




В последнее время проводятся интенсивные исследования роли механизмов апоптоза в развитии большой группы наследуемых дегенеративных заболеваний сетчатой оболочки. Это направление исследований имеет большое практическое значение, поскольку известно более 100 генетически наследуемых синдромов,

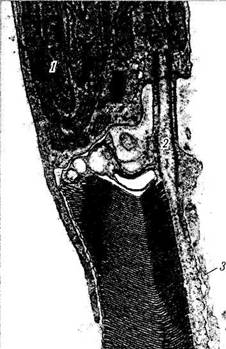
Рис. 3.6.11. Электроннограмма, иллюстрирующая стадии переваривания фрагментов наружных члеников фоторецепторов клетками пигментного эпителия:
/ — наружный членик колбочки; 2 — отделившийся фрагмент наружного членика колбочки и погруженный в цитоплазму клетки пигментного эпителия; 3 — фагосома, содержащая фрагмент наружного членика колбочки; 4 — фагосома на более поздней стадии переваривания фрагмента наружного членика; 5 — мела-носомы; 6 — митохондрии
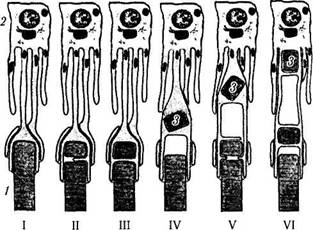
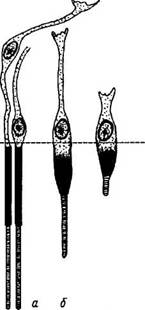
Рис. 3.6.12. Последовательные стадии (I—VI) поглощения и лизиса наружных члеников фоторецепторов пигментными эпителиоцитами сетчатой оболочки. При этом отмечается регенерация наружного членика фоторецептора:
/ — наружный членик фоторецептора; 2 — клетка пигментного эпителия; 3 — фагосома
Сетчатка
235
 сопровождающихся гибелью нейронов сетчатой оболочки. Показано, что при некоторых наследуемых синдромах механизмы апоптоза играют ведущую роль. При этом апоптоз рассматривается как конечный механизм гибели клеток, независимо от характера первичного повреждения. Основные типы повреждения фоторецепторов довольно разнообразны и сводятся к нарушению их важных функций (синтез зрительного пигмента, структуры цитоскелета клеток, последовательности процессов при восприятии световой энергии и ее трансформации в нервный импульс, фагоцитарные функции клеток пигментного эпителия и др.) [169, 886—888, 891]. Раскрытие механизмов апоптотической гибели нейронов сетчатки и участия в этом генетического аппарата рассматривается как наиболее перспективное направление в лечении этих заболеваний.
сопровождающихся гибелью нейронов сетчатой оболочки. Показано, что при некоторых наследуемых синдромах механизмы апоптоза играют ведущую роль. При этом апоптоз рассматривается как конечный механизм гибели клеток, независимо от характера первичного повреждения. Основные типы повреждения фоторецепторов довольно разнообразны и сводятся к нарушению их важных функций (синтез зрительного пигмента, структуры цитоскелета клеток, последовательности процессов при восприятии световой энергии и ее трансформации в нервный импульс, фагоцитарные функции клеток пигментного эпителия и др.) [169, 886—888, 891]. Раскрытие механизмов апоптотической гибели нейронов сетчатки и участия в этом генетического аппарата рассматривается как наиболее перспективное направление в лечении этих заболеваний.
Нередким структурным включением цитоплазмы клетки пигментного эпителия сетчатки является липофусцин.
Липофусцин содержится во многих тканях организма и его количество нарастает с возрастом. Именно по этой причине этот пигмент был назван «пигментом старения». Возникает он в результате накопления в лизосомах стареющих клеток нелизирующихся агрегатов белка и липидов [1021]. Этот пигмент отличается характерными физико-химическими свойствами, включая естественную желтовато-зеленую флюоресценцию. Накопление липофусцина происходит не только в процессе старения, но и при ряде метаболических заболеваний [1148, 1217]. Причины и механизмы возникновения ли-пофусциноза оставались загадкой более 100 лет. В настоящее время известно, что липофусцин возникает в результате перекисного окисления клеточных компонентов, особенно липидов [1210].
В глазном яблоке, как было указано выше, липофусцин обнаруживается в пигментном эпителии сетчатки [134, 258, 291, 306, 557, 562, 1159, 1176]. Максимальное его накопление происходит в клетках, расположенных в заднем полюсе. К 80 годам липофусциновые гранулы занимают до 19% объема эпителиоцитов [134, 309, 949]. В отличие от других клеток организма, в которых возникает липофусцин в результате аутофагоцитоза внутриклеточных органелл [1021], липофусцин в клетках пигментного эпителия сетчатки возникает в результате фагоцитоза наружных сегментов фоторецепторов [135, 307, 559] с последующим перекисным окислением липидной фракции этих фрагментов. В этом процессе участвует коротковолновой спектр световой энергии [440, 563].
В последнее время указывается на большую роль в формировании липофусцина в эпителиальных клетках сетчатки витамина А и его производных. Об этом свидетельствуют многочисленные экспериментальные биохимические, фи-
зикохимические исследования [291, 292, 558, 559, 561, 1148].
Зерна липофусцина необходимо морфологически отличать от меланосом. Это имеет практическое значение при диагностике пигментных новообразований. Меланиновые гранулы эпителиоцитов имеют круглую или овальную форму. При этом круглые гранулы располагаются в апикальной части клетки, а овальные — в микроворсинках. Липофусциновые гранулы круглые, но менее электронноплотные. Окрашиваются они судановыми красителями и флюоресцируют. Число зерен липофусцина прогрессивно увеличивается с возрастом. Наоборот, количество меланосом с возрастом уменьшается [309, 974, 1159, 1176]. Полагают, что уменьшение количества меланосом связано с деятельностью лизосомного аппарата клеток и возрастным измнением меланина.
Меланин клеток пигментного эпителия поглощает световую энергию достаточно широкого спектра, защищая фоторецепторы и цитоплазму пигментных эпителиоцитов от повреждающего действия света [436]. Меланин обладает свойством свободного радикала и функционирует так же, как полимер, участвующий в обмене электронов. Меланин связывает ряд металлов и лекарственных веществ.
Важно также помнить, что меланиновые гранулы пигментного эпителия сетчатки отличаются от меланосом стромальных меланоцитов уве-ального тракта. Гранулы увеального меланина значительно меньшего размера и имеют овальную форму. Это важно знать патоморфологам, особенно при дифференциальной диагностике внутриглазных пигментных новообразований.
В апикальной части, а также вблизи комплекса Гольджи клеток пигментного эпителия выявляется большое количество пиносом [812]. Размер их меньше (53 нм), чем в эндотели-альных и других клетках (более 100 нм). Эти структуры указывают на наличие интесивных процессов эндоцитоза, характерного для клеток пигментного эпителия.
В цитоплазме эпителиальных клеток можно также обнаружить дискретные темные частицы и пластинчатые тельца. Последние представляют собой фрагменты поглощенных наружных сегментов фоторецепторов [1028, 1219].
3.6.2. Сенсорная часть сетчатки
Сенсорная часть сетчатки представляет собой тонкую прозрачную оболочку, содержащую чувствительные к свету клетки, которые и превращают световую энергию в нервные импульсы. При диафаноскопии глазного яблока сетчатка выглядит пурпурно-красной из-за наличия в фоторецепторах зрительного пигмента (родопсин). Однако этот цвет быстро исчезает при освещении энуклеированного глаза на про-
236
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

 тяжении 5—10 минут. При этом сетчатка становится белой и полупрозрачной [1102].
тяжении 5—10 минут. При этом сетчатка становится белой и полупрозрачной [1102].
Толщина сетчатки в области зрительного нерва равняется 0,4 мм. Она истончается в области зубчатой линии с назальной стороны до 0,15 мм. Темпорально ее толщина 0,4 мм. В области центральной ямки (0,2 мм) [959].
Основу сенсорной части сетчатки составляют нервные клетки — фоторецептор, биполярная и ганглиозная клетки, ассоциативные горизонтальные нейроны, амакриновые клетки, а также глиальные элементы — клетка Мюллера, фиброзные и протоплазматические астроциты, микроглия и олигодендроциты.
Фоторецепторы (палочки и колбочки). Слой палочек и колбочек является самым наружным слоем сенсорной сетчатки. Складывается он из цитоплазматических выростов палочек и колбочек фоторецепторных клеток. Фоторецепторы являются не чем иным, как высокоспециализированными нейроэпителиаль-ными клетками. По структуре и направленности выполняемой функции они близки к ре-цепторным клеткам других тканей и органов (тельца Пачини, Краузе, Мейснера).
Тела фоторецепторных клеток располагаются в плоскости наружной пограничной мембраны, а их апикальные отростки (внутренние сегменты) лежат только снаружи этой мембраны.
Большое значение имеет знание распределения и пространственной ориентации фоторецепторных клеток, что в значительной мере способствует пониманию зрительных связей в сетчатке. Плотное расположение фоторецепторов и их точная ориентация вдоль зрительной оси обеспечивают детальный анализ поля зрения. Любое изменение расположения фоторецепторов приводит к нарушению зрения. Если между фоторецепторами появляются пространства (при центральной серозной ретинопатии) и они неравномерно распределены, развивается мик-ропсия. Нарушение ориентации фоторецепторов вдоль зрительной оси приводит к метамор-фопсии.
Фоторецепторы распределяются закономерным образом, в виде мозаики (рис. 3.6.13). В области желтого пятна лежат только колбочки. Вне желтого пятна колбочки кольцевидно окружены палочками.
В сетчатой оболочке обнаруживается от 77,9 до 107,3 млн (в среднем 92 млн) палочек и 4,08—5,29 млн (в среднем 4,6 млн) колбочек.
Существуют индивидуальные отличия плотности палочек и колбочек в зависимости от топографического отдела сетчатки [223]. Наибольшее разнообразие плотности выявляется вблизи центральной ямки и у зубчатой линии, а наименьшее — в средней части сетчатки и по периферии.
Плотность колбочек максимальна в области центральной ямки (199 000 колбочек в мм2). При этом их число колеблется в широких пре-
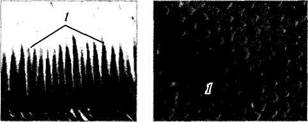

Рис. 3.6.13. Особенности «мозаичного» строения периферии сетчатки (а) и области центральной ямки (б):
/ — палочки; 2 — колбочки. Слева иллюстрируется срез сетчатки, а справа — плоскостной препарат (по Curcio et al., 1990)
делах (от 100 000 до 324 000 колбочек в мм2) [223]. По мере удаления от центральной ямки плотность колбочек существенно уменьшается. Так, плотность колбочек уменьшается до 75 000 мм2 в 130 мкм от центра центральной ямки. Примерно в трех миллиметрах от центра центральной ямки отмечается наибольшая плотность палочек, а плотность колбочек уменьшается. Степень этого уменьшения различна в зависимости от направления. Так, плотность колбочек с назальной стороны на 40— 45% выше, чем с темпоральной. В периферических отделах сетчатки плотность колбочек опять возрастает (рис. 3.6.13—3.6.15).
Считают, что пространственное расположение колбочек в области желтого пятна является фактором, определяющим разрешающую способность глаза. Так, среднее расстояние между центрами колбочек колеблется от 2,53 ±0,29 мкм до 6,16 ±1,04 мкм. Наименьшее расстояние между клетками обнаружено в области центральной ямки. Это свидетельствует о наибольшей разрешающей способности сетчатки именно в этой области [223].
Необходимо отметить, что данные психофизиологических исследований относительно остроты зрения не полностью совпадают с приведенными выше анатомическими данными. По всей видимости, большое значение имеют другие факторы [1171]. Единственная область в сетчатке, где функциональная острота зрения совпадает с анатомической разрешающей способностью, располагается между 0,2 и 2,0°. Интересно, что острота зрения у новорожденных на два порядка ниже, чем у взрослых [131]. В ближайшее время после рождения колбочки,
Сетчатка
237
 Количество палочек и колбочек на мм 2 Пик колбочек Диск зрительного нерва
Количество палочек и колбочек на мм 2 Пик колбочек Диск зрительного нерва
 |
Пик палочек |
| Колбочки |
 180 000
180 000
160 000
140 000
120 000
100 000 80 000 -60 000 -40 000 -20 000 -
0
70 60 50 40 30 20 10 0 Темпоральная
| Колбочки |
\

 10 20 30 40 50 60 70 80 90 Назальная
10 20 30 40 50 60 70 80 90 Назальная
Рис. 3.6.14. Плотность палочек и колбочек вдоль горизонтального меридиана (по Osterberg, 1935)
|
|
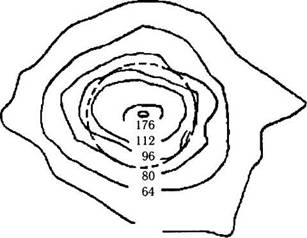
| 44 |
Рис. 3.6.15. Топографические особенности распределения плотности колбочек в области центральной ямки (по Curcio et al., 1987):
контурные сплошные линии очерчивают области с количеством колбочек в одном квадратном миллиметре, равном цифре, приведенной на рисунке и умноженной на 1000. Окружность (пунктирная линия) очерчивает поле зрения, равное 1 градусу
палочки и клетки пигментного эпителия перемещаются к центру желтого пятна. При этом дифференциация фоторецепторов в центре сетчатки происходит медленнее, чем по периферии [474]. Изучение сетчатки обезьян показало, что плотность колбочек, свойственная взрослым животным, появляется только к 15—18 месяцам после рождения [813]. У человека плотность колбочек нарастает вплоть до 5—8-летнего возраста [474]. Наиболее важным фактором, определяющим низкую остроту зрения
у новорожденных, является не плотность расположения колбочек, а неполная дифференциация желтого пятна [224]. Косвенным подтверждением этому является альбинизм. У этих больных острота зрения низкая, а желтое пятно в структурном отношении напоминает желтое пятно новорожденного [1172].
Сниженная острота зрения у этих больных связана также с недостаточностью развития межнейронных связей на уровне наружного коленчатого тела и зрительной коры головного мозга [535].
Плотность палочек и их распределение также являются объектом пристального внимания исследователей. Установлено, что диаметр свободной от палочек области желтого пятна равняется 0,35 мм. Это соответствует 1,25 градуса поля зрения (рис. 3.6.14, 3.6.15) [154]. Самая высокая концентрация палочек выявлена в области сетчатки, имееющей вид горизонтального эллипса. Этот эллипс несколько расширяется в носовом направлении и кверху. Именно от этого места плотность палочек медленно уменьшается по мере продвижения к периферии сетчатки.
С назальной стороны плотность палочек на 20—25% выше, чем с височной стороны. В верхней половине сетчатки палочек больше на 2%, чем в нижней половине. Равное соотношение палочек и колбочек обнаруживается на расстоянии 0,5 мм кнутри и на 0,4 мм выше центральной ямки [223].
Внутренние и наружные сегменты фото рецепторов. Внутренние и наружные сегменты фоторецепторов являются местом трансформации световой энергии в нервный импульс. Они
238
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

|

имеют следующее строение. От тела фото-рецепторной клетки отходит цитоплазматичес-кий вырост. Этот вырост подразделяется на две части — внутренний и наружный сегменты (рис. 3.6.16—3.6.19). Наружный сегмент лежит

Рис. 3.6.16. Схематическое изображение особенностей Рис. 3.6.18. Ультраструктурные различия наружных
| и внутренних члеников палочек и колбочек (по Kolb et al., 1998): I —палочки; //—колбочки; /—диски; 2 — наружный сегмент; 3 — внутренний сегмент |
строения колбочки и палочки:
|
|
 / — наружный плексиформный слой; 2 — наружный ядерный слой; 3 — наружная пограничная мембрана; 4 — внутренний сегмент; 5 — наружный сегмент; б — синаптическое тело; 7 — ядра; 8 —миоидная часть; 9 — эллипсоидная часть; 10 — диски фоторецепторов; //— пигментный эпителий
/ — наружный плексиформный слой; 2 — наружный ядерный слой; 3 — наружная пограничная мембрана; 4 — внутренний сегмент; 5 — наружный сегмент; б — синаптическое тело; 7 — ядра; 8 —миоидная часть; 9 — эллипсоидная часть; 10 — диски фоторецепторов; //— пигментный эпителий
Наружная
пограничная
мембрана
Внутренний сегмент
| в |
Наружный сегмент
Рис. 3.6.17. Топографические особенности строения
фоторецепторов сетчатки человека (по Tripathi et al.,
| Рис. 3.6.19. Электроннограмма наружного и внутреннего члеников палочки (по Hogan et al., 1971): 1 —митохондрии; 2 —ресничка; 3 —цитоплазматические отростки внутреннего сегмента, окутывающие наружный членик |
1984):
а — колбочки из области центральной ямки; б — колбочки в области сетчатки, лежащей между зубчатой линией и диском зрительного нерва; в — колбочка области зубчатой линии; г — палочка
Сетчатка
239
 в интерфоторецепторном матриксе и обращен к апикальной поверхности клеток пигментного эпителия. Основной функцией этого образования является преобразование световой энергии в электрические импульсы. В дальнейшем, нервные импульсы обрабатываются на уровне сетчатки и передаются по зрительному нерву коре головного мозга. Восприятие света и преобразование его в нервный импульс начинаются с активации последовательных реакций фотохимической стереоизомеризации зрительного пигмента, расположенного на дисках наружного сегмента фоторецепторов. К зритель ным пигментам относятся родопсин и иодо-псин. Родопсин, обнаруживаемый в палочках, обеспечивает фотопическое зрение.
в интерфоторецепторном матриксе и обращен к апикальной поверхности клеток пигментного эпителия. Основной функцией этого образования является преобразование световой энергии в электрические импульсы. В дальнейшем, нервные импульсы обрабатываются на уровне сетчатки и передаются по зрительному нерву коре головного мозга. Восприятие света и преобразование его в нервный импульс начинаются с активации последовательных реакций фотохимической стереоизомеризации зрительного пигмента, расположенного на дисках наружного сегмента фоторецепторов. К зритель ным пигментам относятся родопсин и иодо-псин. Родопсин, обнаруживаемый в палочках, обеспечивает фотопическое зрение.
Фотопическое зрение происходит в колбочках и обеспечивается трихроматическими пигментами. Колбочки содержат одну из трех молекул иодопсина, поглощающих свет в трех различных спектрах — 440 нм (синий), 540 нм (зеленый) и 577 нм (оранжевый). Обозначаются эти колбочки как S- (коротковолновые), М- (средневолновые) и L- (длинноволновые) [229]. Более подробные сведения о химических процессах, происходящих в фоторецепторных клетках в процессе формирования нервного импульса будут приведены в разделе «Зрительные пигменты и фоторецепция».
Для понимания механизмов цветового зрения большое значение имеет морфологическая дифференциация различных типов колбочек. Это необходимо для определения их связи с другими нейронами сетчатки. В настоящее время морфологическими методами дифференцируют средне- и длинноволновые колбочки у некоторых рыб, лягушек, птиц и рептилий. К сожалению, у приматов и человека возможна только дифференциация коротковолновых колбочек (S-колбочка) от отстальных. У «синей» колбочки более длинный и больший диаметр внутреннего членика, который интенсивно окрашивается [44, 221, 1058]. Кроме того, «синие» колбочки не столь равномерно и закономерно распределены в сетчатой оболочке. Они составляют 3—5% от общего числа фоторецепторов в центре желтого пятна, и их число увеличивается до 15% на склоне области желтого пятна. Использование антител, обладающих аффинитетом к синему опсину, подтвердило то, что «синие» колбочки в области желтого пятна редки и лежат изолированно или полностью отсутствуют в зоне, расположенной недалеко от пика наибольшей плотности колбочек. Диаметр этой зоны равен 100 мкм (0,35 градуса) [224]. Самая высокая плотность «синих» колбочек (более чем 2000 клеток в мм2) выявлена в зоне шириной 0,1—0,3 мм.
Наружные членики колбочек и палочек являются результатом выпячивания плазматической мембраны фоторецептора. Наружный членик соединяется с внутренним сегментом по-
средством цитоплазматического перешейка. Общая длина обоих сегментов определяется локализацией и типом фоторецептора.
Наружный членик постоянно обновляется. Этот процесс иллюстрируется рис. 3.6.11, 3.6.12. При этом постоянно регенерируют и зрительные пигменты. Опсиновая часть молекулы родопсина синтезируется аппаратом Гольд-жи фоторецепторной клетки [250, 371, 816]. Другая составная часть зрительного пигмента (ретинал — производное витамина А) поставляется дискам наружных члеников клетками пигментного эпителия сетчатки при помощи транспортной молекулы [42, 371].
Внутренние и наружные членики палочек имеют длину 40—60 мкм на всем протяжении сетчатки. Длина сегмента колбочки максимальна в области желтого пятна (80 мкм) и постепенно уменьшается до 40 мкм к периферии сетчатки. В области зубчатой линии колбочки короче (4 мкм) и толще. Наружный сегмент палочки (длина 25—28 мкм и диаметр 1 —1,5 мкм) не изменяется на протяжении всей сетчатки.
Необходимо отметить, что колбочки в области желтого пятна напоминают по форме и размеру палочки. Наружные сегменты колбочек вдали желтого пятна имеют диаметр 6 мкм в основании и 1,5 мкм на верхушке [154, 1102].
Наружные членики (сегменты) палочек имеют цилиндрическую форму и содержат плотно упакованные двойные дисковидные пластины, количество которых колеблется от 600 до 1000 (рис. 3.6.19). Каждый диск имеет толщину 22,5—24,5 мкм. Расстояние между дисками равно 21 мкм [1008]. Никаких специализированных контактов между дисками, а также дисками и цитоплазматической мембраной не обнаруживается. Наружная поверхность каждого наружного сегмента покрыта слоем нейрокератана.
В дисках содержится до 90% молекул зрительного пигмента. Остальное количество его рассеяно по поверхности плазматической мембраны. Наружный и внутренний сегменты соединяет модифицированная ресничка [1008]. Именно в месте перехода наружного сегмента во внутренний сегмент цитоплазма суживается. Ширина этого перешейка равна 0,3 мкм, а длина 1 мкм. В основании внутреннего сегмента лежит базальное тельце, состоящее из одной пары центриолей. Ресничка состоит из девяти пар микротрубочек, расположенных кольцевидно. Пучки филаментов исходят из базального тельца и простираются поперек эллипсоидной части цитоплазмы, заканчиваясь в миоидной области цитоплазмы (рис. 3.6.19). В фибриллах ресничек высока активность АТФ-азы, свидетельствующая об интенсивном метаболизме этого образования. Правда, функции ресничек пока неизвестны.
Цитоплазма поверхности наружного сегмента формирует 9—12 микроворсинок, длиной
240
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА


 12,5 мкм. Функция микроворсинок пока неизвестна.
12,5 мкм. Функция микроворсинок пока неизвестна.
Внутренние членики (сегменты) палочек имеют цилиндрическую форму. Гистологически различают две части внутреннего членика: эозинофильную наружную, называемую эллипсоидной частью, и внутреннюю базофильную, называемую миоидной частью (рис. 3.6.16— 3.6.19). Тинкториальные свойства этих двух областей изменяются в зависимости от метаболической активности фоторецептора. Эллипсоидная часть окрашивается эозинофильно в связи с наличием в ней большого количества митохондрий. В одной палочке можно найти до 600 митохондрий. Цитоплазма также содержит гладкий эндоплазматический ретикулум, нейро-трубочки, свободные рибосомы и гранулы гликогена. Базофилия миоидной части зависит от большой концентрации в ней свободных рибосом. Миоидная область является центром белкового синтеза. По этой причине в ней располагаются шероховатая эндоплазматическая сеть, аппарат Гольджи, микротрубочки, микрофила-менты и гранулы гликогена. Таким образом, основной функцией этой части фоторецептора является обеспечение метаболических и синтетических функций клетки.
Наружные сегменты (членики) колбочек имеют различное строение в различных участках сетчатки. В области зубчатой линии и по периферии сетчатки они короткие и конические, а в fovea centralis продолговатые, напоминают наружные сегменты палочек (рис. 3.6.16—3.6.18).
Ультраструктурными исследованиями установлено, что наружный сегмент колбочки имеет больше дисков (1000—1200), чем наружный сегмент палочки. Междисковые пространства палочки более широкие (в колбочках — 3,5 мкм, в палочках— 16,5 мкм).
В отличие от дисков палочек диски колбочек соединены между собой и прикрепляются к плазматической мембране.
Внутренние сегменты (членики) колбо чек. Наружные и внутренние членики колбочек связаны друг с другом посредством тонкого цитоплазматического перешейка, содержащего видоизмененную ресничку. Они изменяются в зависимости от их топографического расположения. В центральной ямке (fovea centralis) они более длинные и узкие. Ультраструктурная организация внутреннего сегмента палочек и колбочек одинаковая, за исключением того, что в колбочках значительно больше митохондрий (200—300 на срезе).
Наружная поверхность миоидной части палочек и колбочек покрыта волосоподобными цитоплазматическими отростками мюллеровс-ких клеток, формирующих «корзины Шульца». Благодаря этому никакого контакта между смежными клетками нет [1008]. Отростки мюл-леровских клеток участвуют также в регуляции состава внеклеточной среды фоторецепто-
ров и служат для жесткой пространственной фиксации палочек и колбочек.
Наружная пограничная мембрана. При световой микроскопии видно, что наружная пограничная мембрана (рис. 3.6.1) отделяет слой палочек и колбочек от подлежащего наружного ядерного слоя сетчатки. Она простирается от диска зрительного нерва до зубчатой линии, где превращается в базальную пластинку, расположенную между пигментированными и беспигментными частями ресничного эпителия. Наружная пограничная мембрана представляет собой не что иное, как скопление в одной плоскости терминальных пластинок (zonulae adhe- rentes), расположенных между мюллеровскими клетками и фоторецепторами, между смежными мюллеровскими клетками и, редко, между соседними фоторецепторами.
Наружная пограничная мембрана, таким образом, не является истинной мембраной. Через нее проходят небольшие молекулы. Главной функцией мембраны является обеспечение функционирования избирательного барьера на пути питательных веществ, которые проходят между рядом расположенными мюллеровскими клетками, а также стабилизация положения фоторецепторов.
Наружный ядерный слой. Наружный ядерный слой находится кнутри от наружной пограничной мембраны и содержит тела и ядра фото-рецепторных клеток (рис. 3.6.1). В зависимости от участка сетчатки ширина этого слоя изменяется, прежде всего, из-за изменения числа рядов ядер.
С назальной стороны диска наружный ядерный слой имеет толщину 45 мкм и состоит из 8—9 рядов ядер. С височной стороны он состоит только из четырех рядов ядер, истончаясь до 22 мкм. В желтом пятне наличие 10 рядов ядер колбочек увеличивают ширину наружного ядерного слоя до 50 мкм. В области зубчатой линии наружный ядерный слой состоит только из одного слоя ядер колбочковых клеток, которые плотно прилежат к наружной пограничной мембране и четырем рядам ядер палочек, расположенным кнутри от них. Толщина ядерного слоя при этом приблизительно 27 мкм.
Ядра колбочек овальные и имеют диаметр 5—7 мкм. Расположены они на 3—4 мкм кнутри от наружной пограничной мембраны. Ядра палочек также овальные, диаметром 5,5 мкм.
Цитоплазма обоих типов клеток скудная. Тела палочек и колбочек окрашиваются по-разному. При использовании метода Унна тело палочек не окрашивается, а колбочки окрашиваются в интенсивно синий цвет. Используя трехцветный метод Маллори, после фиксации сетчатки жидкостью Ценкера можно четко дифференцировать центральную ямку. Центральная ямка окрашена в интенсивно красный цвет. Это связано с тем, что методом Маллори окрашиваются только колбочки.
Сетчатка
241
 Наружный сетчатый (плексиформный) слой (рис. 3.6.1) является местом соединения первого и второго нейронов, т. е. местом передачи информации от первого нейрона (фоторецептора) второму (биполярной клетке). Помимо указанных клеток в нем располагаются ассоциативные нейроны (горизонтальная клетка).
Наружный сетчатый (плексиформный) слой (рис. 3.6.1) является местом соединения первого и второго нейронов, т. е. местом передачи информации от первого нейрона (фоторецептора) второму (биполярной клетке). Помимо указанных клеток в нем располагаются ассоциативные нейроны (горизонтальная клетка).

