. Снижается также руминация.
Если корова находится в стаде, признаки охоты выпажены более отчетливо. Животные в охоте в 2-3 раза увеличивают двигательную активность (регистрируемую с помощью педометров), следуют за другими животными и вспрыгивают на них, независимо от того, в охоте эти животные или нет. Наибольшая активность прыгать проявляется в конце проэструса, в эструс и в начале метэструса. Если замечено, что корова повторяет прыжки, следует продолжить наблюдение за ней, так как возможно наступление охоты. Однако этот признак не является определяющим. Он может наблюдаться и у животных, не находящихся в охоте.
8 Более важным для выявления состояния охоты является поведение коровы в момент, когда на нее вспрыгивает другое животное: если она стоит спокойно или слегка уклоняется, это указывает на наличие у нее охоты. В разгар охоты у коров проявляется, как правило, только такой типичный признак (рефлекс неподвижности) и лишь изредка - активный обнимателъный рефлекс, но без совокупительных движении. У животных в это время хорошо выражены и специфические изменения в наружных половых органах. Слизь вытекает в виде длинной вязкой нити, приклеивается к ягодицам и хвосту. Если животное лежало, возле его таза можно увидеть скопление мутноватой или голу6оватого цвета слизи. У телок наблюдается и четкий подъем температуры во влагалище.
У животных с "тихой" охотой эти изменения также имеют место. Отмечается у них и возбужденное состояние, рев, попытки вспрыгивать на других коров. Но сами они не допускают садки других животных и не являются особо "привлекательными". При осеменении таких животных может произойти оплодотворение. При контакте.коровы в охоте с быком животные
облизывают друг друга, после чего корова пытается вспрыгнуть на быка прежде, чем допускает садку сама. Если корова в охоте долгое время находилась в стаде, то вследствие вспрыгивания других животных у нее стирается волос и появляются ссадины на коже крестца. Эти признаки сохраняются 4-6 дней. Но если ссадины влажные, с выделением экссудата или кожа при дотрагивании болезненна и имеет повышенную температуру, это говорит о том, что корова в охоте или охота только что прошла.
Иногда коровы в охоте следуют за другими коровами, с фырканьем обнюхивают их половые органы, опираются головой на поясницу, поднимают хвост и помахивают им. Нередко за такими коровами активно следуют другие животные.
Наблюдается повторяемость особенностей проявления и длительности половых циклов у одного и того же животного в различные годы.
Охоту у коров выявляют на основании проявления у них специфических признаков. Целесообразно трехкратное в день наблюдение по 20-30 мин до раздачи корма во время движения животных. При беспривязном содержании животные должны быть помечены бирками или ошейниками с хорошо заметными номерами. На крупных фермах наблюдение можно проводить с помощью телеустановок.
С целью более полного и своевременного выявления охоты у коров и определения оптимального времени их осеменения используют быков-пробников, а также специально подготовленных коров-выявительниц или натренированных собак, детекторы охоты различных конструкций, электрометрические приборы и т. д.
ПОЛОВОЙ ЦИКЛ ОВЦЫ
Половая зрелость у ярочек наступает в возрасте 5-8 мес. Осеменяют их на втором году жизни. При хорошем развитии самки возможно и более раннее покрытие.
Овцы — полициклические животные с выраженным половым сезоном. У диких овец половой сезон совпадает с осенними месяцами, у одомашненных пород продолжительность его колеблется в зависимости от климатических условий, кормления и содержания, породных особенностей животных. У тонкорунных овец, разводимых на юге Украины и Северном Кавказе, половая цикличность проявляется со второй половины сентября по март. Но, если в июле-августе выпадает достаточно осадков, а температура воздуха невысокая, то взрослые овцы начинают приходить в охоту со второй декады августа, а переярки и полутарогодовалые ярки – с конца июля. В Беларуси овцы могут проявлять половую цикличность в период с июля до марта.
Сезоннность в размножении овец регулируется факторами внешней среды и, главным образом, суточным количеством света. Начинается половой сезон с уменьшением светового дня и продолжается в течение всего короткого дня. Уменьшение света стимулирующим образом действует на гипофиз, повышая его гонадот-ропную активность. Это действие осуществляется через гормон эпифиза - мелатонин.
После родов охота может наступить в первый месяц, но это зависит от сезона ягнения. После весеннего ягнения охота наступает медленнее и значительно быстрее - после осеннего. Повышение содержания прогестерона в организме регистрируется после 50-го дня.
Длительность стадий полового цикла: проэструса – 2-3 дня, эструса – 24-38 ч, метэструса – 2-3 дня и диэструса – 10-12 дней.
Половая охота у овец проявляется менее ярко, чем у других видов, и определить ее у большинства животных без барана-пробника практически невозможно. Сопровождается охота выделением специфических пахучих веществ – аттрактантов (феромонов), которые легко воспринимаются самцами. Выключение зрительной рецепции не дезориентирует баранов: руководствуясь обонянием, бараны и в таких случаях безошибочно находят овец в состоянии охоты. Продолжительность охоты 24-38 ч (от 12 до 72 ч). У старых маток охота длится меньше, чем у молодых. В начале полового сезона охота короче, чем в разгар сезона. В прохладные месяцы охота продолжается дольше, чем в теплые.

