Характеристика яйцеклетки. Яйцеклетки млекопитающих неподвижны и имеют значительно большие размеры, чем спермин. Это обусловлено и большей массой цитоплазмы, и, главным образом, наличием запасного питательного материала в виде желтка (дейтоплазмы). Этот материал представлен желточными гранулами и пластинками. Состоит из белков, фосфолипидов и нейтральных жиров. Желток может находиться в цитоплазме яйцеклетки или быть обособленным от цитоплазмы. Образуется в печени и других органах и через кровь поступает в ооцит.
У домашних животных диаметр яйцеклетки без прозрачной оболочки колеблется от 0,11 до 0,15 мкм и мало зависит от размеров тела. Яйцеклетка коровы имеет диаметр около 115 мкм, толщина прозрачной оболочки 15-30 мкм и общий диаметр клетки составляет 165 мкм. Яйцеклетка овцы несколько больше – соответственно 112-125, 9-16 и 167 мкм. Яйцеклетка свиньи имеет диаметр 130 мкм, толщина прозрачной оболочки – 15 мкм.
Яйцеклетка, вышедшая из фолликула, вскоре погибает, если не будет оплодотворена. Уже в период прохождения истмической части яйцепровода цитоплазма и ядро клетки распадаются на отдельные фрагменты и после размягчения прозрачной оболочки постепенно рассасываются. Происходит это уже в матке, куда неоплодотворенные яйцеклетки попадают примерно в то же время, когда и оплодотворенные. У кобыл неоплодотворенные клетки остаются в яйцепроводах, где подвергаются медленной (в течение нескольких недель или даже месяцев) цитоплазматической дегенерации. У этого вида животных развивающийся зародыш оказывает, по-видимому, более мощное эндокринное (или иное) влияние на функцию яйцепроводов, что позволяет ему проникнуть в матку, тогда как неоплодотворен-ная клетка туда не проходит. Продолжительность жизни яйцеклетки коровы составляет 10-12 ч; овцы – 12-15; лошади – 8-10 ч; свиньи – 6-15 ч. В течение этого периода яйцеклетка может быть оплодотворена, но чаще способность к оплодотворению сохраняется более короткое время и обычно не превышает 6 ч.
По мнению Ф.И.Осташко и др., яйцеклетки млекопитающих сохраняют способность к оплодотворению in vivo 4-5 сут и оплодотворение их возможно не только в яйцепроводе, но и в роге матки.
Сущность оплодотворения. Оплодотворение - физиологический процесс, в результате которого происходит объединение генетического материала яйцеклетки и спермия и образование новой клетки (зиготы) с диплоидным набором хромосом.
Во время продвижения яйцеклетки по яйцепроводу в начале ампулярной части его вследствие беспорядочного движения спермиев происходит встреча их с яйцеклеткой и оплодотворение. Наступление оплодотворения зависит от числа присутствующих спермиев, а также биологической полноценности обеих гамет.
У некоторых млекопитающих яйцеклетка окружена фолликулярными клетками (лучистым венцом). Эти клетки склеены между собой материалом, имеющим в своем составе гиалуроновую кислоту. Под влиянием фермента гиалуронидазы фолликулярные клетки довольно быстро рассеиваются уже в воронке яйцепровода. Фермент деполимеризует гиалуроновую кислоту и разрушает склеивающий клетки материал. Гиалуронидаза содержится в плазме спермы и спермиях (акросоме). Особенно много содержится фермента в семенниках и сперме кролика. Функция гиалуронидазы при оплодотворении заключается в том, что она позволяет отдельным спермиям пройти между фолликулярными клетками и достичь яйца. Для этого не требуется разрушение всего лучистого венца, хотя полное обнажение клетки важно и необходимо для продвижения ее по яйцепроводу, но оно может произойти и без Участия спермиев.
Следует отметить, что этот процесс не имеет видовой специфичности. Это послужило поводом использовать фермент гиалуронидазу для повышения оплодотворяемости путем добавления ее в сперму. Однако результаты испытаний противоречивы.
Лучистый венец хорошо выражен у яйцеклеток свиней, значительно слабее - у коров. Клетки венца могут направлять спермий, когда он проходит между ними, и подводить его к яйцеклетке. Определенную роль в соединении гамет играют гормоны оплодотворения (гамоны). Известно четыре гамона. Два из них (гинога-моны) выделены из яйцеклетки. Гиногамон 1 активизирует действие спермиев, гинагамон 2 вызывает их агглютинацию. Два других (андрогамоны) выделены из спермиев. Андрогамон 1 парализует движение спермиев и нейтрализует гиногамон 1. Андрогамон 2 (акрозин) растворяет прозрачную оболочку и облегчает достижение спермиев поверхности яйцеклетки; он нейтрализует гиногамон 2.
После достижения спермием прозрачной оболочки следует видоспецифическая реакция, которая приводит к прочной связи половых клеток. Сродство определенных фракций белков из плазматических мембран спермиев и прозрачной оболочки яйца, по-видимому, имеет решающее значение в прикреплении к ней. Вскоре после этого спермий по узкому извилистому пути быстро проникает через эту оболочку. При этом отделяется и утрачивается на поверхности прозрачной оболочки пузырчатая акросомная и плазматическая мембраны спермия. Головка спермия попадает в околожелточное пространство и вскоре соприкасается с желточной оболочкой. После этого прозрачная оболочка становится малочувствительной к другим спермиям. Вот почему у ряда видов (хомяк, собака, овца, лошадь и др.) число проникших через прозрачную оболочку спермиев не велико. У других животных (крольчиха, корова, свинья и др.) ответная реакция прозрачной оболочки замедленная или совсем не проявляется и в околожелточное пространство могут попасть десятки и даже сотни спермиев.
После контакта спермия с желточной оболочкой яйцеклетки, когда произойдет слияние их плазматических мембран (у ряда видов под желточной оболочкой находится вторая оболочка – плазматическая мембрана), начинается кортикальная реакция. Кортикальные зерна, расположенные в поверхностном слое цитоплазмы яйцеклетки, вздуваются, затем их оболочки вместе с плазматической мембраной гаметы разрываются. В пространство между ее поверхностью и желточной оболочкой изливается содержимое этих гранул (зерен), которое содержит протеолитический фермент. Он разрушает связи желточной оболочки с другими спермиями, и это предотвращает полиспермию. Кортикальная реакция распространяется по поверхности яйцеклетки волнообразно в течение 10-20 с. У некоторых видов в процессе кортикальной реакции желточная оболочка приподнимается над поверхностью яйцеклетки, образуя перивителиновое пространство. На внутренней поверхности оболочки накапливаются тонким слоем вещества, освобождающиеся из гранул. В результате желточная оболочка утолщается и ее называют оболочкой оплодотворения.
После соприкосновения спермия с желточной оболочкой происходит образование второго полярного тельца и ооцит второго порядка превращается в зрелую яйцеклетку. Затем спермий полностью погружается в цитоплазму клетки. Входит он не перпендикулярно, а по касательной по отношению к желточной оболочке (рис. 58). После вхождения в яйцеклетку головка спермия набухает, теряет форму и оболочку и становится едва заметной, ядерный материал выходит из нее и вскоре появляются в виде мелких точек ядрышки. Они постепенно соединяются и образуют большие, круглые частицы, которые включаются в мужской пронуклеус; пронуклеус окружен ядерной оболочкой.
Также происходит образование женского пронуклеуса. После второго редукционного деления ядро яйцеклетки разрывается и его содержимое смешивается с цитоплазмой. Материал хромосом превращается в раздробленную зернистую массу. Затем образуются ядрышки, которые вскоре окружаются оболочкой. Образуется женский пронуклеус. Эти изменения наступают в течение 2-3 ч после проникновения спермия.
Слияние пронуклеусов (сингамия) является конечной стадией оплодотворения. После сближения в цент-Ре клетки пронуклеусы уменьшаются- в величине, исчезают ядрышки; затем исчезают и ядерные оболочки. Появляются две группы хромосом, которые движутся навстречу друг другу и в стадии метафазы происходит их слияние (соединение ядер клеток при конъюгации). Почти непосредственно за этим следуют анафаза и телофаза первого деления мейоза.
В отдельных случаях в яйцеклетке оказывается 2 или даже 3 спермия (полиспермия). В результате образуется 3 или 4 пронуклеуса. Возможность оплодотворения несколькими спермиями может возникать в тех случаях, когда второй спермий проникает через прозрачную оболочку в точке, диаметрально противоположной точке первоначального проникновения. Но если осеменение проведено в оптимальное время, то яйцепроводы и оболочки яйцеклетки, действуя совместно, исключают до минимума возможность одновременного И0стижения поверхности яйцеклетки двумя жизнеспособными спермиями. При старении яйцеклетки создаются условия для полиспермии.
Б. П. Хватов пересаживал крольчихам яйцеклетки с тремя пронуклеусами и получил нормальное потомство. По его мнению, в оплодотворении обычно участвует женский и один мужской пронуклеус, остальные же редуцируются. Но могут быть и такие случаи, когда из трех пронуклеусов сливаются два мужских, а женский редуцируется; или же участвуют все три пронуклеуса, но один из них – целиком, а два других – по половине. При этом диплоидный набор хромосом не нарушается. Б.П.Хватов считает, что наличием таких вариантов можно объяснить появление различных наследственных признаков при осеменении самок смешанной спермой.
Суперфекундация (superfecundatio) – оплодотворение нескольких яйцеклеток в период одной половой охоты спермой различных самцов. Суперфекундация характерна, главным образом, для многоплодных животных (собак, кошек, реже свиней, овец). Если самцы принадлежат к одной и той же породе, то суперфекундацию выявить трудно. Легче это сделать, если они относятся к различным породам или видам. В литературе описан случай, когда кобыла родила близнецов: жеребенка и мула. Несомненно, что в этом случае она покрывалась и жеребцом и ослом.
ТРАНСПЛАНТАЦИЯ ЗАРОДЫШЕЙ
Оплодотворенная яйцеклетка попадает в матку и развивается там до состояния зрелого плода. Развитие яйцеклетки при удачной трансплантации (пересадке) может происходить и в матке другого животного (реципиента), если в момент пересадки стадии полового Цикла коровы-донора эмбриона и реципиента совпадали.
Впервые пересадка зародышей была осуществлена Хиппом в 1890 г. у крольчих. С тех пор работы в этом направлении проводились и на других видах животных и это сделало трансплантацию зародышей одним из фундаментальных методов исследования воспроизводительной функции, в частности, – процессов оплодотворения и беременности. В настоящее время трансплантация зародышей имеет огромное практическое значение. Во многих странах функционируют коммерческие центры по трансплантации зародышей крупного рогатого скота и других животных.
Начало практического внедрения способа связано с успехами центра при Макдональдском колледже (Канада). Сотрудники колледжа в период с 1965 по 1969 г. успешно осуществляли трансплантацию яйцеклеток у крольчих, овец, свиней. Затем работы были перенесены на крупный рогатый скот и в 1973 г. получен живой теленок. С тех пор масштабы применения способа трансплантации зародышей резко возросли и в 1985 г. в США было получено несколько десятков тысяч телят-трансплантантов. В СССР за 1984-1987 гг. было проведено около 17 тыс. пересадок зародышей и получено более 3 тыс. телят. В эти годы началось внедрение метода трансплантации зародышей крупного рогатого скота и в Беларуси. В настоящее время в республике функционируют областные центры трансплантации эмбрионов, а также несколько пунктов в хозяйствах. Координируют их работу племживобъединения и лаборатория БелНИИЖ,
Значение метода в скотоводстве. В молочном скотоводстве пересадка зародышей позволяет ускорить и более точно провести оценку коров по качеству их дочерей, путем раннего получения зародышей от телок сократить интервалы между поколениями и ускорить селекцию, значительно увеличить распространение ценных генов путем отбора матерей для получения быков-производителей. В мясном скотоводстве можно добиться повышения продуктивности стад путем увеличения рождения двоен. Естественная двойневость У коров мясных пород составляет 2,8%, но вследствие размещения обоих эмбрионов в одном роге матки примерно у половины животных развитие их нарушается. Размещая эмбрионы в обоих рогах, можно увеличить число двоен до 20% и более. А при ранней идентификации пола станет возможным пересаживать только однополые зародыши и это предотвратит рождение телочек-фримартинов.
Пересадка зародышей может производиться коровам и с целью устранения бесплодия, если оно вызвано непроходимостью яйцепроводов, спайками яичников с яичниковым карманом, нарушением условий в матке вследствие старения и в других случаях. Применение этого метода позволяет многим странам вести обмен генофондом, который из-за ограничений правилами ветеринарного контроля в виде покупок животных осуществляется в небольших масштабах.
Пересадка зародышей дает возможность получать идентичных близнецов путем деления морулы или ранней бластоцисты на две или более частей, переносить ядра соматических клеток в зиготы и воспроизводить затем подобных особей.
Для получения оплодотворенных яйцеклеток in vitro и последующей пересадки их животным-реципиентам разработаны способы культивирования ооцитов из яичников убитых животных (фолликулярных ооцитов). Достигнут успех оплодотворения in vitro тубальных (извлеченных из яйцепроводов) зрелых ооцитов и последующей пересадки их реципиентам. В 1978 г. была извлечена посредством кесарева сечения девочка у 30-летней многорожавшей женщины, которой была пересажена оплодотворенная вне организма яйцеклетка. В 1982 г. впервые после оплодотворения тубальных ооцитов in vitro был получен теленок.
Большие возможности открывает трансплантация зародышей для развития генной инженерии (получение трансгенных животных, химерных животных и т.д.).
Метод трансплантации зародышей включает ряд приемов: вызывание суперовуляции у доноров и их осеменение; извлечение оплодотворенных яйцеклеток; синхронизацию половой охоты у реципиентов или же сохранение извлеченных зародышей до момента пересадки реципиентам с естественной половой охотой; пересадку зародышей реципиентам.
Вызывание суперовуляции у коров-доноров. Трансплантация зародышей может быть эффективной в том случае, когда за один прием от донора будет получено несколько половых клеток. А это возможно только в результате искусственного вызывания суперовуляции.
Вызвать суперовуляцию можно путем применения гонадотропина СЖК или очищенного ФСГ. Эти гормоны применяют в отдельности или в комплексе с гонадотропин-рилизинг-гормоном, простагландином, эстрогенами, хорионическим гонадотропином.
Существуют два основных способа вызова суперовуляции с помощью ГСЖК (фоллигон, интергонан, маротропин и др. препараты).
Дни цикла:
8-12-й
10-14-й
Через 48 часов 0
1-й
2-й
6-8-й
1500-3000 ЕД ГСЖК внутримышечно
25 мг ПГ-Ф2а или 500 мкг клопростенола внутримышечно
Половая охота
Первое осеменение
Второе осеменение
Нехирургическое извлечение зародышей
Гонадотропин СЖК оказывает более продолжительное действие, чем эндогенный или экзогенный ФСГ, что может быть причиной высокой изменчивости в числе и качестве овуляций, низкого выхода качественных эмбрионов. Для предотвращения нежелательных явлений вводят дополнительно антисыворотку на СЖК. Эстрогены вводятся для повышения оплодотворяемости, а хорионический гонадотропин и простагландины -для ускорения овуляции.
Главное достоинство применения ГСЖК для вызова суперовуляции – однократное введение животному всей дозы препарата.
Однако более стабильные результаты получают при использовании очищенного ФСГ, получаемого из гипофиза свиней. Общая доза на обработку препарата ФСГ-П (США) – от 32 до 50 мг. Инъецируют гормон два раза в день с промежутком 12 ч.
Дни цикла:
8-12-й
9-13-й
10-14-й
11-15-й
(0) 12-16-й
(1)
(8)
Простагландин целесообразнее вводить на 4-й день после начала обработки. Используют и другие схемы гормональной обработки коров-доноров эмбриона (в течение 5 дней по 10 мг ФСГ-П, простагландин на 3-4-й • день, начало обработки на 8-й день полового цикла; в течение 4 дней: 15 мг, 12 мг, 10 мг и 5 мг ФСГ-П, простагландин на 3-й день и т. д.). Дозу гормона регулируют в зависимости от серии препарата, срока его хранения, индивидуальной чувствительности животных и других факторов. Препараты фоллитропин (Литва), ФСГ (Беларусь), "ФСГ-СУПЕР" (Россия), фолликотропин (Чехия) и др. применяют в дозах, рекомендуемых наставлениями по применению этих препаратов.
У обработанных животных число овуляций варьирует от 0 да 50, чаще – 10-15. Это зависит от дозы и качества гормональных препаратов, индивидуальных особенностей животных и ряда других факторов. Увеличение дозы ГСЖК увеличивает число зрелых фолликулов, но процент овуляций может снизиться. Большие дозы препарата нередко вызывают образование кист яичников, геморрагических фолликулов или их лютеинизацию. Некоторые животные совершенно не реагируют на введение гонадотропных гормонов; таких коров может быть до 40%. Существует и ряд других проблем. Так, большое число созревающих фолликулов приводит к продлению процесса овуляции на несколько суток, а это отрицательно отражается на оплодотворяемости яйцеклеток и приживляемости их после пересадки реципиентам. При множественных овуляциях оплодотворяемость снижается, часть оплодотворенных яйцеклеток теряется вследствие увеличения яичника и неспособности воронки яйцепровода воспринять их. На результаты обработки оказывают влияние порода возраст животных, сезон года (в зимние месяцы число овуляций снижается), лактация и др.
В качестве коров - доноров эмбрионов отбирают ценных в генетическом отношении животных, хорошо реагирующих на введение гонадотропинов и дающих биологически полноценные зародыши. Молочная продуктивность донора за ряд лет должна быть выше стандарта данной породы в среднем на 50-60%, содержание жира в молоке – на 20% и более. Используют обычно коров, имеющих не менее двух-трех лактации, в возрасте не более 8-9 лет. После родов гормональную обработку не следует начинать ранее чем через 2-3 мес. До обработки животное должно проявить минимум два нормальных естественных половых цикла, иметь положительный баланс энергии, хорошее состояние половых органов и нормальный воспроизводительный статус в предыдущий репродуктивный период. Учитывают при отборе доноров эмбрионов и такие селекционные признаки, как породность, экстерьер, конституция, морфофункциональные свойства вымени, жизнеспособность приплода.
После отбора доноров и при необходимости вызвать суперовуляцию у нескольких животных одновременно им вводят дважды с промежутком 11 дней простагландин (клопростенол). Можно подкожно имплантировать синхромат (норгестамет). Через 9 дней имплантант удаляют и внутримышечно инъецируют простагландин (прозолвин, 3 мл). Через двое суток наблюдается половая охота.
Извлечение зародышей. Для извлечения зародышей предложено два метода: хирургический и нехирургический. Хирургический метод пригоден для всех видов животных и человека, но для крупного рогатого скота и лошадей в настоящее время он применяется главным образом в научных целях.
При хирургическом извлечении зародышей животное не кормят 1-1,5 сут и не поят 5-6 ч. Под общим трахеальным наркозом "веселящим" газом галотам в сочетании с тио- или оксибарбитуратом делают разрез по белой линии живота. Осматривают половые органы, подсчитывают количество желтых тел в яичниках, подтягивают один из рогов матки к просвету паны, делают небольшой разрез вблизи тела матки и катетер Фоллея вставляют в полость маточного рога до изгиба в сторону верхушки.
Катетер из латексной резины имеет основной канал диаметром 2-3 мм для оттока промывной жидкости и второй канал диаметром 0,5 мм для нагнетания воздуха в резиновый баллончик, находящийся на расстоянии 4-5 см от верхушки катетера. На передней части катетера имеются два отверстия на различных уровнях для оттока жидкости. Резиновый баллончик необходим для предотвращения вытекания жидкости в тело или другой рог матки. После введения катетера и нагнетания воздуха в баллончик прокалывают тупой иглой верхушку маточного рога и вводят через прокол канюлю, через которую подают промывную жидкость (60-120 мл); жидкость вытекает через катетер Фоллея. Для улучшения результатов нередко делают тубо-маточное промывание. В этом случае канюлю вставляют в яйцепровод в ампулярную часть и подают промывную жидкость (10-20 мл). Промывание яйцепроводов и матки позволяет получать более 70% яйцеклеток и зародышей от числа желтых тел, а при промывании только матки – около 45%. Однако промывание матки более безопасно.
У коров оперативный доступ может быть сделан под паралюмбальной и дополнительно местной инфильтрационной анестезией в области левой или правой голодной ямки.
У овец извлечение зародышей хирургическим способом проводят через 40-80 ч после проявления первых признаков половой охоты; возможно проведение операции и через 5-12 сут. В зависимости от сроков выполнения процедуры промывают различные участки половых путей. В первые трое суток промыванию подвергают яйцепроводы, а иногда и конечную часть рогов матки; на 5-7-й день делают смывы из большей части рогов матки, а после 9-го дня – промывают всю матку. При извлечении зародышей из яйцепровода в воронку его вводят стеклянную трубочку (внутренний диаметр 2 мм) на глубину до 3 см и свободный конец ее помещают в маркированную чашку Петри. Овечью сыворотку (2 мл и больше) вводят при помощи шприца в яйцепровод в месте соединения с маточным рогом, и всю жидкость собирают. Если собирают зародышей из рога матки, тогда шприц или канюлю вводят в рог матки на расстоянии 4 см от тубо-маточного соединения и свободный конец помещают в емкость (чашку Петри) для зародышей. В просвет матки в месте соединения с яйцепроводом шприцем вводят 2 мл сыворотки. Чтобы сыворотка не терялась, рог матки за канюлей или шприцем пережимают рукой. Если зародыши извлекают на 5-7-й день от начала охоты, то используют 5-10 мл сыворотки и делают смывы из большей части рогов матки. А для получения бластоцист через 9 дней после начала охоты промывают всю матку. При этом шейку матки пережимают зажимом, сыворотку вводят в верхушку одного рога матки, а собирают ее через канюлю, вставленную в противоположный яйцепровод.
У овец к числу овуляций получают до 80% зародышей.
При нехирургическом извлечении зародышей у коров катетер вводят поочередно в рога матки через шейку матки под ректальным контролем (рис. 59). Оптимальное время для извлечения - 7-8-й день от начала половой
охоты. К этому времени большинство зародышей будет находиться в верхушках рогов матки на протяжении около 10 см от соединения матки с яйцепроводом.
Для извлечения зародышей используются двух- или трехходовые катетеры (типа Фоллея) из латексной резины или телескопический комбинированный трехходовой инструмент с внутренним выдвигающимся эластическим катетером фирмы IMV (Франция).
Перед извлечением зародышей путем ректального исследования подсчитывают число желтых тел в яичниках коровы-донора. Наличие 10 или более желтых тел указывает на хорошую ответную реакцию животного на гормональную обработку, менее 5 желтых тел – на слабую реакцию. Для проведения операции корову фиксируют в станке в положении стоя. Желательно, чтобы тазовые конечности животного находились несколько выше передних. Для предупреждения частой дефекации и снятия мышечного напряжения делают сакральную анестезию. В эпидуральное пространство между крестцовой костью и первым хвостовым позвонком вводят 5 мл 2%-ного раствора новокаина. Строптивым коровам вводят внутримышечно миорелаксант (рампунь) 0,5-0,7 мл, или маточный релаксант комбилен 0,7-1,0 мл, илиханегиф 10 мл, или аминазин 5 мл. Теплой водой с мылом моют наружные половые органы и корень хвоста, вытирают бумажными салфетками или ватными тампонами.
Металлический катетер и переходники стерилизуют кипячением, латексный катетер обеззараживают в 96%-ном спирту, после чего промывают фосфатно-буферной средой для трансплантации зародышей. В латексный катетер затем вставляют стерильный металлический стилет и фиксируют в замковом узле кате-бра. Предварительно стилет смазывают стерильным вазелином или вазелиновым маслом. Снаружи кате-Р также можно слегка смазать вазелиновым маслом, чего его помещают в защитный полиэтиленовый чехол.
Катетер в чехле вводят во влагалище и направляют в канал шейки матки. Придерживая инструмент в та ком положении, сдвигают полиэтиленовый чехол назад, а катетер проводят через шейку матки и направляют в один из рогов матки. При этом металлический стилет вынимают из катетера постепенно, по мере введения катетера в рог матки. Важно правильно разместить катетер в роге матки. В большинстве моделей инструментов из латексной резины баллончик для воздуха находится на расстоянии около 5 см от его кончика. На этом участке в катетере проделано с различных сторон на различных уровнях четыре отверстия для выхода и входа промывной жидкости. Желательно, чтобы кончик катетера находился на расстоянии около 6 см от соединения яйцепровода с маткой. В этом случае расстояние от тубо-маточного соединения до баллончика составит приблизительно 10 см. Это как раз тот участок, за пределы которого зародыши еще не вышли. Нагнетают воздух в баллончик в количестве 15-30 см3. При этом обязательно контролируют рукой степень его наполнения и расширение рога матки, чтобы не допустить разрыва маточной стенки.
Как только баллончик будет наполнен воздухом, приступают к промыванию перекрытого (баллончиком) участка рога матки. Для этого полиэтиленовый шприц емкостью 50 мл (фирмы IMV) с фосфатно-буферной средой присоединяют к катетеру и вводят осторожно 30-35 мл среды. Затем шприц отсоединяют и жидкость из матки собирают в подготовленный сосуд; при этом открытый конец катетера оберегают от загрязнения. Опять присоединяют шприц и вводят столько же среды. В момент присоединения и отсоединения шприца катетер впереди металлического замкового устройства сдавливают пальцами. С каждым последующим введением количество среды увеличивают, доводя до 50-60 мл. А всего для промывания рога матки используют 500 мл среды. Обычно пользуются несколькими шприцами, что при быстром заполнении и смене их значительно уменьшает затраты времени на промывание.
Для предотвращения контакта жидкости для промывания матки с внешней средой и обеспечения асептики при извлечении зародышей при помощи двухходовых катетеров нами в систему катетер-шприц ключен трехходовый переходник фирмы IMV, из которого предварительно извлекаются клапаны и отсоединяется передняя канюля. К боковому патрубку переходника присоединяют эластичную трубку с зажимом. Свободный конец трубки помещают в сосуд для сбора поступающей из матки среды. После введения катетера в матку подготовленный переходник передним 'концом соединяют с замковым устройством катетера. Задний конец переходника служит для присоединения шприца со средой. В процессе промывания при открытом зажиме на эластичной трубке среда из рога матки поступает в сосуд, не контактируя с воздухом; шприц в таком положении должен оставаться соединенным с переходником. После закрытия зажима и передавливания пальцами катетера производят замену освобожденного шприца другим, заполненным средой. Такое усовершенствование процедуры обеспечивает стерильность среды для промывания и предупреждает потери зародышей, что возможно при.собирании поступающей из матки жидкости через открытый задний конец катетера. Можно сделать и совершенно замкнутую систему, если емкость со средой соединить трубкой (с зажимом) с задним концом переходника. В этом случае емкость должна быть расположена выше таза животного, чтобы среда поступала в матку самотеком.
После использования всего объема среды (500 мл) в рог матки можно ввести шприцем 30-40 мл воздуха. Иногда вводят воздух два-три раза. Это необходимо для более полного возвращения вводимой в матку среды. При этом имеет значение и умение специалиста контролировать состояние матки рукой через прямую кишку. Отсутствие больших потерь среды (из 500 мл не более 10-15 мл) и примесей в ней крови указывает на Достаточно техничное проведение операции.
Когда будет завершено промывание одного рога матки, воздух из баллончика выпускают и катетер извлекают из половых путей. Другой подготовленный катетер вводят в другой рог матки, и всю процедуру повторяют заново. Можно пользоваться и одним катетером, переводя его из одного рога в другой.
В качестве среды для промывания матки используют фосфатно-солевой буфер (ФСБ) Дюльбекко. ФСБ готовят путем смешивания трех различных стерильных растворов.
Раствор ФСБ-А
В 10 л деионизированной бидистиллированной воды растворяют:
натрий хлористый 100 г
калий хлористый 2,5 г
натрий фосфорнокислый двузамещенный
12-водный, высушенный до постоянной массы 14,4 г
калий фосфорнокислый однозамещенный 2,5 г
Буфер разливают по 400 мл в бутыли объемом 500 мл. Для стерилизации автоклавируют при давлении 1 атм в течение 20 мин. Хранят при комнатной температуре.
Раствор ФСБ-Б
В 1 л деионизированной бидистиллированной воды растворяют кальций хлористый безводный, высушенный до постоянной массы, 1 г. Для стерилизации автоклавируют при давлении 1 атм в течение 15 мин. Хранят при комнатной температуре.
Раствор ФСБ-В
В 1 л деионизированной бидистиллированной воды растворяют магний хлористый 6-водный, высушенный до постоянной массы 1 г. Для стерилизации автоклавируют при давлении 1 атм в течение 15 мин. Хранят при комнатной температуре.
Для получения полного ФСБ смешивают 400 мл ФСБ-А с 50 мл ФСБ-Б и 50 мл ФСБ-В. В таком виде буфер можно хранить при 4°С до 2 недель. Непосредственно перед употреблением в ФСБ вводят следующие компоненты (в расчете на 1 л): фетальную сыворотку теленка – 10-20 мл, пенициллин (калиевая соль) – 100 тыс. ЕД, глюкозу – 1 г, натрий пировино-граднокислый – 0,036 г. После этого выдерживают Б термостате при температуре 37,5°С в течение 1 ч и используют для промывания матки.
Завершив промывание, катетер полностью не извлекают и через него вводят в тело матки в 50-75 мл ФСБ, пенициллии и стрептомицин по 500 тыс. ЕД и тетрациклин 200 тыс. ЕД. Это необходимо для предупреж-цикия эндометрита. Если корову-донора используют неоднократно, то целесообразно в день половой охоты перед началом гормональной обработки (т. е. за 8-12 дней до нее) также ввести в матку антимикробные средства. Это будет хорошей гарантией нормального состояния эндометрия в день извлечения зародышей.
Отбор и подготовка животных-реципиентов. В момент трансплантации зародыша материнский организм находится в динамичном состоянии и изменяется в основном под действием стероидных гормонов (эстрогенов, прогестерона), которые доминируют в это время. Для успешного поддержания беременности необходимо четкое взаимодействие между зародышем и материнским организмом. Такое взаимодействие может быть достигнуто только при точной синхронизации половых циклов донора и реципиента. Необходимость синхронизации становится очевидной из того факта, что после овуляции и наступления естественной беременности развитие эмбриона и желтого тела идет одновременно. А потребность в питательных веществах у зародыша генетически скоординирована на основе избирательного поступления молекул субстрата через клеточные мембраны и паратипически контролируется химическим составом секрета яйцепроводов и матки. Любое нарушение синхронности изменений состояния зародыша и полового тракта может отрицательно сказаться на последующем протекании процессов формирования и развития зародыша.
У крупного рогатого скота разница во времени проявления половой охоты у донора и реципиента не должна быть более 12 ч. Если разница превышает 24 ч, то приживляемость пересаженных эмбрионов резко снижается.
При наличии многочисленной группы реципиентов (телок) в день трансплантации можно подобрать ивотных с желательной стадией полового цикла. Однако чаше половые циклы донора и реципиента синхронизируют искусственно. Достигается это инъекцией простагландина Ф2α животным-реципиентам в тот же срок, что и коровам-донорам. В качестве реципиентов могут быть как телки, так и коровы. Телок используют чаще, так как приживаемость зародышей у них выше, чем у коров. Телки-реципиенты должны иметь хорошее развитие, живую массу 350 кг и более и проявлять естественную половую цикличность. Отбирают их на второй день после начала гормональной обработки коров-доноров эмбрионов. Выбирают только животных с хорошо пальпируемыми желтыми телами в яичниках. Это гарантирует полный успех вызывания половой охоты после инъекции простагландина. Если синхронизация охоты проводится препаратами гестагенов в комбинации с простагландином, то обязательным условием для включения телок в группу реципиентов является наличие у них естественной половой цикличности, независимо от стадии цикла и развития желтого тела. Сроки начала применения гестагенов вычисляют исходя из продолжительности их воздействия. После введения простагландина за животными наблюдают и регистрируют время начала половой охоты. В день трансплантации зародышей их исследуют ректально и используют только тех телок, у которых в яичниках будут обнаружены хорошо сформированные, нормальные по величине желтые тела.
Оценка качества зародышей. После завершения промывания матки емкости со средой (нередко промывную жидкость собирают по отдельности из каждого рога матки) помещают на 20 мин в термостат при 37°С. Можно оставить их и при комнатной температуре (20-25°С). За это время зародыши, находящиеся в верхних слоях жидкости, осядут на дно. С помощью шприца и короткой эластичной трубки осторожно собирают пузырьки воздуха, которые располагаются по бокам сосуда на поверхности жидкости. Затем верхний слой жидкости удаляют с помощью сифона. Удоб^-но при этом пользоваться специальной металлической трубкой (фирмы IMV). Нижняя часть такой трубки на протяжении 1-1,5 см не имеет просвета, а сбоку на этом уровне в ней проделано отверстие. Вверху трубка изогнута под прямым углом. К этой части ее присоединяется эластичная трубка со шприцем. Длина эластичной трубки должна превышать высоту емкости, чтобы свободный конец ее опускался ниже дна емкости. Металлическая трубка помещается через отверстие в ообке (для устойчивости) до самого дна емкости. Шприцем слегка засасывают жидкость, чтобы она заполнила эластичную трубку, и затем отсоединяют шприц. Жидкость самостоятельно вытекает из емкости. Остается только нижняя часть ее высотой 1-1,5 см. Эту жидкость разливают в маркированные чашки Петри (2-4), дно которых расчерчено на полосы шириной около 1 см для удобства поиска зародышей. Содержимое чашек просматривают под бинокулярным микроскопом МБС 9 или МБС-10 при 15-25-кратном увеличении. В качестве источника освещения лучше использовать лампу дневного света.
В последнее время для облегчения процедуры отыскания зародышей все чаще используют специальные устройства с фильтром, через который пропускают промывную жидкость. Зародыши остаются в небольшом количестве среды на поверхности фильтра.
Обнаруженные зародыши специальным микроманипулятором или короткой пипеткой Пастера с резиновой трубочкой переносят в среду для кратковременного хранения (ФСБ + 20% фетальной сыворотки теленка). Такую среду готовят заранее и разливают по 2,5 мл в небольшие полиэтиленовые чашки Петри (2 мл ФСБ со всеми компонентами +0,5 мл фетальной сыворотки) или наносят на часовые стекла по 0,5-1 мл. С часовыми стеклами работать удобнее (зародыши собираются в центре), однако для соблюдения асептики при хранении зародышей при комнатных температурах более подходящими являются закрытые чашки Петри.
Оценивают качество зародышей в проходящем свете под микроскопом МБС-9 (или специальным инвертированным микроскопом) при 100-160-кратном увеличении. Часовые стекла или чашки Петри осторожно покачивают с целью осмотра зародышей со всех сторон. К моменту извлечения (7-8-й день) нормально Развитые зародыши находятся на стадии развития поздней морулы, ранней или поздней бластоцисты. Они имеют неповрежденную равномерную по ширине опа-лесцирующую прозрачную оболочку и цитоплазму в
тоянии дробления, соответствующем возрасту эмбриона. Морула ранняя (Мо I) или поздняя (Мо II) представляет собой скопление бластомеров, не всегда одинаковых по размеру из-за асинхронности дробления. Цитоплазма бластомеров гомогенная, а связь их полигональная. В пространстве прозрачной оболочки не имеется гранул или других включений. У бластоцисты ранней (Бл I) хорошо различимы группы зародышевых клеток (эмбриобласт) и поверхностных клеток (трофобласт), которые разделены бластополостью. Пространство прозрачной оболочки почти целиком заполнено. Поздняя (8-суточная) бластоциста (Бл II) имеет большую полость; клетки трофобласта уплощены, соприкасаются с прозрачной оболочкой, которая истончена и растянута; клетки эмбриобласта сгруппированы на одном полюсе. На 9-й день прозрачная оболочка разрывается и бластоциста полностью высвобождается из нее (рис. 60).
Морула ранняя с признаками дегенерации характеризуется несимметричным расположением бластомеров, различной их величиной, наличием в пространстве прозрачной оболочки гранул и других включений. Отдельные бластомеры могут быть разрушенными, но ДРУ; гие продолжают дробиться, достигая стадии поздней морулы или бластоцисты ("частичный" зародыш). ^ бластоцист также отмечается частичное разрушение клеток (в основном трофобласта), сжатие бластополос-ти и увеличение пространства прозрачной оболочки.
ма оболочка может иметь небольшие трещины. Де-рированные зарОдыШи отстают на 3-4 деления от Г<ормально развивающихся. Для них характерен рас-ал бластомеров, смещение или фрагментация цитоп-гзмы. Прозрачная оболочка имеет значительные де-(йекты, а в пространстве ее находятся фрагменты разрушенных бластомеров.
Морфологическую оценку морул и бластоцист проводят по 5-балльной шкале. Для трансплантации,отбирают морфологически нормальные зародыши с оценкой отлично и хорошо. При наличии свободных реципиентов допускается использование зародышей с оценкой удовлетворительно и условно годные. У морул и бластоцист хорошего качества в пространстве прозрачной оболочки имеются гранулы и включения, отмечаются некоторые различия в величине бластомеров, слабо выражена полость бластоцисты; с оценкой удовлетворительно - у морул разрушены единичные бластомеры, "частичный" зародыш, у бластоцист - бла-стополость не выражена, нет дифференциации между клетками трофобласта и эмбриобласта, у морул и бластоцист увеличено пространство прозрачной оболочки и в нем находятся гранулы и другие включения; с оценкой условно годные - сжатие и частичное разрушение бластомеров и нарушение связи между ними, фрагментация цитоплазмы, дефекты прозрачной оболочки и наличие в ее пространстве гранул, других включений. Выбраковывают неоплодотворенные яйцеклетки и зародыши с оценкой непригодные (несоответствие стадии развития возрасту зародыша, значительные дефекты прозрачной оболочки, распад бластомеров, сильное сжатие их).
Оттаянные после замораживания зародыши оценивают по тем же морфологическим признакам, что и свежеполученные.
Хранение и культивирование зародышей. В среде Для кратковременного хранения зародыши можно выдерживать при комнатной температуре не более 5-6 ч До момента использования. Однако наиболее высоких показателей приживаемости достигают при трансплантации их реципиентам в первые 1—1,5 ч с момента получения.
Глубокое замораживание и хранение зародышей в жидком азоте обеспечивает длительное поддержание их жизнеспособности, а также дает возможность проводить трансплантацию в любое удобное время. Для замораживания используют только свежеполученные зародыши отличного и хорошего качества. От момента получения до замораживания должно проходить как можно меньше времени. Замораживание проводят в растворах глицерина на ФСБ (или других криопротек-торов) с добавлением пенициллина и 20% фетальной сыворотки. Перед замораживанием зародыши последовательно помещают в заранее приготовленные растворы криопротектора с концентрацией 0,25 М (1 мл 0,5 М раствора + 1 мл ФБС), 0,50 М (2 мл 1,0 М раствора + 2 мл ФСБ), 0,75 М (1 мл 1 М раствора + 1 мл 0,5 М раствора глицерина) и 1,ОМ (0,74 мл глицерина + 9,26 мл ФСБ).
После выдержки в последнем растворе глицерина зародыши помещают в соломинки (по 1-2 зародыша в каждую). Заправку соломинок проводят в такой последовательности: раствор глицерина (2/5 объема соломинки), пузырек воздуха, зародыш в 1,4 М растворе глицерина (1 мл глицерина + 9 мл ФСБ, 1/5 объема соломинки), пузырек воздуха, 1,4 М раствор глицерина (2/5 объема). Столбик раствора криопротектора должен коснуться пробки. Свободный конец соломинки закрывают пластмассовой пробкой и подвергают замораживанию с помощью специальных устройств. Хранят в кассетах с держателем в сосудах Дьюара.
Оттаивают зародыши в водяной бане при 25°С или 37°С в течение 10-12 с. После оттаивания зародыши со средой переносят на часовые стекла и проводят предварительную морфологическую оценку. Затем помещают последовательно в растворы глицерина с концентрацией 1,0 М, 0,75 М, 0,50 М и 0,25 М, после чего трижды промывают в свежих порциях ФСБ и оставляют в такой же среде на 10-20 мин. По окончании промывки оценивают качество зародышей. Пригодные используют для трансплантации, зародыши сомнительного качества инкубируют в течение 2 ч и снова оценивают по морфологическим признакам.
Нередко морфологическая оценка свежеполученных зародышей также не дает полной уверенности в их жизнеспособности. Особенно это касается зародышей с значительными повреждениями прозрачной оболочки, разрушением бластомеров, сжатием бластополости (оцененных как условно годные). В таких случаях их культивируют в среде для кратковременного хранения в небольших чашках Петри, на часовых стеклах или в закрытых соломинках.
На часовые стекла наносят 0,2 мл среды, помещают в нее зародыши и среду покрывают слоем стерильного вазелинового масла (25 капель). Стекла помещают в чашки Петри и содержат их в эксикаторе при 37°С в термостате. До начала и в процессе культивирования через эксикатор пропускают увлажненную газовую смесь (90% азота, по 5% кислорода и углекислого газа).
В соломинку зародыши вводят с помощью специального микроманипулятора или шприца емкостью 1 мл с соединительным устройством. Сначала набирают в соломинку столбик среды (0,03 мл), затем столбик воздуха (0,01 мл), после этого среду с зародышем (0,03 мл), опять столбик воздуха и среду (0,02 мл). После соприкосновения среды с пробкой (поливиниловым спиртом) и затвердевания ее шприц отсоединяют, а соломинку с этой стороны закрывают пластмассовой пробкой и оставляют в термостате при 37°С на время инкубации.
Возможна инкубация зародышей и в малых чашках Петри в 2,5 мл среды без помещения их в эксикатор. Крышка чашки почти герметично закрывает среду, доступ воздуха ограничен, и это обеспечивает удовлетворительные условия для непродолжительной их выдержки (до 24 ч).
После культивирования проводят оценку жизнедеятельности зародышей. Продолжающие развитие зародыши с оценкой "отлично" и "хорошо" считают пригодными для пересадки.
Трансплантация зародышей реципиентам. При хирургическом методе трансплантации зародышей у телок-реципиентов под общим наркозом производят лапаротомию по белой линии живота или делают боковой разрез под местной анестезией. Через разрез извлекают рог матки, ипсилатеральныи (соответствующий) яичнику с желтым телом. Тупой иглой делают прокол в стенке матки ближе к месту отхождения широкой маточной связки. Непосредственно перед трансплантацией с помощью шприца или резиновой трубки с пробкой в пипетку Пастера, в кончике которой уже имеется небольшое количество среды для культивирования засасывается подходящий зародыш. Через проделанное отверстие кончик пипетки вводится в полость матки и нажатием на поршень шприца или сдавливанием резиновой трубки выталкивается зародыш со средой.
При совпадении половых циклов донора и реципиента приживаемость зародышей достигает 90%, при разнице в пределах 24 ч до 70%.
Овцам-реципиентам после лапаротомии по белой линии живота зародыши трансплантируют в яйцепро-вод или матку. Зиготы в состоянии от 2 до 6 бластоме-ров вводят пипеткой Пастера в минимальном количестве жидкости в яйцепровод на глубину 2-3 см. Зародыши с 8-16 бластомерами или бластоцисты вводят в матку пипеткой через отверстие, проделанное тупой иглой в стенке матки на границе с широкой маточной связкой. Используют зародыши в течение первых 3 ч с момента их получения. Хранят в стерильной сыворотке на часовых стеклах при 35°С. Перед трансплантацией зародыши отделяют от сыворотки. Успех трансплантации зависит от того, в какой рог помещена зигота: в ипсилатеральныи или контрлатеральный (противоположный) яичнику с желтым телом. В первом случае приживляемость зародышей значительно выше (85% против 35%). В момент трансплантации разница во времени наступления половой охоты у донора и реципиента не должна превышать 24 ч.
У крупного рогатого скота в практике используется нехирургический способ трансплантации зародышей реципиентам. Для пересадки применяют инструмент, предназначенный для ректо-цервикального способа осеменения (ШО-3 и ШО-4 или другой инструмент для введения спермы, расфасованной в соломинках). В соломинку зародыши помещают таким же способом, как и при их культивировании. Подготовленную соломинку стерильным пинцетом вставляют в инструмент и надевают на него полиэтиленовый одно-пазовый чехол.
Животное-реципиент фиксируют в станке в манеже центра (пункта) трансплантации зародышей или в помещении непосредственно в стойле. Наружные половые органы обмывают водой и дезинфицируют 0,2% -ным раствором фурацилина, 2%-ным раствором дио-цида или 70%-ным этиловым спиртом. Сильно возбудимым животным вводят комбилен внутримышечно в дозе 0,5 мл или ханегиф 10 мл. Затем делают сакральную анестезию 2%-ным раствором новокаина (5 мл). Подготовленный к пересадке инструмент вводят во влагалище до шейки матки, сдвигают назад защитный полиэтиленовый чехол и, пользуясь теми же приемами, что и при ректо-цервикальном способе осеменения, проводят его через канал шейки матки. После этого инструмент направляют в ипсилатеральныи яичнику с желтым телом рог матки. Осторожными движениями руки через прямую кишку расправляют рог матки и натягивают его на инструмент так, чтобы кончик инструмента был ближе к верхушке рога матки. Удерживая инструмент в таком положении, оператор дает команду помощнику, который медленным надавливанием на стилет вводит зародыш с содержимым в матку.
Все манипуляции с введением инструмента в половые пути должны быть выполнены в течение 1,5-2 мин. Если затрачивают больше времени, то это может стимулировать сокращение матки и привести к изгнанию зародыша. Следует отметить, что с 6-го дня матка проявляет слабую сократительную функцию, но длительные манипуляции с ней могут стимулировать эту функцию.
Приживаемость при нехирургической трансплантации свежеполученных зародышей составляет 50-60%, а при использовании сохраняемых в жидком азоте зародышей – 40-45%. С учетом этого и процента извлекаемых и бракуемых зародышей от коровы-донора можно получить в среднем два-три теленка. А поскольку Донора можно использовать для получения зародышей каждые 2,5-3 мес., то в течение года от такой коровы можно получить в среднем 10-12 телят. В Канаде от одной коровы за 9 мес. (9 процедур извлечения зароды шей) прижился после трансплантации реципиентам 51 зародыш.
БЕРЕМЕННОСТЬ
Беременность (graviditas) начинается с момента оплодотворения и в норме завершается рождением живого плода (плодов). Рост и развитие плода является результатом размножения клеток, их роста, изменения формы, структуры и функции. И хотя этот процесс является непрерывным, беременность принято делить на три периода: зиготы (сегментации), эмбриона (формирования тканей и органов и образования плаценты) и плода.
ПЕРИОД ЗИГОТЫ
Этот период начинается первым делением зиготы и заканчивается слабым прикреплением бластоцисты к стенке матки. Деление (дробление) зиготы происходит одновременно с продвижением ее по яйцепроводу. В результате первого деления образуется 2 бластомера, при следующем – 4 и т. д. (рис. 61). Дробление продолжается, но становится неравномерным и часто происходит неодновременно. Образуется все большее и большее количество клеток различного диаметра. Вся клеточная масса в полости прозрачной оболочки принимает форму плотного шара и похожа на небольшую ягоду шелковицы (морулу). У крупного рогатого скота морула образуется через 6 дней, у козы – через 5-6, у овцы – через 4 дня, у свиньи – через 110-114 ч. В это время она находится уже в матке. Но попадает зигота в матку еще раньше: у коров через 72-84 ч, у овец – через 66-72 ч после овуляции на стадии развития от 8 до 16 клеток; у свиней – через 46-48 ч на стадии 4 клеток, у кобыл – через 140-144 ч, когда они находятся уже на стадии бластоцист, у собак и кошек – на 5-8-й день.
У всех животных зиготы движутся по яйцепроводу неравномерно. Наиболее быстро они проходят расширенную часть и надолго задерживаются в истмической части, особенно в месте соединения яйцепровода с ро-ча матки. Продвижение их осуществляется за счет сокращений ресничек эпителия и движения жидкости которые направлены в сторону матки, а также за счет перистальтических сокращений яйцепровода, хорошо выраженных в первые дни после овуляции. Продвижение зигот регулируется гормонами: эстрогены замедляют и даже останавливают их продвижение, прогестерон, наоборот, ускоряет его.
В процессе дробления величина оплодотворенной яйцеклетки в первые дни почти не изменяется. И только с начала образования бластоцисты происходит ее увеличение. У коров в состоянии 80-120 бластомеров ранняя бластоциста имеет в диаметре 130-150 мкм, бла-стоциста (300-480 бластомеров) – 140-200 мкм и освободившаяся бластоциста (1500 бластомеров или более) – уже 200-1000 мкм. Образование бластоцисты происходит следующим образом. Клетки морулы разделяются (у коров на 7-й день, у свиней – на 4-й день) и начинает образовываться полый шар. Из скопившихся внутри этого шара клеток (эмбриобласта) развивается тело эмбриона. Тонкий наружный слой клеток (трофобласт) в основном обеспечивает питание зародыша и из него в дальнейшем образуются оболочки плода. Увеличивающаяся между ними полость приводит к разрушению прозрачной оболочки. У коров бластоциста освобождается от оболочки на 9-11-й (обычно на 10-й) день, у овец – между 7- и 8-м днями, у свиней – на 6-й день, у кобыл – через 8 дней. После этого бластоциста изменяет форму: из округлого мешка превращается в цилиндрический, быстро вытягивающийся по длине. Так, у овец длина зародыша составляет 10-22 мм к 11-му дню, около 10 см к 13-му дню и к 15-му дню его длина равна 15-19 см. В это время он может заходить уже в контрлатеральный рог матки. У свиней длина бластоцисты к 13-му дню достигает 33 см и более (иногда 55-191 см). У коров элонгация (удлинение) бластоцисты происходит позднее. Связано это с замедленной начальной фазой предимплантационного роста. У кобыл на 10-й день зародышевый пузырек имеет сферическую форму, которая в последующие 10 дней слабо изменяется.
Слабое прикрепление бластоцисты с помощью микроворсинок к эндометрию происходит у коров на 18-22-й день, у овец – на 16-й, у свиней – на 14-й, У собак – на 13-17-й, у кобыл – на 36-38-й день. У жвачных животных с двурогой маткой бластоциста прикрепляется обычно в том роге, на стороне которого в яичнике произошла овуляция. Иногда отмечается внутренняя (через тело матки) миграция зародыша в другой рог. У овец при одиночных овуляциях такое перемещение зародышей встречается в 8%, при множественных – в 85-90% случаев. У кобыл с 11-го по 15-й день отмечается частое перемещение зародыша из одного рога в другой. С 17-го дня внутриматочная миграция прекращается, начинается быстрый рост зародыша и он фиксируется в задней части тела вблизи бифуркации. У многоплодных животных бластоцисты располагаются по всей длине матки почти равномерно. Это связано с тем, что имплантированная бластоциста взаимодействует с эндометрием и создает вокруг себя зону невосприимчивости, в которой невозможно прикрепление другого зародыша. Обеспечивается это перистальтическими сокращениями миометрия. Каждая бластоциста служит местным раздражителем таких сокращений, в результате которых они раздвигаются на одинаковые расстояния. Мышечная реакция определяется уровнем секреции эстрогенов и (или) простагландинов самой бластоцистой. Наблюдения за свиньями с использованием меченых зародышей показали, что перераспределение их происходит внутри рога и между рогами матки на 9-11-й день и завершается к моменту имплантации (к 13-му дню). По мере имплантации чрезвычайно тонкий и длинный зародыш свиньи располагается в складках эндометрия в зигзагообразной форме.
В первый период питание зародыша осуществляется в основном за счет собственных энергетических запасов, а также за счет питательных веществ, всасываемых в яйцепроводе и матке. У свиней, например, до 8-го дня в клетках трофобласта сохраняется желток.
ПЕРИОД ЭМБРИОНА
В период эмбриона образуется большинство частей тела и органов зародыша, а также плодные оболочки. В конце периода наружная оболочка (хорион) прикрепляется к стенке матки и образующаяся плацента становится единственным источником питания плода.
У крупного рогатого скота период эмбриона продолжается с 13-го по 45-й день беременности. За это время из клеток эмбриобласта образуются ткани и органы эмбриона. Уже на 20-22-й день начинается сердцебиение (180-190 сокращений в минуту, на 26-й день -140-150); развиваются легкие, печень, поджелудочная железа. Начинается развитие первичной почки и половых органов. На 25-28-й день обнаруживаются зачатки конечностей, а через одну-две недели на них можно различить суставы. В 37 дней ясно выражены "черты лица". Плодные оболочки развиваются с третьей недели. В начале энтодерма (один из трех зародышевых листков) обрастает трофобласт изнутри (рис. 62). Трофобласт с боков приподнимается, смыкается над зародышевым узелком, срастается и перешнуровывается. Образуется две оболочки: амнион (водная), которая окружает плод, и хорион (сосудистая). Позднее между ними из мезодермы через пупочное кольцо выпячивается в виде слепого мешка третья оболочка – алланто-ис (мочевая). В 18 дней амнион окружает эмбрион; вскоре (около 23 дней) образуется и аллантоис. Весь зародышевый пузырек (трофобласт) быстро вытягивается в длину; в 16 дней он занимает две третьи длины рога матки (от 70 до 240 мм), на 17-й день – полностью беременный рог, на 19-20-й день достигает верхушки противоположного рога матки. В период удлинения толщина зародышевого пузырька на всем протяжении составляет около 2 мм. К концу первого месяца эмбрион имеет длину около 1 см и вместе с амнионом представляет собой маленький пузырек. К 35 дням хорион хорошо сформирован в обоих рогах матки, а аллантоис заполняет' полость хориона в беременном роге и подводит сосуды к ворсинкам.
Первые плацентомы обнаруживаются в 35 дней, когда начинается развитие и прикрепление котиледонов к карункулам. В наиболее широкой части диаметр плодного пузыря составляет 4-5 см. С 35-40-го дня можно определить наличие беременности ректальным методом.
У овец в момент слабого прикрепления к эндометрию (16-й день) зародышевый пузырек имеет длину 15-19 см (рис. 63). Образующиеся микроворсинки трофобласта взаимодействуют с поверхностью эндометрия. Растут оболочки и начинают заходить в противоположный рог матки. На 18-й день они полностью заполняют его полость. Процесс имплантации ускоряется и в основном завершается к 50-му дню. У эмбриона заложены все органы, оформлены видовые очертания.
У свиней период эмбриона длится с 14-го по 45-й день. Прикрепление зародышевого пузырька начинается с появлением на 13-14-й день коротких ворсинок на трофобласте и микроворсинок в соответствующих участках эндометрия. К 18-му дню прикрепление зародыша хорошо выражено, на 21-22-й день при "снятии" его с эндометрия необходимо проявить большую осторожность, чтобы не повредить целостность трофобласта.
У лошадей интенсивность роста бластоцисты после 17-го дня не увеличивается, а с 27-го дня становится даже меньше. В 36 дней она остается скорее цилиндрической (диаметром около 9 см) и более сильно растянута аллантоисной жидкостью. Благодаря этому происходит заметное увеличение матки, что способствует проведению ректальной диагностики беременности в ранние сроки. На 36-38-й день происходит тесное присоединение клеток хориона к лежащему под ним эпителию эндометрия. На 45-й день рудиментарные ворсинки появляются на всей поверхности аллантохорио-на и внедряются в материнские крипты.
С самого начала беременности зародыш проявляет трофическое воздействие на желтое тело, поэтому функция его продолжается и после истечения срока существования, характерного для желтого тела полового цикла. Конец этого срока у оплодотворенных самок является наиболее критическим моментом, после которого секреция прогестерона усиливается; у небеременных животных в матке в это время вырабатываются лютео-лизины, которые вызывают прекращение функционирования желтого тела. Однако установление беременности у животных является не простым включением или выключением функции желтого тела, а представляет комплекс взаимоотношений, зависящих от тонкого баланса между лютеотропными и лютеолитически-ми факторами. Причем имеют значение не только взаимоотношения между яичниками и маткой, но также взаимосвязь между этими органами и гипофизом и определенное посредничество гипоталамуса.
Особое значение для имплантации и развития беременности имеет активность самой бластоцисты. В основе имплантации, как и других процессов развития, лежит взаимодействие тканей. Конечным результатом ее является установление функциональной связи зародыша с сосудистой системой матери и образование плаценты. У крупного рогатого скота распознавание беременности матерью происходит на 16-17-й день, у кобыл – на 14-16-й, у свиней – на 12-й, у овец – на 12-13-й и у коз – на 17-й день.
Для распознавания беременности и сохранения функции желтого тела у крупного рогатого скота большое значение имеет удлинение бластоцисты. Удаление ее из матки на 17 или 19-й день увеличивает продолжительность полового цикла до 25-26 дней, а при удалении на 13-й день продолжительность цикла не изменяется (Northey and French, 1980). Антилютеолитиче-ский сигнал зародыш дает путем секреции трофобластного протеина (bovine TP-1) с молекулярной массой около 24000; начинается секреция с 16-17-го дня и продолжается до 38-го дня с максимумом между 16 и 19 днями. Это вещество аналогично овечьему ТР-1 и ос-интерферону и обладает антивирусной активностью. При внутриматочном введении ТР-1 нестельным коровам на 14-17-й день цикла срок функционирования желтого тела увеличивается. Это же наблюдается и при внутриматочном или внутримышечном введении интерферона (INF-oc). Антилютеолитическое действие протеина проявляется путем модификации эндометриальных рецепторов к окситоцину, угнетения выделения ПГ-Ф2а-
Желтое тело беременности функционирует в течение всего срока стельности. Удаление его вместе с яичником в первые 200 дней вызывает аборт. Удаление после 200-го дня только желтого тела, а после 220-го яня яичника с желтым телом не прерывает беременности, но укорачивает ее. При этом повышается возможность задержания последа. У этого вида желтое тело является основным источником прогестерона, а другие ткани яичника, надпочечники и плацента сек-ретируют гормон в небольших количествах. Однако после 220-го дня благодаря накоплению прогестерона в жировой ткани и постепенному его использованию оказывается достаточным и плацентарного прогестерона. Поэтому овариэктомия аборта не вызывает.
В первые 14 дней у коров стельности уровень прогестерона такой же, как и в период диэструса. К 18-му дню в отличие от нестельных животных, у которых наблюдается резкое снижение прогестерона, падение гормона незначительное. Наивысшая концентрация прогестерона наблюдается к концу формирования плаценты (на 60-70-й день – 6,2-6,4 нг/мл). Затем уровень его несколько снижается к 150-160-му дню (3,0-3,8 нг/мл) и значительно падает за 2-5 дней до родов (менее 1,5 нг/мл). При многоплодной беременности концентрация гормона значительно выше как в период формирования плаценты (14,2 нг/мл), так и перед отелом – 4,9 нг/мл (П.И.Гавриченко, 1996). За 8 ч до родов начинается быстрое снижение уровня гормона с достижением минимального уровня сразу же после родов.
Концентрация эстрогенов низкая в начале и середине беременности (менее 100 пг/мл), но в конце ее, особенно после 250-го дня, уровень эстрогенов повышается, достигая пика за 2-5 дней до отела – 7 нг/мл эстрона сульфата и 1,2 нг/мл эстрона (Thatcher et. al., 1980). За 8 ч до родов начинается быстрое снижение гормона с достижением минимального уровня сразу же после родов. Концентрация активной фракции эст-рогенов-эстрадиола 17р в период беременности колеблется незначительно (101-140 пг/мл), существенно возрастая только с приближением отела – до 1,8-3,0 нг/мл (Н.И. Гавриченко, 1996).
Содержание ФСГ и ЛГ в течение стельности низкое, а колебания незначительные. Низкое содержание и пролактина; однако количество его увеличивается непосредственно перед отелом с 50-60 нг/мл до 320 нг/адл за 20 ч до родов с последующим уменьшением до ба-зального уровня через 30 ч. Плацентарный (bovine) лак-тоген (по-видимому, проявляющий активность пролак-тина и гормона роста) появляется в периферической крови к 160-му дню стельности и достигает максимальной концентрации (1000 нг/мл) между 200 днями и концом стельности.
У небеременных овец содержание прогестерона быстро падает в конце полового цикла. После оплодотворения максимальная концентрация гормона, характерная для диэструса (2,5-4 нг/мл), поддерживается и постепенно увеличивается до 60 дней суягности. В это время желтое тело является основным источником прогестерона. Затем плацента начинает секретировать значительное количество гормона и его содержание достигает максимума на 130-140-й день. В последнюю неделю содержание прогестерона быстро снижается и достигает 1 нг/мл к моменту родов. У маток с одним плодом на 105-110-й день суягности содержание прогестерона максимальное – 3,78 нг/мл; с двумя и тремя плодами на 125-130-й день – 5,09 и 9,18 нг/мл (Emady et. al., 1974).
Бластоциста овец секретирует 3-4 изоформы протеина с молекулярной массой около 18000. Секреция трофэктодермой оТР-1, который также аналогичен интерферону, начинается с момента удлинения зародышевого пузырька (около 10 дней) и продолжается до 23-го дня. Периодическое (пульсирующее) выделение ПГ-Ф2а, которое имеет место на 15-17-и день у небеременных овец, снижается. Интерферон и протеин тормозят активность маточных рецепторов к окситоцину, концентрация которых снижается под влиянием прогестерона. Предполагается также, что протеин непосредственно или косвенно угнетает секрецию ПГ-Ф2о и увеличивает секрецию Пг-К2. Возможно и превращение





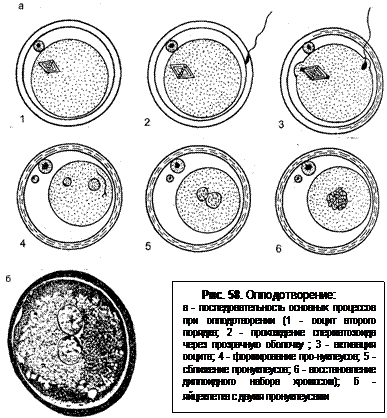 После соприкосновения спермия с желточной оболочкой происходит образование второго полярного тельца и ооцит второго порядка превращается в зрелую яйцеклетку. Затем спермий полностью погружается в цитоплазму клетки. Входит он не перпендикулярно, а по касательной по отношению к желточной оболочке (рис. 58). После вхождения в яйцеклетку головка спермия набухает, теряет форму и оболочку и становится едва заметной, ядерный материал выходит из нее и вскоре появляются в виде мелких точек ядрышки. Они постепенно соединяются и образуют большие, круглые частицы, которые включаются в мужской пронуклеус; пронуклеус окружен ядерной оболочкой.
После соприкосновения спермия с желточной оболочкой происходит образование второго полярного тельца и ооцит второго порядка превращается в зрелую яйцеклетку. Затем спермий полностью погружается в цитоплазму клетки. Входит он не перпендикулярно, а по касательной по отношению к желточной оболочке (рис. 58). После вхождения в яйцеклетку головка спермия набухает, теряет форму и оболочку и становится едва заметной, ядерный материал выходит из нее и вскоре появляются в виде мелких точек ядрышки. Они постепенно соединяются и образуют большие, круглые частицы, которые включаются в мужской пронуклеус; пронуклеус окружен ядерной оболочкой.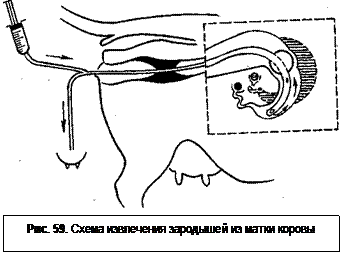 охоты. К этому времени большинство зародышей будет находиться в верхушках рогов матки на протяжении около 10 см от соединения матки с яйцепроводом.
охоты. К этому времени большинство зародышей будет находиться в верхушках рогов матки на протяжении около 10 см от соединения матки с яйцепроводом.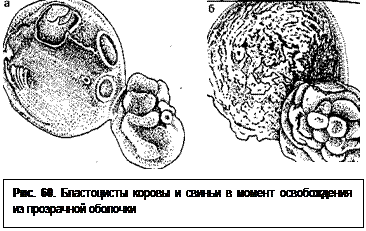 Глубокое замораживание и хранение зародышей в жидком азоте обеспечивает длительное поддержание их жизнеспособности, а также дает возможность проводить трансплантацию в любое удобное время. Для замораживания используют только свежеполученные зародыши отличного и хорошего качества. От момента получения до замораживания должно проходить как можно меньше времени. Замораживание проводят в растворах глицерина на ФСБ (или других криопротек-торов) с добавлением пенициллина и 20% фетальной сыворотки. Перед замораживанием зародыши последовательно помещают в заранее приготовленные растворы криопротектора с концентрацией 0,25 М (1 мл 0,5 М раствора + 1 мл ФБС), 0,50 М (2 мл 1,0 М раствора + 2 мл ФСБ), 0,75 М (1 мл 1 М раствора + 1 мл 0,5 М раствора глицерина) и 1,ОМ (0,74 мл глицерина + 9,26 мл ФСБ).
Глубокое замораживание и хранение зародышей в жидком азоте обеспечивает длительное поддержание их жизнеспособности, а также дает возможность проводить трансплантацию в любое удобное время. Для замораживания используют только свежеполученные зародыши отличного и хорошего качества. От момента получения до замораживания должно проходить как можно меньше времени. Замораживание проводят в растворах глицерина на ФСБ (или других криопротек-торов) с добавлением пенициллина и 20% фетальной сыворотки. Перед замораживанием зародыши последовательно помещают в заранее приготовленные растворы криопротектора с концентрацией 0,25 М (1 мл 0,5 М раствора + 1 мл ФБС), 0,50 М (2 мл 1,0 М раствора + 2 мл ФСБ), 0,75 М (1 мл 1 М раствора + 1 мл 0,5 М раствора глицерина) и 1,ОМ (0,74 мл глицерина + 9,26 мл ФСБ).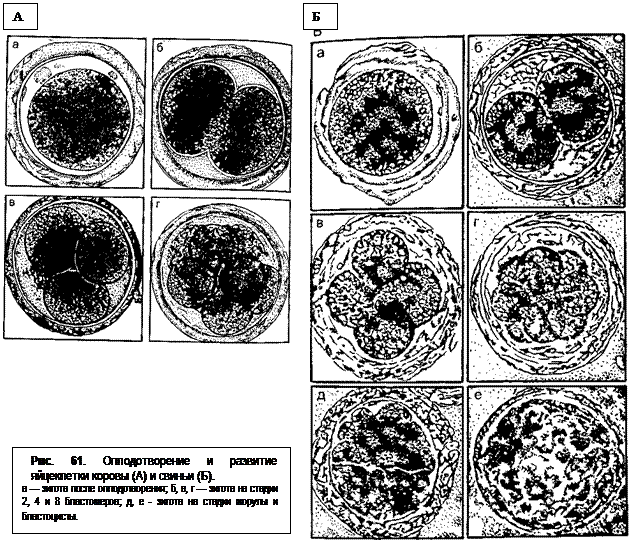 У всех животных зиготы движутся по яйцепроводу неравномерно. Наиболее быстро они проходят расширенную часть и надолго задерживаются в истмической части, особенно в месте соединения яйцепровода с ро-ча матки. Продвижение их осуществляется за счет сокращений ресничек эпителия и движения жидкости которые направлены в сторону матки, а также за счет перистальтических сокращений яйцепровода, хорошо выраженных в первые дни после овуляции. Продвижение зигот регулируется гормонами: эстрогены замедляют и даже останавливают их продвижение, прогестерон, наоборот, ускоряет его.
У всех животных зиготы движутся по яйцепроводу неравномерно. Наиболее быстро они проходят расширенную часть и надолго задерживаются в истмической части, особенно в месте соединения яйцепровода с ро-ча матки. Продвижение их осуществляется за счет сокращений ресничек эпителия и движения жидкости которые направлены в сторону матки, а также за счет перистальтических сокращений яйцепровода, хорошо выраженных в первые дни после овуляции. Продвижение зигот регулируется гормонами: эстрогены замедляют и даже останавливают их продвижение, прогестерон, наоборот, ускоряет его.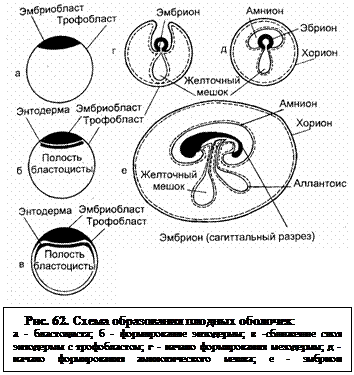 В период эмбриона образуется большинство частей тела и органов зародыша, а также плодные оболочки. В конце периода наружная оболочка (хорион) прикрепляется к стенке матки и образующаяся плацента становится единственным источником питания плода.
В период эмбриона образуется большинство частей тела и органов зародыша, а также плодные оболочки. В конце периода наружная оболочка (хорион) прикрепляется к стенке матки и образующаяся плацента становится единственным источником питания плода.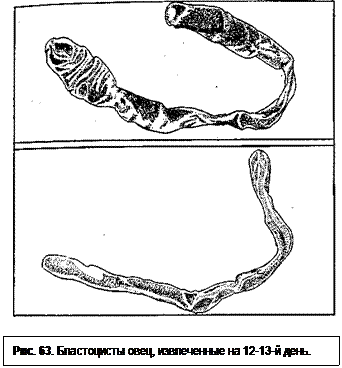 Первые плацентомы обнаруживаются в 35 дней, когда начинается развитие и прикрепление котиледонов к карункулам. В наиболее широкой части диаметр плодного пузыря составляет 4-5 см. С 35-40-го дня можно определить наличие беременности ректальным методом.
Первые плацентомы обнаруживаются в 35 дней, когда начинается развитие и прикрепление котиледонов к карункулам. В наиболее широкой части диаметр плодного пузыря составляет 4-5 см. С 35-40-го дня можно определить наличие беременности ректальным методом.
