




Стекловидное тело после его повреждения (проникающее ранение или хирургическое вмешательство с «выпадением» стекловидного тела) не восстанавливается. В области повреждения отсутствует волокнистый компонент, а дефект выполняется содержащими белок электролитами. При этом стекловидное тело мутнеет. К морфологическим признакам заместительной регенерации стекловидного тела можно отнести миграцию в область повреждения и последующую пролиферацию глиальных элементов сетчатой оболочки, а также расположенных преретинально микроглиальных клеток. Разрастание указанных клеток приводит к еще большему помутнению и развитию глиальных тяжей. Последнее обстоятельство является одной из основных причин развития отслойки сетчатки.
Необходимо отметить, что проводится большое количество исследований для выяснения возможности стимуляции репаративной регенерации стекловидного тела. Для этих целей пытаются использовать культуру ткани гиалоци-тов, синтезирующих волокнистый и основной компоненты стекловидного тела. К сожалению, до сих пор исследования носят экспериментальный характер.
3.6. СЕТЧАТКА
Сетчатая оболочка (retina) привлекала внимание исследователей на протяжении многих веков. Первым описал ее Chacedon в 330 г. до н. э. Название этой структуре дал Rufos Ephe-sus (приблизительно 110 г. н. э.), который предполагал, что сетчатка является сетью, поддерживающей стекловидное тело.
На протяжении многих веков ни у одного из исследователей не возникало мысли о связи сетчатки с мозгом. Лишь Кеплер в 1608 г. предположил о том, что сетчатка является «первичной тканью зрительного рецептора».
Первое детальное микроскопическое исследование сетчатки проведено Тревианусом (Тге-vianus) в 1835 г. Последующее совершенствование микроскопической техники, приготовления тонких срезов и методов окрашивания препаратов позволило выявить нейронную организацию сетчатки, а также особенности синаптических контактов между нейронами и роль нейронных связей в обработке зрительной информации.
Изучению сосудистой сети сетчатки способствовало развитие методов исследования плоскостных препаратов сетчатки после обработки ее трипсином, применения методов флюоресцентной ангиографии. Бурное развитие нейроанатомии сетчатой оболочки связывают с развитием методов иммуногистохимии, позволяющих с большой точностью выявить в определенной структуре сетчатки специфические вещества, в частности нейротрасмиттеры. Сочетание методов морфологии, иммуногистохимии и нейрофизиологии (регистрация мембранных потенциалов отдельной клетки) позволило к настоящему времени получить достаточно полную картину относительно механизмов восприятия и обработки световой энергии сетчатой оболочкой.
Общая анатомия. Сетчатка является частью внутренней оболочки глаза (tunica internet bulbi) и представляет собой прозрачную ткань, выстилающую внутреннюю поверхность глазного яблока, занимая при этом 3/4 ее площади. Распространяется она от диска зрительного нерва до зубчатой линии (ora serrata), переходя в этой области в пигментный эпителий ресничного тела. Сенсорная (световоспринимаю-щая) часть сетчатки прилежит к пигментному эпителию сетчатки, от которого она легко отделяется. Наиболее сильная связь с подлежащими тканями определяется в области зубчатой линии и у края диска зрительного нерва, вблизи желтого пятна (macula luted).
В области экватора сетчатка имеет вертикальный диаметр 24,08 ±0,94 мм и горизонтальный — 24,06 ±0,60 мм. Расстояние от края диска зрительного нерва до верхней части экватора равняется 14,71 ±1,08 мм, до нижней части— 14,51 ±1,01 мм, с носовой стороны— 13,27 ±1,11 мм, с височной стороны — 17,29 ±1,6 мм. В указанных границах площадь сетчатой оболочки равняется 1206 мм2. Переднюю часть сетчатки рассматривают от экватора до зубчатой линии. При этом расстояние от экватора до зубчатой линии с височной стороны равно 6,0 ±1,22 мм, с носовой стороны — 5,8 ±1,12 мм, сверху — 5,07 ±1,11 мм, снизу— 4,79 ±1,22 мм. Расстояние от переднего края сетчатки до линии Шальбе сверху равно 6,14 ±0,85 мм, снизу — 6,2 ±0,76 мм, с носовой стороны — 5,73 ±0,81 мм и с височной — 6,52 ±0,75 мм [1044].
Микроскопическая анатомия. Сетчатка является наиболее сложным в структурном и
Сетчатка
229

 функциональном отношениях образованием глаза и выполняет основную функцию — фоторецепцию. Столь сложное в структурном и функциональном отношениях образование можно рассматривать с разных позиций. По этой причине существует несколько классификаций ее строения — функциональная классификация, гистогенетическая и анатомическая. В соответствии с функциональной классификацией сетчатку подразделяют на нейроны, глию и сосудистую систему.
функциональном отношениях образованием глаза и выполняет основную функцию — фоторецепцию. Столь сложное в структурном и функциональном отношениях образование можно рассматривать с разных позиций. По этой причине существует несколько классификаций ее строения — функциональная классификация, гистогенетическая и анатомическая. В соответствии с функциональной классификацией сетчатку подразделяют на нейроны, глию и сосудистую систему.
Гистогенетическая классификация отличается тем, что отдельные структуры сетчатки подразделяют в соответствии с особенностями их происхождения. В этой связи выделяют производные нейроэпителия (нейроны, глия), мезенхимы (сосудистая система).
Анатомическая классификация описывает особенности микроскопического строения сетчатки. Именно на ней мы и остановимся в этом разделе. Морфо-функциональные особенности сетчатой оболочки будут приведены в главе 4.
При световой микроскопии в сетчатке выделяют 11 слоев (рис. 3.6.1, см. цв. вкл.):
1. Мембрана Бруха.
2. Пигментный эпителий сетчатки.
3. Слой фоторецепторов, палочек и кол
бочек.
4. Наружная пограничная мембрана.
5. Наружный ядерный слой.
6. Наружный плексиформный (сетчатый)
слой.
7. Внутренний ядерный слой.
8. Внутренний плексиформный (сетчатый)
слой.
9. Слой ганглиозных клеток.
10. Слой нервных волокон.
11. Внутренняя пограничная мембрана.
Ряд исследователей мембрану Бруха рассматривают одновременно с сосудистой оболочкой. Гистогенетически мембрана Бруха одновременно относится как к сосудистой оболочке, так и сетчатой оболочке.
Пигментный эпителий
При удалении внутренней сенсорной части сетчатки от внутренней поверхности глазного яблока открывается пигментный эпителий (пигментная часть сетчатки; pars pigmentosa). Выглядит он в виде коричневой непрерывной пластинки, простирающейся от зрительного нерва до зубчатой линии. Затем он переходит на ресничное тело в виде пигментного эпителия. Наиболее пигментирован эпителий в области желтого пятна. Пигментный эпителий сетчатой оболочки выполняет многообразные функции. Первоначально предполагали, что пигментный эпителий является просто черным фоном, снижающим рассеивание света в процессе фоторецепции. В конце XIX в. установили, что отделение сенсорной части сетчатки от
пигментного эпителия приводит к потере зрения [298]. Это исследование позволило предположить важную роль пигментного эпителия в фоторецепции. Многочисленные исследования последнего времени установили наличие взаимодействия клеток пигментного эпителия с фоторецепторами. Использование электоронной микроскопии выявило наличие фагоцитарной активности эпителиоцитов [1032, 1219, 1226].
Определенную роль в установлении функции пигментных клеток сыграло применение культуры тканей [278, 484, 821].
Мы перечислим лишь некоторые из функций пигментного эпителия сетчатки. Более подробные сведения приведены в табл. 3.6.1.
Та б л и ца '3.6.1. Функции пигментного эпителия сет чатой оболочки (по linn, Benjamin-Henkind, 1979)
 Физические
Физические
1.  Выполняет барьерные функции по отношению
Выполняет барьерные функции по отношению
сенсорной части сетчатки, не допуская крупные моле
кулы со стороны хориоидеи.
2. Обеспечивает адгезию сенсорной части сетчат
ки с пигментным эпителием посредством транспорта
специфических жидких компонентов и взаимодействия
микроворсинок клеток пигментного эпителия с наруж
ными члениками фоторецепторов и синтеза компонен
тов межфоторецепторного матрикса.
Оптические
1. Абсорбция световой энергии (гранулы мелани
на), «обрезая» рассеянный свет, повышает при этом
разрешающую способность зрительной системы.
2. Является барьером на пути проникновения све
товой энергии через склеру, повышая разрешающую
способность зрительной системы.
3. Максимально поглощает энергию лазерных из
лучателей (аргоновый, рубиновый, криптоновый ла
зеры) благодаря абсорбционной способности мелано-
сом, приводя к фототермическому эффекту. Последнее
свойство является основой фотокоагуляции.
Метаболические
1. Фагоцитирует наружные членики палочек и кол
бочек.
2. Переваривает структурные элементы фагоцити
рованных наружных члеников палочек и колбочек (ге-
терофагия) благодаря наличию хорошо развитой лизо-
сомной системы.
3. Эстерификация, изомеризация, хранение и транс
порт витамина А.
4. Синтез межклеточного матрикса; апикального
компонента межфоторецепторного матрикса: базально-
го компонента базальной мембраны.
5. Содержит ферменты для синтеза зрительного
хроматофора 11-цис-ретинала; гранул меланина (тиро-
зиназы); ферментов детоксикации (цитохром Р450);
и др.
6. Транспорт большого количества метаболитов к
зрительным клеткам и от них в направлении сосудис
той оболочки.
230
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 Окончание табл. 3.6.1
Окончание табл. 3.6.1
 Транспортные
Транспортные
1. Активный транспорт ионов НСО3, определяющих
выведение жидкости из субретинального пространства.
2. Na+/K+-Hacoc, обеспечивающий перенос солей
через клетки пигментного эпителия. Перенос воды
осуществляется пассивно.
3. Активный АТФ-зависимый перенос ионов Mg2+—
Са2+.
4. Насосная система, обеспечивающая отток боль
шого объема воды из стекловидного тела.
 Пигментный эпителий способствует формированию фоторецепторов в эмбриогенезе, индуцируя этот процесс, обеспечивает функционирование гемато-ретинального барьера, поддерживает постоянство среды между пигментным эпителием и фоторецепторами, поддерживает структуру контакта между наружными сегментами палочек и колбочек и клетками пигментного эпителия, обеспечивает активный избирательный транспорт метаболитов между сетчаткой и увеальным трактом, осуществляет транспорт, накопление и изомеризацию витамина А, осуществляет фагоцитоз наружных сегментов фоторецепторов, а также поглощение световой энергии гранулами меланина, осуществляет синтез гликозаминогликанов, окружающих наружные сегменты фоторецепторов.
Пигментный эпителий способствует формированию фоторецепторов в эмбриогенезе, индуцируя этот процесс, обеспечивает функционирование гемато-ретинального барьера, поддерживает постоянство среды между пигментным эпителием и фоторецепторами, поддерживает структуру контакта между наружными сегментами палочек и колбочек и клетками пигментного эпителия, обеспечивает активный избирательный транспорт метаболитов между сетчаткой и увеальным трактом, осуществляет транспорт, накопление и изомеризацию витамина А, осуществляет фагоцитоз наружных сегментов фоторецепторов, а также поглощение световой энергии гранулами меланина, осуществляет синтез гликозаминогликанов, окружающих наружные сегменты фоторецепторов.
Клетки пигментного эпителия фагоцитируют до 10% наружных члеников фоторецепторов ежедневно. Способность фагоцитировать наружные сегменты палочек и колбочек является прямым доказательством постоянной регенерации последних.
Поглощение световой энергии меланиновы-ми гранулами обеспечивает четкую топографическую регистрацию световой энергии наружными сегментами фоторецепторных клеток, окутанных отростками клеток пигментного эпителия, содержащими зерна меланина. Это обеспечивает световую изоляцию каждого фоторецептора. При усилении освещенности глазного яблока зерна меланина мигрируют в отростки клеток пигментного эпителия. При этом степень изоляции фоторецепторов усиливается.
Поглощение и транспортировка ретинола (витамин А) обеспечивается рецепторами, расположенными на базальной и латеральной поверхностях клеток пигментного эпителия. Клетки пигментного эпителия синтезируют особый гликопротеид, который переносит ретинол в интерфоторецепторный матрикс, откуда он и поступает в фоторецепторы [371, 483].
Необходимо отметить, что нарушение функции пигментного эпителия лежит в основе развития ряда заболеваний. Его структурные изменения выявлены при возрастной макулопатии, центральной серозной ретинопатии, дистрофии сетчатки. Эти изменения хорошо выявляются офтальмоскопически.
Клетки пигментного эпителия чувствительны к ряду токсинов [397].
Пигментный эпителий сетчатки расположен между хориокапиллярным слоем сосудистой оболочки и сенсорной частью сетчатки (рис. 3.6.1 (см. цв. вкл.) —3.6.4). Он представляет собой один слой уплощенных интенсивно пигментированных клеток, плотно прилежащих друг к другу и имеющих гексагональную форму (рис. 3.6.2; 3.6.3, см. цв. вкл.). Размеры клеток широко варьируют в зависимости от их расположения. В фовеолярной области они выше (высота 14—16 мкм), уже (10—14 мкм), чем в области зубчатой линии (ширина 60 мкм) [1046].
Клетки, лежащие по периферии, уплощены и менее пигментированы. Вблизи зубчатой линии встречаются многоядерные клетки, а зерен меланина меньше.
На момент рождения у человека обнаруживается порядка 4—6 млн клеток [496]. В процессе развития организма плотность клеток пигментного эпителия увеличивается в области желтого пятна, достигая максимума к 6 месяцам. И, наоборот, в области зубчатой линии число клеток быстро уменьшается на протяжении первого года жизни [126, 496].
С возрастом пигментные клетки в области желтого пятна увеличиваются в высоте и уменьшаются в ширине. Обратная закономерность обнаруживается по периферии сетчатки [1152]. Фигуры митотических делений в эпителиальном пласте практически не обнаруживаются.


Рис. 3.6.2. Пигментный эпителий сетчатой оболочки:
а — поперечный срез (/ — наружные членики палочек и колбочек; 2 — клетки пигментного эпителия; 3 — базальная пластинка (мембрана Бруха); 4 — собственно сосудистая оболочка); б —плоскостной препарат
Сетчатка
231
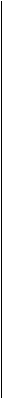 | |||||
 | |||||
 | |||||
^w^^ |

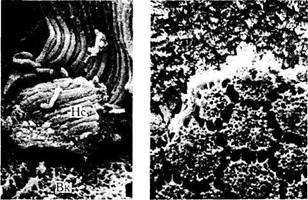
Рис. 3.6.4. Сканирующая электронная микроскопия
сетчатки (Ст) и связи ее с пигментным эпителием (Пм)
(по Kessel, Kardon, 1979):
наружные сегменты (Не) фоторецепторов контактируют с отдельными клетками пигментного эпителия (I, II). Вакуоли (Вк) в клетках пигментного эпителия появляются в результате потери зерен меланина при гистологической обработке тканей. Слева внизу показано большее увеличение участка, приведенного в рамке на верхнем снимке. Справа снизу показана базальная поверхность клеток пигментного эпителия после снятия мембраны Бруха. Между клетками виден юнкциональный комплекс в виде мостиков
Строение клеток. Как и в любых эпителиальных клетках организма человека в клетках пигментного эпителия сетчатой оболочки различают апикальную и базальную части. С ба-зальной стороны к ним прилежит базальная мембрана (рис. 3.6.5).
При световой микроскопии ткань, лежащая между пигментным эпителием и хориокапил-лярным слоем сосудистой оболочки гомогенного строения, и была названа Брухом стекловидной пластинкой (lamina vitrea), в последующем она получила название мембрана Бруха (compexus (lamina) basalis (Bruch)). При использовании более точных методов световой микроскопии в мембране Бруха выделены следующие части: наружная кутикулярная часть и более волокнистая — внутренняя часть. Поскольку внутренняя часть мембраны Бруха интенсивно окрашивается при применении мето-
Рис. 3.6.5. Особенности ультраструктурной организации клеток пигментного эпителия сетчатки и контактов между клетками:
/ — цитоплазматические отростки; 2 — юнкциональный комплекс, расположенный между соседними клетками; 3 — мембрана Бруха; 4 — соединительная ткань
дов, выявляющих эластическую ткань, ее назвали «lamina elastica». Особенности строения мембраны Бруха и ее толщина зависят как от локализации исследуемого участка, так и от возраста индивидуума. У взрослых толщина мембраны в перипапиллярной области равна 2—4 мкм, а в периферических—1—2 мкм [429]. У детей толщина ее в центральных участках равна 2 мкм.
Ультраструктурные исследования позволили выделить в мембране Бруха пять слоев (зон): базальная мембрана пигментного эпителия, внутренний коллагеновый слой, слой волокон (эластический), наружный коллагеновый слой, базальная мембрана клеток эндотелия хорио-капилляров (рис. 3.6.6—3.6.8). В действительности можно считать, что мембрана Бруха состоит только из трех внутренних слоев, поскольку наружные слои относятся к другим образованиям.
Наиболее внутренний слой мембраны, представленный базальной мембраной пигментного эпителия сетчатки, имеет толщину приблизительно 0,3 мкм. Внутренняя коллагеновая зона (толщиной 1,5 мкм) состоит из плотно упакованных и строго ориентированных фибрилл коллагена (диаметр волокон — 60 нм, а периодичность исчерченности — 64 нм). Коллаген относится, в основном, к коллагену IV типа. Волокна погружены в основное вещество, состоящее преимущественно из протеогликанов [429].
Средняя зона (эластический слой) имеет толщину порядка 0,8 мкм, и в ней эластические волокна располагаются беспорядочно. Именно в этой зоне при старении и различных патологических состояниях отмечается накопление солей кальция и липидов [502].
Наружная коллагеновая зона схожа по структуре с внутренней зоной. Единственным отличием является то, что она толще (0,7 мкм).
232
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА


Рис. 3.6.6. Объемное схематическое изображение внутреннего слоя сосудистой оболочки и пигментного эпителия сетчатки, между которыми располагается мембрана Бруха (по Hogan ei al., 1971):
1 — цитоплазматические отростки клеток пигментного эпителия;
2 — наружный сегмент палочки; 3 — запирающая лента; 4 —
десмосома; 5 — ядро клетки пигментного эпителия; 6 — мито
хондрии; 7 — комплекс Гольджи; 5 — пигментные гранулы; 9 —
фагосомы; 10 —гладкий эндоплазматический ретикулум; //—
базальная мембрана; 12 — эластическая зона мембраны Бруха;
13 — коллагеновые фибриллы мембраны Бруха; 14 — хорио-
капилляры сосудистой оболочки (стрелкой указаны поры);
15 — коллагеновые волокна, расположенные между капилляра
ми сосудистой оболочки

| 3 4 5 |
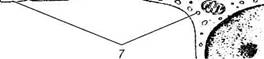
Рис. 3.6.7. Схема структурной организации мембраны Бруха (по Hogan et al., 1971):
1 — базальная мембрана пигментного эпителия сетчатки; 2 — передняя коллагеновая зона; 3 — эластический слой; 4 — наружный коллагеновый слой; 5 — базальная мембрана хориокапилляров; 6 — пигментный эпителий; 7 — эндотелиальная клетка хориокапилляров
Рис. 3.6.8. Ультраструктура мембраны Бруха:
/ — базальная мембрана клеток пигментного эпителия; 2 — внутренний коллагеновый слой мембраны Бруха толщиной 2,5 мкм; 3 — эластический слой мембраны Бруха; 4 — наружный коллагеновый слой толщиной 0,7 мкм, 5 — базальная мембрана эндоте-лиальных клеток хориокапиллярного слоя сосудистой оболочки; 6 — эндотелиальная клетка
Наиболее наружный слой мембраны Бруха, представленный базальной мембраной эндоте-лиальных клеток капилляров сосудистой оболочки, самый тонкий (0,14 мкм).
Нередко в области мембраны Бруха и клеток пигментного эпителия при офтальмоскопии можно обнаружить друзы, развивающиеся в результате процессов старения или различных заболеваний (рис. 3.6.9). Различают твердые и мягкие друзы. Они могут то появляться, то регрессировать. Твердые друзы чаще встречаются у молодых людей и являются продуктом синтетической деятельности клеток пигментного эпителия. Мягкие друзы, содержащие в своем составе мембранные структуры, отражают общие нарушения функции клеток [429, 960].
Мембрана Бруха выполняет разнообразные и важные функции, в первую очередь по избирательному транспорту питательных веществ и воды в направлении сетчатки [429]. Именно мембрана Бруха вместе с хориокапиллярным слоем сосудистой оболочки и клетками пигментного эпителия обрузует своеобразную структурно-функциональную единицу, обеспечивающую барьерные функции. Нарушение строения мембраны является причиной различных дегенеративных заболеваний пигментного эпителия (отслойка эпителия) и сенсорной части сетчатки (тапеторетинальная дегенерация, дегенерация макулярной области и др.). Способствуют этому ее возрастные изменения и формирование друз [121, 308].
Продолжая описание клеток пигментного эпителия, необходимо указать на то, что они,
Сетчатка
233

|

| «**■.: |
Рис. З.6.9. Формирование друзы во внутреннем колла-геновом слое мембраны Бруха:
/ — клетки пигментного эпителия; 2 — часть друзы, расположенной во внутреннем коллагеновом слое; 3 — наружная часть друзы, распространяющаяся на большом протяжении (стрелки)
как и другие эпителиальные клетки, в базаль-ной своей части образуют многочисленные складки. На апикальной поверхности клеток определяется множество микроворсинок, простирающихся в пространстве между наружными сегментами фоторецепторов и окутывающих их. Выделяют два типа микроворсинок. Первый тип имеет длину 5—7 мкм, а второй — 3 мкм. Микроворсинки значительно увеличивают площадь контакта клеток пигментного эпителия с фоторецепторами, способствуя тем самым высокому уровню метаболизма, благодаря увеличению интенсивности поставки питательных веществ сетчатке из хориокапиллярного слоя сосудистой оболочки и выведения из сетчатки воды, ионов и конечных продуктов метаболизма [196].
Между цитоплазматической мембраной микроворсинок эпителиоцитов и мембраной фоторецепторов никаких специализированных соединений нет и обнаруживается щелевидное пространство (рис. 3.6.10). Выполнено это пространство «цементирующей субстанцией» сложного химического состава. Называют его «интерфоторецепторный матрикс». Синтезируется он клетками пигментного эпителия. Интерфоторецепторный матрикс состоит из хондроитинсульфата (60%), сиаловой кислоты (25%) и гиалуроновой кислоты (15%) [40, 1080]. В настоящее время уточнен состав и функции этого вещества.
Первоначально предполагали, что матрикс представляет собой гомогенное скопление про-теогликанов. В настоящее время выявлено довольно сложное пространственное взаимодействие протеогликанов матрикса с наружными сегментами колбочек. Именно это взаимодействие и обеспечивает достаточно плотный контакт между пигментным эпителием и сетчаткой.

Рис. 3.6.10. Электроннограмма, иллюстрирующая характер взаимоотношения пигментного эпителия сетчатки с наружными сегментами палочек (по Hogan et al., 1971):
I — ядро клетки пигментного эпителия; 2 — митохондрии; 3 — гладкая эндоплазматическая сеть; 4 — гранулы меланина; 5 — микроворсинки, расположенные на апикальной поверхности клеток пигментного эпителия и окружающие наружные членики палочек; 6 — наружный сегмент фоторецептора
Интерфоторецепторный матрикс участвует в метаболизме сетчатки, а именно в переносе ретиноида [154, 371, 484]. Содействует он также фагоцитозу наружных сегментов фоторецепторов.
Нарушение структурной организации матрикса является немаловажной причиной возникновения отслойки сетчатки, а также сопровождает различные виды ее дегенерации.
Клетки пигментного эпителия плотно соединены между собой при помощи зон замыкания, опоясывающей десмосомы и щелевых контактов [154, 201, 513]. Органоиды опоясывают клетки с апикальной стороны, плотно скрепляя их. В средней части клеток располагаются десмосомы. Подобный контакт делает невозможным прохождение метаболитов, особенно высокомолекулярных веществ, вдоль межклеточного пространства. Этот перенос происходит только через цитоплазму клетки активным путем. Именно подобный плотный межклеточный контакт обеспечивает возможность функционирования гемато-ретинального барьера.
В разных участках пигментного эпителио-цита цитоплазма имеет отличающееся ультраструктурное строение. Именно по этой причине
234
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 цитоплазму клетки условно подразделяют на 3 зоны. Во всех зонах определяется хорошо развитый агранулярный эндоплазматический ретикулум.
цитоплазму клетки условно подразделяют на 3 зоны. Во всех зонах определяется хорошо развитый агранулярный эндоплазматический ретикулум.
Внешняя треть цитоплазмы эпителиоцитов отличается наличием большого количества митохондрий и складок базальной мембраны. Внутренняя треть цитоплазмы эпителиоцитов насыщена гранулами меланина. Видны также многочисленные свободные и связанные рибосомы. Промежуточная зона цитоплазмы относительно бедна органоидами (рис. 3.6.10). Именно здесь располагается ядро. Комплекс Гольджи выражен нечетко. Его цистерны содержат светлый материал, что свидетельствует о высокой секреторной активности клеток.
Во всех частях цитоплазмы эпителиоцитов располагаются лизосомы обычного строения. Основной их функцией является ферментативное расщепление фагоцитированных фрагментов наружных члеников фоторецепторов [109, 154, 454, 484, 501, 644, 1219, 1971].
Поскольку фагоцитарная активность клеток пигментного эпителия является одной из основных функций [185, 196, 643, 714, 826], их цитоплазма содержит фаголизосомы, образующиеся в результате слияния поглощенных наружных члеников фоторецепторов с первичной лизосомой [524, 1216].
В фаголизосоме первым подвергается лизису белковый компонент фоторецепторных дисков [306, 524, 1216].
Процесс фагоцитоза и лизиса сегментов наружных члеников фотороцепторов происходит довольно быстро. Одна клетка пигментного эпителия кролика в сутки подвергает лизису 2000 дисков в парафовеолярной области сетчатки, 3500 дисков в перифовеолярной области и почти 4000 по периферии сетчатки [484, 1216] (рис. 3.6.11, 3.6.12). Отмечено, что при интенсивном освещении количество фагосом увеличивается. Клетки пигментного эпителия отщепляют наружные членики колбочек таким же образом, как и палочек, но более интенсивно после прекращения освещения [644, 1033]. Процесс разрушения наружных члеников колбочек и палочек фоторецепторов и их утилизации является адаптивным механизмом, способствующим поддержанию структурной и функциональной целостности фоторецепторного аппарата. Тем не менее гибель фоторецепторов возникает также при различных патологических состояниях. Нередко гибель клеток происходит благодаря механизмам апоптоза, находящимся под генетическим контролем [888].

