




С возрастом развиваются структурные изменения дренажной системы, увеличивающие сопротивление оттоку камерной влаги и способствующие развитию глаукомного процесса [20]. Степень структурных изменений дренажной системы коррелирует со степенью изменений сосудов организма при общих сосудистых заболеваниях. Это отмечено при атеросклерозе, гипертонической болезни, сахарном диабете и др. [3, 4, 5, 20].
При старении в два-три раза утолщаются трабекулы, главным образом в результате накопления спиралевидного коллагена. Увеличивается количество базального материала. Однако количество протеогликанов (хондроитинсуль-фат) уменьшается [387]. Исчезает микрофибриллярный компонент эластических волокон
204
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 [1103]. Показано, что в процессе физиологического старения в дренажной зоне глаза происходят незначительные нарушения в виде му-коидного набухания. Эти изменения могут привести к нарушению оттока камерной влаги, но глаукома не развивается, так как гомеостати-ческие механизмы, обеспечивающие поддержание внутриглазного давления на физиологическом уровне, компенсируют этот сдвиг.
[1103]. Показано, что в процессе физиологического старения в дренажной зоне глаза происходят незначительные нарушения в виде му-коидного набухания. Эти изменения могут привести к нарушению оттока камерной влаги, но глаукома не развивается, так как гомеостати-ческие механизмы, обеспечивающие поддержание внутриглазного давления на физиологическом уровне, компенсируют этот сдвиг.
Рядом авторов выявлены дегенеративные изменения трабекулярных клеток, число которых прогрессивно снижается. Слой клеток истончается, трабекулы «сливаются». Этот процесс расценивают как «гиалиноз» трабекулярного аппарата [61, 389, 404, 727, 1097, 1098, 1205], который приводит к увеличению сопротивляемости оттоку камерной влаги и повышению внутриглазного давления. Отмечается и уменьшение числа клеток в юкстаканаликуляр-ной ткани. В ней накапливается материал, являющийся продуктом распада эластических волокон и других молекул типа спиралевидного коллагена. Содержание в этой области гиалуро-новой кислоты с возрастом также снижается [386, 580]. Биохимическими исследованиями показано увеличение количества фибронектина, коллагена VI типа и тромбоспондина. При этом уменьшается количество ламинина [154, 738], который, тем не менее, в повышенном количестве обнаружен под эндотелиальной выстилкой шлеммова канала [697].
Приведенные выше изменения выявлены и при развитии первичной открытоугольной глаукомы [59, 386, 1097, 1098].
Трабекулярная сеть при глаукоме. Наиболее ранние изменения трабекулярного аппарата при открытоугольной глаукоме пока не установлены. Исследование участков трабекулярного аппарата, удаленного во время операции на поздних стадиях глаукомы, позволило Rohen и Witmer [920] выявить материал в виде «бляшки», располагающийся в сетчатой части тра-бекулярной сети и под эндотелиальными клетками шлеммова канала. Они различают 3 типа «бляшек». Первый тип «бляшек» преимущественно располагается у шлеммова канала и состоит из гомогенного или мелкозернистого материала. «Бляшки» второго типа выглядят на тангенциальных срезах в виде точек. При элект-ронномикроскопическом исследовании они представляют собой центральные участки эластопо-добных волокон, разрезанных поперек. «Бляшки» третьего типа состоят из электронноплот-ного материала, содержащего зернистый компонент и исчерченные фибриллы [671, 676, 907].
Количество всех трех типов «бляшкоподоб-ного» материала увеличивается с возрастом, а при открытоугольной глаукоме количество этого материала значительно больше независимо от возраста больного [58, 79, 676, 899]. Накопление «бляшкоподобного» материала может являться препятствием на пути оттока камер-
ной влаги, особенно при локализации его вблизи эндотелиальных клеток шлеммова канала.
Микроскопически также выявлено, что при открытоугольной глаукоме возможно спадение наружной и внутренней стенок шлеммова канала. При этом отсутствует эндотелиальная выстилка. Сочетались эти изменения со значительным скоплением «бляшкоподобного» материала.
При открытоугольной глаукоме нередко обнаруживаются также признаки воспаления, проявляющиеся инфильтрацией трабекулярной сети лимфоцитами [59, 60, 403, 416]. Подобная инфильтрация выявляется только на поздних стадиях развития заболевания.
При глаукоме уменьшается также количество трабекулярных клеток [403, 416]. Уменьшение количества трабекулярных клеток сопровождается появлением в сохранившихся клетках так называемых матричных пузырьков, представляющих собой морофлогическую форму лизосом [915]. Прогрессивное уменьшение количества трабекулярных клеток может стать причиной «слипания» трабекул между собой [403].
Нередко при глаукоме наступает гиалиноз корнеосклеральных и увеальных трабекул. Этому, как правило, предшествует накопление ба-зальноподобного материала. Подобные изменения довольно сильно напоминают возрастные, что ряду авторов дает основание предполагать наличие единых механизмов, лежащих в основе старения и возникновения открытоугольной глаукомы [20]. Подтверждении тому являются данные о нарушении процессов пере-кисного окисления, как при старении, так и при глаукоме [831, 834]. Показано первичное повреждение продуктами перекисного окисления клеточных мембран эндотелиальных клеток, что может явиться пусковым механизмом развития сосудистых заболеваний глаза и глаукомы.
3.4. ХРУСТАЛИК И РЕСНИЧНЫЙ ПОЯСОК (ЗОНУЛЯРНЫЙ АППАРАТ)
Особое внимание строению хрусталика уделялось на самых ранних этапах микроскопии. Именно хрусталик впервые исследован микроскопически Левенгуком, который указал на его волокнистую структуру.
Хрусталик
Форма и размер. Хрусталик (Lens) представляет собой прозрачное, двояковыпуклое в виде диска, полутвердое образование, расположенное между радужкой и стекловидным телом (рис. 3.4.1, см. цв. вкл.).
Хрусталик уникален тем, что он является единственным «органом» тела человека и большинства животных, состоящим из одного типа
Хрусталик и ресничный поясок (зонулярный аппарат)
205

 клеток на всех стадиях — от эмбрионального развития и постнатальной жизни вплоть до смерти. Существенным его отличием является отсутствие в нем кровеносных сосудов и нервов. Уникален он и в отношении особенностей метаболизма (преобладает анаэробное окисление), химического состава (наличие специфических белков — кристаллинов), отсутствия толерантности организма к его белкам. Большинство этих особенностей хрусталика связано с характером эмбрионального его развития, о чем будет сказано несколько ниже.
клеток на всех стадиях — от эмбрионального развития и постнатальной жизни вплоть до смерти. Существенным его отличием является отсутствие в нем кровеносных сосудов и нервов. Уникален он и в отношении особенностей метаболизма (преобладает анаэробное окисление), химического состава (наличие специфических белков — кристаллинов), отсутствия толерантности организма к его белкам. Большинство этих особенностей хрусталика связано с характером эмбрионального его развития, о чем будет сказано несколько ниже.
Передняя и задняя поверхности хрусталика соединяются в так называемой экваториальной области. Экватор хрусталика открывается в заднюю камеру глаза и при помощи цинновой связки (ресничный поясок) присоединен к ресничному эпителию (рис. 3.4.2). Благодаря расслаблению цинновой связки при сокращении


Рис. 3.4.2. Соотношение структур переднего отдела глаза (схема) (по Rohen; I979):
а — срез, проходящий через структуры переднего отдела глаза (/ — роговая оболочка; 2 — радужная оболочка; 3 — ресничное тело; 4 — ресничный поясок (циннова связка); 5 — хрусталик); б — сканирующая электронная микроскопия структур переднего отдела глаза (/ — волокна зонулярного аппарата; 2 — ресничные отростки; 3 — ресничное тело; 4 — хрусталик; 5 — радужка; 6 — склера; 7 — шлеммов канал; 8 — угол передней камеры)
ресничной мышцы происходит деформация хрусталика (увеличение кривизны передней и, в меньшей степени, задней поверхностей). При этом выполняется основная его функция — изменение рефракции, позволяющее на сетчатке получить четкое изображение независимо от расстояния до предмета. В покое без аккомодации хрусталик дает 19,11 из 58,64 дптр преломляющей силы схематического глаза. Для выполнения своей основной роли хрусталик должен быть прозрачным и эластичным, каковым он и является.
Хрусталик человека растет непрерывно на протяжении всей жизни, утолщаясь примерно на 29 мкм в год [158, 785]. Начиная с 6—7-й недели внутриутробной жизни (18 мм эмбриона) он увеличивается в передне-заднем размере в результате роста первичных хрусталиковых волокон. На стадии развития, когда эмбрион достигает размера в 18—24 мм, хрусталик имеет приблизительно сферическую форму. С появлением вторичных волокон (размер эмбриона 26 мм) хрусталик уплощается и его диаметр увеличивается. Зонулярный аппарат, появляющийся при длине эмбриона 65 мм, не влияет на увеличение диаметра хрусталика. В последующем хрусталик быстро увеличивается в массе и объеме. При рождении он имеет почти сферическую форму.
В первые два десятилетия жизни увеличение толщины хрусталика прекращается, но продолжает увеличиваться его диаметр. Фактором, способствующим увеличению диаметра, является уплотнение ядра. Натяжение цинновой связки способствует изменению формы хрусталика [157].
Диаметр хрусталика (измеренный по экватору) взрослого человека равен 9—10 мм. Толщина его на момент рождения в центре равна приблизительно 3,5—4,0 мм, 4 мм в 40 лет, а затем медленно увеличивается до 4,75—5,0 мм к старческому возрасту. Толщина изменяется и в связи с изменением аккомодационной способности глаза.
В отличие от толщины экваториальный диаметр хрусталика с возрастом изменяется в меньшей степени. При рождении он равняется 6,5 мм, на втором десятилетии жизни — 9— 10 мм. В последующем он практически не меняется (табл. 3.4.1).
Передняя поверхность хрусталика менее выпуклая, чем задняя (рис. 3.4.1). Она представляет собой часть сферы с радиусом кривизны, равным в среднем 10 мм (8,0—14,0 мм). Передняя поверхность граничит с передней камерой глаза посредством зрачка, а по периферии с задней поверхностью радужки. Зрачковый край радужки опирается на переднюю поверхность хрусталика. Боковая поверхность хрусталика обращена в сторону задней камеры глаза и посредством цинновой связки присоединяется к отросткам ресничного тела.
206
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 Таблица 3.4.1. Размеры хрусталика (по Rohen, 1977)
Таблица 3.4.1. Размеры хрусталика (по Rohen, 1977)
Сагиттальный диаметр (толщина), мм
новорожденный.......................................................... 3,5
10 лет............................................................................ 3,9
20—50 лет........................................................ 4,0—4,14
60—70 лет.................................................................. 4,77
80—90 лет.................................................................... 5,0
Экваториальный диаметр, мм
новорожденный.............................................................. 6,5
после 15 лет................................................................ 9,0
Вес, мг
новорожденный............................................................... 65
первый год жизни..................................................... 130
20—30 лет.................................................................. 174
40—50 лет.................................................................. 204
90 лет............................................................................ 250
Объем, мл
30—40 лет................................................................ 0,163
80—90 лет................................................................ 0,244
Толщина капсулы, мкм
передний полюс................................................... 8—14
экватор.................................................................... 7—17
задний полюс........................................................... 2—4
Хрусталиковые волокна
длина, мм................................................................ 8—12
толщина, мм................................................................. 4,6
количество................................................... 2100—2300
Центр передней поверхности хрусталика называют передним полюсом. Располагается он примерно на расстоянии 3 мм позади задней поверхности роговой оболочки.
Задняя поверхность хрусталика обладает большей кривизной (радиус кривизны равен 6 мм (4,5—7,5 мм)). Ее обычно рассматривают в комплексе со стекловидной мембраной передней поверхности стекловидного тела. Тем не менее между этими структурами существует щелеподобное пространство, выполненное жидкостью. Это пространство позади хрусталика
было описано еще Бергером (Berger) в 1882 году. Его можно наблюдать при использовании щелевой лампы.
Экватор хрусталика лежит в пределах ресничных отростков на расстоянии от них в 0,5 мм. Экваториальная поверхность неровная. Она обладает многочисленными складками, образование которых связано с тем, что к этой области прикрепляется цинновая связка. Складки исчезают при аккомодации, т. е. при прекращении натяжения связки.
Коэффициент преломления хрусталика равен 1,39, т.е. несколько больший, чем коэффициент преломления камерной влаги (1,33). Именно по этой причине, несмотря на меньший радиус кривизны, оптическая сила хрусталика меньше, чем роговой оболочки. Вклад хрусталика в рефракционную систему глаза равен приблизительно 15 из 40 диоптрий.
При рождении аккомодационная сила, равная 15—16 диоптриям, уменьшается наполовину к 25 годам, а в возрасте 50 лет равна лишь 2 диоптриям.
При биомикроскопическом исследовании хрусталика с расширенным зрачком можно обнаружить особенности его структурной организации (рис. 3.4.3). Во-первых, выявляется мно-гослойность хрусталика. Различаются следующие слои, считая спереди к центру: капсула; подкапсулярная светлая зона (кортикальная зона С 1а); светлая узкая зона неоднородного рассеивания (С1); полупрозрачная зона коры (С2). Перечисленные зоны и составляют поверхностную кору хрусталика. Существует еще две более глубоко расположенные зоны коры. Их называют еще пернуклеарными. Эти зоны флюоресцируют при освещении хрусталика синим светом (СЗ и С4).

а 6 6
Рис. 3.4.3. Послойность строения хрусталика при биомикроскопическом его исследовании у индивидуумов различного возраста (по Bron et al., 1998):
а — возраст 20 лет; б — возраст 50 лет; s — возраст 80 лет (/ — капсула; 2 — первая кортикальная светлая зона (С1 альфа); 3 — первая зона разобщения (С1 бета); 4 — вторая кортикальная светлая зона (С2): 5 — рассеивающая свет зона глубокой коры (СЗ); 6 — светлая зона глубокой коры; 7 — ядро хрусталика. Отмечается увеличение хрусталика и усиление рассеивания света
Хрусталик и ресничный поясок (зонулярный аппарат)
207

|

| • ««;■ '4 -,,..■ |
Ядро хрусталика рассматривают как его пре-натальную часть. Оно также обладает слоистостью. В центре располагается светлая зона, называемая «зародышевым» (эмбриональным) ядром. При исследовании хрусталика с помощью щелевой лампы также можно обнаружить швы хрусталика. Зеркальная микроскопия при большой кратности увеличения позволяет увидеть эпителиальные клетки и волокна хрусталика.
Определяются следующие структурные элементы хрусталика (рис. 3.4.4—3.4.6):
1. Капсула.
2. Эпителий.
3. Волокна.
Капсула хрусталика (capsula lentis). Хрусталик со всех сторон покрыт капсулой, которая является не чем иным, как базальной мембраной эпителиальных клеток. Капсула хрусталика самая толстая базальная мембрана тела чело-

Рис. 3.4.4. Схема микроскопического строения хрусталика:
/ — капсула хрусталика; 2 — эпителий хрусталика центральных участков; 3 — эпителий хрусталика переходной зоны; 4 — эпителий хрусталика экваториальной области; 5 — эмбриональное ядро; 6 — фетальное ядро; 7 — ядро взрослого; 8 — кора
Рис. 3.4.6. Особенности ультраструктуры капсулы
хрусталика экваториальной области, цинновой связки
и стекловидного тела:
/ — волокна стекловидного тела; 2 — волокна цинновой связки; 3 —прекапсулярные волокна; 4 —капсула хрусталика
века. Спереди капсула толще (15,5 мкм спереди и 2,8 мкм — позади) [798] (рис. 3.4.7). Более выражено утолщение по периферии передней капсулы, поскольку в этом месте прикрепляется основная масса цинновой связки. С возрастом толщина капсулы увеличивается, что более выражено спереди [13, 321, 798, 959]. Это связано с тем, что эпителий, являющийся источником базальной мембраны, расположен спереди и участвует в ремодуляции капсулы, отмечаемой по мере роста хрусталика.
14 мкм
 21 мкм
21 мкм
| 23 мкм |
17 мкм

Рис. 3.4.5. Особенности строения экваториальной области хрусталика (по Hogan et al., 1971):
I — капсула хрусталика; 2 — экваториальные эпителиальные клетки; 3 — хрусталиковые волокна. По мере пролиферации эпителиальных клеток, расположенных в области экватора хрусталика, они смещаются к центру, превращаясь в хрусталиковые волокна
4 мкм
Рис. 3.4.7. Толщина капсулы хрусталика в различных зонах
Способность эпителиальных клеток к кап-сулообразованию сохраняется на протяжении всей жизни [17] и проявляется даже в условиях культивирования эпителиальных клеток [22, 23].
Динамика изменения толщины капсулы приведена в табл. 3.4.2. Эти сведения могут понадобиться хирургам, производящим экстракцию катаракты и использующим капсулу для крепления заднекамерных интраокулярных линз.
Капсула является довольно мощным барьером на пути бактерий и воспалительных клеток, но свободно проходима для молекул, размер которых соизмерим с размером гемоглобина [321, 798]. Хотя капсула не содержит элас-
208
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА
 Таблица 3.4.2. Динамика изменения толщины кап сулы хрусталика с возрастом, мкм (по Hogan, Alva- rado, Wedell, 1971)
Таблица 3.4.2. Динамика изменения толщины кап сулы хрусталика с возрастом, мкм (по Hogan, Alva- rado, Wedell, 1971)
| Зоны хрусталика | Возраст, лет | ||
| 2,5 | 35 | 71 | |
| Передний полюс Передний максимальный Экватор Задний максимальный Задний полюс | 8 12—15 7 18—22 2 | 14 21 17 23 4 | 14 21 9 9 23 |
тических волокон, она исключительно эластична и практически постоянно находится под действием внешних сил, т. е. в растянутом состоянии. По этой причине рассечение или разрыв капсулы сопровождается скручиванием. Свойство эластичности используется при проведении экстракапсулярной экстракции катаракты. Благодаря сокращению капсулы выводится содержимое хрусталика. Это же свойство используется также при лазерной капсулотомии.
В световом микроскопе капсула выглядит прозрачной, гомогенной (рис. 3.4.8). В поляризованном свете выявляется ее пластинчатая волокнистая структура. При этом волокнистость располагается параллельно поверхности хрусталика [203, 420]. Капсула также положительно окрашивается при проведении ШИК-реакции, что свидетельствует о наличии в ее составе большого количества протеогликанов [798].

Рис. 3.4.8. Светооптическое строение капсулы хрусталика, эпителия капсулы хрусталика и хрусталиковых волокон наружных слоев:
/ — капсула хрусталика; 2 — эпителиальный слой капсулы хрусталика; 3 — хрусталиковые волокна
Ультраструктурно капсула имеет относительно аморфное строение (рис. 3.4.6, 3.4.9). Незначительная пластинчатость намечается благодаря рассеиванию электронов нитевидными элементами, складывающимися в пластины.
Выявляется около 40 пластин, толщина каждой из которых равна приблизительно 40 нм [1197]. При большем увеличении микроскопа выявляются нежные коллагеновые фибриллы диаметром 2,5 нм [798].
В постнатальном периоде происходит некоторое утолщение задней капсулы, что свиде-
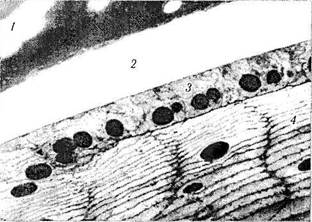
Рис. 3.4.9. Ультраструктура цинновой связки, капсулы хрусталика, эпителия капсулы хрусталика и хрусталиковых волокон наружных слоев:
/ — циннова связка; 2 — капсула хрусталика; 3 — эпителиальный слой капсулы хрусталика; 4 — хрусталиковые волокна
тельствует о возможности секреции базального материала задними кортикальными волокнами [798].
Fisher [320] установил, что 90% утраты эластичности хрусталика наступает в результате изменения эластичности капсулы.
В экваториальной зоне передней капсулы хрусталика с возрастом появляются электрон-ноплотные включения, состоящие из коллагено-вых волокон диаметром 15 нм и с периодом поперечной исчерченности, равной 50—60 нм. Предполагается, что они образуются в результате синтетической деятельности эпителиальных клеток [993]. С возрастом появляются и волокна коллагена, периодичность исчерченности которых равна 110 нм.
Места прикрепления цинновой связки к капсуле названы пластинами Бергера (Berger, 1882) (другое название—перикапсулярная мембрана). Это поверхностно расположенный слой капсулы, имеющий толщину от 0,6 до 0,9 мкм. Он менее плотный и содержит больше гликозаминогликанов, чем остальная часть капсулы. Волокна этого фиброгранулярного слоя перикапсулярной мембраны имеют толщину только 1—3 нм, в то время как толщина фибрилл цинновой связки 10 нм.
В перикапсулярной мембране обнаруживается фибронектин, витреонектин и другие матричные белки, которые играют определенную роль в прикреплении связок к капсуле [381, 435, 522, 657, 798]. В последнее время установлено наличие еще одного микрофиблиллярного материала, а именно фибриллина [743], о роли которого указано выше (см. 1-ю главу).
Подобно другим базальным мембранам капсула хрусталика богата коллагеном IV типа [790]. Она также содержит коллагены I, III и V типов. Обнаруживается и множество других внеклеточных матричных компонентов — лами-
Хрусталик и ресничный поясок (зонулярный аппарат)
209
 нин, фибронектин, гепаран сульфат и энтактин [588, 696].
нин, фибронектин, гепаран сульфат и энтактин [588, 696].
Проницаемость капсулы хрусталика человека изучалась многими исследователями [321, 338, 346, 481, 490]. Капсула свободно пропускает воду, ионы и другие молекулы небольшого размера. Она является барьером на пути белковых молекул, имеющих размер гемоглобина. Различий в пропускной способности капсулы в норме и при катаракте не обнаружил никто [346, 1104].
Эпителий хрусталика (epithelium lentis) состоит из одного слоя клеток, лежащих под передней капсулой хрусталика и распространяющихся на экватор (рис. 3.4.4, 3.4.5, 3.4.8, 3.4.9). Клетки на поперечных срезах кубовидной формы, а в плоскостных препаратах полигональные. Количество их колеблется от 350 000 до 1000 000 [556, 622, 798, 1215]. Плотность эпи-телиоцитов в центральной зоне — 5009 клеток в мм2 у мужчин и 5781—у женщин [428]. Плотность клеток несколько увеличивается по периферии хрусталика.
Необходимо подчеркнуть, что в тканях хрусталика, в частности в эпителии, преобладает анаэробный тип дыхания. Аэробное окисление (цикл Кребса) наблюдается только в эпителиальных клетках и наружных хрусталиковых волокнах, при этом этот путь окисления обеспечивает до 20% потребности хрусталика в энергии [798, 1126]. Эта энергия используется для обеспечения активных транспортных и синтетических процессов, необходимых для роста хрусталика, синтеза мембран, кристаллинов, белков цитоскелета и нуклеопротеинов. Функционирует и пентозофосфатный шунт, обеспечивающий хрусталик пентозами, необходимыми для синтеза нуклеопротеидов.
Эпителий хрусталика и поверхностные волокна коры хрусталика участвуют в выведении натрия из хрусталика, благодаря деятельности Na+—К+-насоса. При этом используется энергия АТФ. В задней части хрусталика ионы натрия во влагу задней камеры распространяются пассивно. Эпителий хрусталика состоит из нескольких субпопуляций клеток, отличающихся, в первую очередь, пролиферативной активностью [444, 510, 798, 926]. Выявляются определенные топографические особенности распределения эпителиоцитов различных субпопуляций. В зависимости от особенностей строения, функции и пролиферативной активности клеток выделяют несколько зон эпителиальной выстилки.
Центральная зона. Центральная зона состоит из относительно постоянного количества клеток, число которых медленно уменьшается с возрастом [22, 23, 798]. Эпителиоциты полигональной формы (рис. 3.4.9, 3.4.10, а), ширина их — 11 —17 мкм, а высота — 5—8 мкм. Своей апикальной поверхностью они прилежат к наиболее поверхностно расположенным хрустали-ковым волокнам. Ядра смещены к апикальной

Рис. 3.4.10. Ультраструктурная организация эпителиальных клеток капсулы хрусталика промежуточной зоны (а) и экваториальной области (б) (по Hogan et al., 1971):
1 — капсула хрусталика; 2 — апикальная поверхность соседней эпителиальной клетки; 3 —пальцевые вдавления в цитоплазму эпителиальной клетки соседних клеток; 4 — эпителиальная клетка, ориентированная параллельно капсуле; 5 — ядросодер-жащая эпителиальная клетка, расположенная в коре хрусталика
поверхности клеток большого размера и имеют многочисленные ядерные поры. В них, как правило, два ядрышка.
Цитоплазма эпителиоцитов содержит умеренное количество рибосом, полисом, гладкий и шероховатый эндоплазматический ретикулум, маленькие митохондрии, лизосомы и гранулы гликогена. Выражен аппарат Гольджи. Видны цилиндрической формы микротрубочки диаметром 24 нм, микрофиламенты промежуточного типа (10 нм), филаменты альфа-актинина [798].
При помощи методов иммуноморфологии в цитоплазме эпителиоцитов доказано наличие так называемых матричных белков — актина, винметина, спектрина и миозина, которые обеспечивают жесткость цитоплазмы клетки [52, 54, 86, 107, 798, 867].
В эпителии присутствует также альфа-крис-таллин. Бета- и гамма-кристаллины отсутствуют.
К капсуле хрусталика эпителиоциты присоединены при помощи полудесмосом [839]. Между эпителиоцитами видны десмосомы и щелевые контакты, имеющие типичное строение (см. главу 1) [858]. Система межклеточных контактов обеспечивает не только сцепление между эпителиальными клетками хрусталика, но определяет ионную и метаболическую связь между клетками.
Несмотря на наличие многочисленных межклеточных контактов между эпителиальными клетками, существуют пространства, выполен-ные бесструктурым материалом низкой электронной плотности. Ширина этих пространств колеблется от 2 до 20 нм. Именно благодаря этим пространствам осуществляется обмен метаболитов между хрусталиком и внутриглазной жидкостью.
210
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

 Эпителиальные клетки центральной зоны отличаются исключительно низкой митотичес-кой активностью [22, 23]. Митотический индекс равен всего 0,0004% и приближается к мито-тическому индексу эпителиоцитов экваториальной зоны при возрастной катаракте [556; 1187]. Существенно митотическая активность возрастает при различных патологических состояниях и, в первую очередь, после травмы [11, 444, 445, 859—862, 881, 926—928, 1157, 1158, 1188, 1189]. Увеличивается число митозов после воздействия на эпителиальные клетки ряда гормонов [929; 1192; 1124], при экспериментальных увеитах [760, 881, 1,157, 1188].
Эпителиальные клетки центральной зоны отличаются исключительно низкой митотичес-кой активностью [22, 23]. Митотический индекс равен всего 0,0004% и приближается к мито-тическому индексу эпителиоцитов экваториальной зоны при возрастной катаракте [556; 1187]. Существенно митотическая активность возрастает при различных патологических состояниях и, в первую очередь, после травмы [11, 444, 445, 859—862, 881, 926—928, 1157, 1158, 1188, 1189]. Увеличивается число митозов после воздействия на эпителиальные клетки ряда гормонов [929; 1192; 1124], при экспериментальных увеитах [760, 881, 1,157, 1188].
Промежуточная зона. Промежуточная зона находится ближе к периферии хрусталика. Клетки этой зоны цилиндрические с центрально расположенным ядром. Базальная мембрана имеет складчатый вид.
Герминативная зона. Герминативная зона прилежит к преэкваториальной зоне. Именно эта зона отличается высокой пролиферативной активностью клеток (66 митозов на 100 000 клеток), которая постепенно снижается с возрастом. Длительность протекания митоза у различных животных колеблется от 30 минут до 1 часа. При этом выявлены суточные колебания митотической активности [22, 23].
Клетки этой зоны после деления смещаются кзади и в последующем превращаются в хрус-таликовые волокна. Некоторые из них смещаются и кпереди, в промежуточную зону.
Цитоплазма эпителиоцитов содержит малочисленные органоиды [201]. Имеются короткие профили шероховатого эндоплазматическо-го ретикулума, рибосомы, маленькие митохондрии и аппарат Гольджи [13] (рис. 3.4.10, б). Количество органоидов нарастает в экваториальной области по мере увеличения количества структурных элементов цитоскелета [863] актина, виментина, белка микротрубочек, спект-рина, альфа-актинина и миозина. Существует возможность различить целые актиновые сете-подобные структуры, особенно видимые в апикальной и базальной частях клеток [865, 866, 1209]. Помимо актина в цитоплазме эпителиальных клеток выявлены виментин и тубулин [531]. Предполагают, что сократительные мик-рофиламенты цитоплазмы эпителиальных клеток способствуют путем их сокращения перемещению межклеточной жидкости.
В последние годы показано, что пролифера-тивная активность эпителиальных клеток герминативной зоны регулируется многочисленными биологически активными веществами — цитокинами [789]. Выявлено значение интерлей-кина-1, фактора роста фибробластов, трансформирующего фактора роста бета, эпидермаль-ного фактора роста, инсулиноподобного фактора роста, фактора роста гепатоцитов, фактора роста кератиноцитов, постагландина Е2. Часть перечисленных факторов роста стимулируют
пролиферативную активность, а часть — инги-бируют ее [73, 516, 789, 1161]. Необходимо отметить, что перечисленные факторы роста синтезируются или структурами глазного яблока, или другими тканями оранизма, поступая в глаз через кровь.
Процесс формирования хрусталиковых во локон. После конечного разделения клетки одна или обе дочерние клетки смещаются в смежную переходную зону, в которой клетки организованы в меридианально ориентированные ряды (рис. 3.4.4, 3.4.5, 3.4.11).


Рис. 3.4.11. Особенности расположения хрусталиковых волокон:
а — схематическое изображение; б — сканирующая электронная микроскопия (по Kuszak, I989)
В последующем эти клетки дифференцируются во вторичные волокна хрусталика, разворачиваясь на 180° и удлиняясь. Новые волокна хрусталика сохраняют полярность таким образом, что задняя (базальная) часть волокна сохраняет контакт с капсулой (базальной пластинкой), в то время как передняя (апикальная) часть отделена от этого эпителием. По мере превращения эпителиоцитов в хрусталиковые волокна фомируется ядерная дуга (при микроскопическом исследовании ряд ядер эпителиальных клеток, расположенных в виде дуги).
Хрусталик и ресничный поясок (зонулярньш аппарат)
211
 Предмитотическому состоянию эпителиальных клеток предшествует синтез ДНК, в то время как дифференциация клеток в хрустали-ковые волокна сопровождается усилением синтеза РНК, поскольку в этой стадии отмечается синтез структурных и мембранных специфических белков. Ядрышки дифференцирующихся клеток резко увеличиваются [629], а цитоплазма становится более базофильной в связи с увеличением количества рибосом [280, 555], что объясняется усилением синтеза мембранных компонентов [106], белков цитоскелета и кристаллинов хрусталика [372; 555]. Эти структурные изменения отражают усиление белкового синтеза [815].
Предмитотическому состоянию эпителиальных клеток предшествует синтез ДНК, в то время как дифференциация клеток в хрустали-ковые волокна сопровождается усилением синтеза РНК, поскольку в этой стадии отмечается синтез структурных и мембранных специфических белков. Ядрышки дифференцирующихся клеток резко увеличиваются [629], а цитоплазма становится более базофильной в связи с увеличением количества рибосом [280, 555], что объясняется усилением синтеза мембранных компонентов [106], белков цитоскелета и кристаллинов хрусталика [372; 555]. Эти структурные изменения отражают усиление белкового синтеза [815].
В процессе образования хрусталикового волокна в цитоплазме клеток появляются многочисленные микротрубочки диаметром 5 нм [686, 863] и промежуточные фибриллы [686, 687], ориентированные вдоль клетки и играющие важную роль в морфогенезе хрусталико-вых волокон [759, 798, 827].
Клетки различной степени дифференциации в области ядерной дуги располагаются как бы в шахматном порядке. Благодаря этому между ними образуются каналы, обеспечивающие строгую ориентацию в пространстве вновь дифференцирующихся клеток. Именно в эти каналы проникают цитоплазматические отростки. При этом образуются меридианальные ряды хрусталиковых волокон.
Важно подчеркнуть, что нарушение мериди-анальной ориентации волокон является одной из причин развития катаракты как у экспериментальных животных [1188, 1190, 1191], так и у человека [1050, 1104].
Превращение эпителиоцитов в хрусталико-вые волокна происходит довольно быстро. Это было показано в эксперименте на животных с использованием тимидина, меченного изотопом [148, 439, 732, 736, 1189]. У крыс эпителиоцит превращается в хрусталиковое волокно спустя 5 недель.
В процессе дифференциации и смещения клеток к центру хрусталика в цитоплазме хрусталиковых волокон уменьшается количество органоидов и включений. Цитоплазма приобретает гомогенный вид. Ядра подвергаются пик-нозу, а затем и полностью исчезают [550, 631, 1141]. Вскоре исчезают органоиды [96, 97, 550, 749, 750, 798, 815]. Basnett [96, 97] выявил, что потеря ядер и митохондрий наступает внезапно и в одном поколении клеток.
Количество хрусталиковых волокон на протяжении жизни постоянно увеличивается. «Старые» волокна смещаются к центру. В результате этого формируется плотное ядро.
С возрастом уменьшается интенсивность образования хрусталиковых волокон. Так, у молодых крыс в сутки формируется приблизительно пять новых волокон, в то время как у старых крыс —одно [148, 439, 736].
Особенности мембран эпителиальных кле ток. Цитоплазматические мембраны соседних эпителиальных клеток формируют своеобразный комплекс межклеточных связей. Если боковые поверхности клеток слегка волнистые, то апикальные зоны мембран образуют «пальцевые вдавления», погружающиеся в надлежащие хрусталиковые волокна. Базальная часть клеток присоединена к передней капсуле при помощи полудесмосом, а боковые поверхности клеток соединяются десмосомами.
На боковых поверхностях мембран смежных клеток обнаружены также щелевые контакты, через которые может происходить обмен небольшими молекулами между хрусталиковыми волокнами [96, 97, 629, 858]. В области щелевых контактов обнаруживаются белки кенне-сины различной молекулярной массы [1071]. Некоторые исследователи предполагают, что щелевые контакты между хрусталиковыми волокнами отличаются от таковых в других органах и тканях.
Исключительно редко можно увидеть плотные контакты [620, 664, 666].
Структурная организация мембран хрусталиковых волокон и характер межклеточных контактов свидетельствуют о возможном наличии на поверхности клеток рецепторов, контролирующих процессы эндоцитоза, который имеет большое значение в перемещении метаболитов между этими клетками [156]. Предполагается существование рецепторов к инсулину, гормону роста и бета-адренергическим антагонистам. На апикальной поверхности эпителиальных клеток выявлены ортогональные частицы, встроенные в мембрану и имеющие диаметр 6—7 нм [251, 452, 612, 635, 1029]. Предполагают, что эти образования обеспечивают перемещение между клетками питательных веществ и метаболитов [156, 623].
Волокна хрусталика (fibrae lentis) (рис. 3.4.5, 3.4.10—3.4.12). Переход от эпителиальных клеток герминативной зоны к хруста-ликовому волокну сопровождается исчезновением между клетками «пальцевых вдавлений», а также началом удлинения базальной и апикальной частей клетки. Постепенное накопление хрусталиковых волокон и смещение их к центру хрусталика сопровождается формированием ядра хрусталика. Это смещение клеток приводит к образованию S- или С-подобной дуги (ядерная дуга), направленной вперед и состоящей из «цепи» ядер клеток. В области экватора зона ядерных клеток имеет ширину порядка 300—500 мкм [629].
Расположенные глубже волокна хрусталика имеют толщину 150 мкм. Когда они теряют ядра, ядерная дуга исчезает. Хрусталиковые волокна имеют веретенообразную или ремнепо-добную форму, располагаясь по дуге в виде концентрических слоев. На поперечном разрезе в области экватора они гексагональной формы.
212
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

|

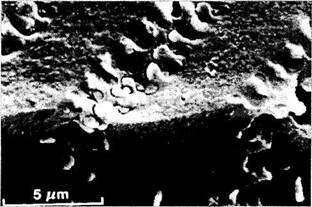
Рис. 3.4.12. Характер расположения хрусталиковых
волокон. Сканирующая электронная микроскопия
(по Kuszak, 1989):
а —плотно упакованные хрусталиковые волокна; б — «пальцевые вдавления»
По мере погружения к центру хрусталика постепенно нарушается их однообразие по размеру и форме. В области экватора у взрослых ширина хрусталикового волокна колеблется от 10 до 12 мкм, а толщина — от 1,5 до 2,0 мкм. В задних частях хрусталика волокна более тонкие, что объясняется асимметричной формой хрусталика и большей толщиной передней коры [621, 624]. Длина хрусталиковых волокон в зависимости от глубины расположения колеблется от 7 до 12 мм [183]. И это при том, что первоначальная высота эпителиальной клетки равняется всего 10 мкм.
Концы хрусталиковых волокон встречаются в определенном месте и формируют швы.
Швы хрусталика (рис. 3.4.13). В феталь-ном ядре имеется передний вертикально расположенный Y-образный и задний инвертированный Y-образный швы. После рождения по мере роста хрусталика и увеличения количества слоев хрусталиковых волокон, формирующих свои швы, происходит пространственное объединение швов с образованием звездоподоб-ной структуры, обнаруживающейся у взрослых.
Рис. 3.4.13. Формирование швов в месте стыка волокон, происходящее в различные периоды жизни:
/ — Y-образный шов, формирующийся в эмбриональном периоде; 2 — более развитая система швов, возникающая в детском периоде; 3 — наиболее развитая система швов, обнаруживаемая у взрослых
Основное значение швов заключается в том, что благодаря такой сложной системе контакта между клетками сохраняется форма хрусталика практически на протяжении всей жизни.
Особенности мембран хрусталиковых во локон. Контакты типа «пуговица — петля» (рис. 3.4.12). Мембраны соседствующих хрусталиковых волокон соединены при помощи разнообразных специализированных образований, изменяющих свое строение по мере смещения волокна с поверхности в глубь хрусталика. В поверхностных 8—10 слоях передних отделов коры волокна соединяются при помощи образований типа «пуговица — петля» («шар и гнездо» американских авторов), распределенных равномерно по всей длине волокна. Подобного типа контакты существуют только между клетками одного слоя, т. е. клетками одного поколения, и отсутствуют между клетками разных поколений. Это обеспечивает возможность передвижения волокон относительно друг друга в процессе их роста.
Между более глубоко расположенными волокнами контакт типа «пуговица — петля» обнаруживается несколько реже. Распределены они в волокнах неравномерно и случайным образом. Появляются они и между клетками различных поколений.
В самых глубоких слоях коры и ядра, кроме указанных контактов («пуговица — петля»), появляются сложные интердигитации в виде гребней, впадин и борозд [629, 798, 1170]. Обнаружены также и десмосомы, но только между дифференцирующимися, а не зрелыми хруста-ликовыми волокнами.
Предполагают, что контакты между хрус-таликовыми волокнами необходимы для поддержания жесткости структуры на протяжении всей жизни, способствующей сохранению про-
Хрусталик и ресничный поясок (зонулярный аппарат)
213
 зрачности хрусталика. Еще один тип межклеточных контактов обнаружен в хрусталике человека. Это щелевой контакт [825]. Щелевые контакты выполняют две роли. Во-первых, поскольку они соединяют хрусталиковые волокна на большом протяжении, сохраняется архитектоника ткани, тем самым обеспечивается прозрачность хрусталика [625]. Во-вторых, именно благодаря наличию этих контактов происходит распространение питательных веществ между хрусталиковыми волокнами. Это особо важно для нормального функционирования структур на фоне пониженной метаболической активности клеток (недостаточное количество органоидов).
зрачности хрусталика. Еще один тип межклеточных контактов обнаружен в хрусталике человека. Это щелевой контакт [825]. Щелевые контакты выполняют две роли. Во-первых, поскольку они соединяют хрусталиковые волокна на большом протяжении, сохраняется архитектоника ткани, тем самым обеспечивается прозрачность хрусталика [625]. Во-вторых, именно благодаря наличию этих контактов происходит распространение питательных веществ между хрусталиковыми волокнами. Это особо важно для нормального функционирования структур на фоне пониженной метаболической активности клеток (недостаточное количество органоидов).
Выявлено два типа щелевых контактов — кристаллические (обладающих высоким омическим сопротивлением) и некристаллические (с низким омическим сопротивлением). В некоторых тканях (печень) указанные типы щелевид-ных контактов могут преобразовываться один в другой при изменении ионного состава окружающей среды. В волокне хрусталика они неспособны к подобному преобразованию [392] Первый тип щелевых контактов найден в местах прилегания волокон к эпителиальным клеткам, а второй — только между волокнами [106, 627].
Низкоомные щелевые контакты содержат внутримембранные частицы, не позволяющие соседним мембранам сближаться более чем на 2 нм. Благодаря этому в глубоких слоях хрусталика ионы и молекулы небольшого размера достаточно легко распространяются между хрусталиковыми волокнами, и их концентрация довольно быстро выравнивается. Имеются и видовые различия в количестве щелевых контактов. Так, в хрусталике человека они занимают поверхность волокна по площади 5%, у лягушки— 15%, у крысы — 30%, а у цыпленка — 60% [625, 626, 665]. Щелевых контактов нет в области швов.
Необходимо кратко остановиться на факторах, обеспечивающих прозрачность и высокую рефракционную способность хрусталика. Высокая рефракционная способность хрусталика достигается высокой концентрацией белковых филаментов, а прозрачность — их строгой пространственной организацией, однородностью структуры волокон в пределах каждого поколения и небольшим объемом межклеточного пространства (менее 1% объема хрусталика). Способствует прозрачности и небольшое количество внутрицитоплазматических органоидов, а также отсутствие в хрусталиковых волокнах ядер. Все перечисленные факторы сводят к минимуму рассеивание света между волокнами.
Есть другие факторы, влияющие на рефракционную способность. Одним из них является увеличение концентрации белка по мере приближения к ядру хрусталика. Именно благодаря увеличению концентрации белка отсутствует хроматическая аберрация.
Не меньшее значение в структурной целостности и прозрачности хрусталика имеет и регуляция ионного содержания и степени гидратации волокон хрусталика. При рождении хрусталик прозрачен. По мере роста хрусталика появляется желтизна ядра. Возникновение желтизны, вероятно, связанно с влиянием на него ультрафиолетового света (длина волны 315—400 нм). При этом в коре появляются флюоресцирующие пигменты. Предполагают, что эти пигменты экранируют сетчатку от разрушительного действия коротковолновой световой радиации [1011]. Пигменты накапливаются в ядре с возрастом, а у некоторых людей участвуют в образовании пигментной катаракты. В ядре хрусталика в старческом возрасте и особенно при ядерной катаракте увеличивается количество нерастворимых белков, которые представляют собой кристаллины, молекулы которых «сшиты».
Метаболическая активность в центральных участках хрусталика незначительна. Практически отсутствует обмен белков [446]. Именно поэтому они относятся к долгоживущим белкам и легко подвергаются повреждению окислителями, приводящими к изменению конформации белковой молекулы из-за образования сульф-гидрильных групп между молекулами белка. Развитие катаракты характеризуется увеличением зон рассеивания света. Это может быть вызвано нарушением регулярности расположения хрусталиковых волокон, изменением структуры мембран и нарастанием рассеивания света, в связи с изменением вторичной и третичной структуры белковых молекул. Отек хрусталиковых волокон и их разрушение приводит к нарушению водно-солевого обмена.
Ресничный поясок
Ресничный поясок (зонулярный аппарат; связка Цинна; подвешивающая связка хрусталика; zonula ciliaris) состоит из волокон, распространяющихся от ресничного тела к экватору хрусталика (рис. 3.4.2). Они достаточно жестко фиксируют хрусталик в определенном положении и позволяют ресничной мышце выполнять свою основную функцию, а именно путем сокращений приводить к деформации хрусталика. При этом, естественно, изменяется его рефракционная способность. Связка Цинна образует кольцо, имеющее вид треугольника на меридианальном срезе. Основание этого треугольника вогнуто и противостоит экватору хрусталика. Верхушка этого треугольника направляется к отросткам ресничного тела, его плоской части и зубчатой линии.
Волокна ресничного пояска (fibrae zonu- lares) состоят из гликопротеида неколлагено-вого происхождения, связанного при помощи О- и N-связей с олигосахаридами. Наличие этих связей объясняет их положительное гистохими-
214
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

 ческое окрашивание при проведении ШИК-ре-акции [798].
ческое окрашивание при проведении ШИК-ре-акции [798].
Волокна зонулярного аппарата имеют строение трубочки диаметром 10 нм (8—12 нм) и напоминают эластические волокна как своим химическим составом, так и отношением к про-теолитическим ферментам (устойчивость к кол-лагеназе и трипсину) [798, 874; 925; 1051 — 1053]. Эту особенность используют при интра-капсулярной экстракции катаракты, применяя альфа-химотрипсин, лизирующий зонулярный аппарат, но не действующий на капсулу хрусталика. В тех случаях, когда волокна складываются в пучок, появляется периодичность в 40—55 мкм. Между волокнами обнаруживается мелкозернистый и волокнистый материал [798, 874, 1047].
Недавно показано, что волокна зонулярного аппарата богаты цистеином и аналогичны микрофибриллярному компоненту эластической ткани. Эти микрофибриллы называются фиб-риллином и окрашиваются соответствующими моноклональными антителами [956, 957, 1047, 1162, 1195].
В других тканях фибриллин является матрицей для образования эластических волокон [240, 924, 925], обеспечивая эластические свойства многих структур. Аналогичную функцию они имеют и в зонулярном аппарате.
Ген, контролирующий синтез фибриллина, располагается в хромосоме 15q21. 1 [649, 685]. Синдром Марфана, при котором выявляются дислокация хрусталика и различные заболевания сердечно-сосудистой системы, связан с мутаций именно этого гена, контролирующего синтез фибриллина [254, 551, 649]. При этом строение микрофибрилл изменяется. Количество волокон зонулярного аппарата уменьшается [303], волокна растянуты, а их диаметр различный [743, 820]. Обнаруживается также уменьшение их эластичности и разрушение [572, 820].
Близкие по характеру изменения фибриллина определяются и при других аномалиях глаза, сопровождающихся подвывихом хрусталика. К ним относятся осевая близорукость, пре-сенильная катаракта, открытоугольная глаукома [533], косоглазие [532], плоская роговица и гипоплазия ресничной мышцы и радужной оболочки, приводящие к миозу [220]. Определяется также удлинение ресничных отростков [482, 869]. Обнаруживается нарушение синтеза фибриллина при синдроме Марфана [1177], псевдо-эксфолиативном синдроме [342, 972, 973]. Нарушение строения фибриллина отмечено и при старении. Сопровождается этот процесс ослаблением зонулярного аппарата [442, 955].
Зонулярный аппарат исходит из наружного слоя капсулы хрусталика в экваториальной области. Причем на передней поверхности капсулы связка образует полосу прикрепления шириной 2,5 мм, а на задней поверхности — 1,0 мм.
При этом фибриллы, исходящие из переднего отдела экваториальной поверхности хрусталика направляются кзади и прикрепляются к ресничным отросткам («передние связки»), а фибриллы, отходящие от задней поверхности капсулы, направляются к плоской части ресничного тела и зубчатой линии («задние связки»). Экваториальные нити распространяются от ресничных отростков непосредственно к экватору. Выделяют и гиалоидные нити связки, которые распространяются от плоской части ресничного тела к краю хрусталика на участке его прилегания к стекловидному телу. Здесь они вплетаются в «гиалоидокапсулярную связку» [300, 791, 904].
В связи с тем, что нити связки, идущие от хрусталика, направляются к различным отделам ресничного тела, между ними образуются потенциальные пространства (пространства пояска; spatia zonularis), выполненные водянистой влагой. Это канал Гановера (Hanover) (между «передними» и «задними» нитями связи) и канал Петита (Petit) (между «задними связками» и передней поверхностью стекловидного тела).
Сканирующая электронная микроскопия способствовала большему пониманию особенностей строения и прикрепления цинновой связки к хрусталику. Подавляющее большинство волокон исходят из плоской части ресничного тела кпереди на расстоянии 1,5 мм от зубчатой линии. Здесь они переплетаются с внутренней пограничной мембраной эпителиальных клеток [904] или продолжаются в волокна переднего отдела стекловидного тела [290, 791] (рис. 3.4.14). Большинство волокон складывается в пучки, состоящие из 2—5 фибрилл. Некоторые фибриллы иногда проникают между эпителиальными клетками. Фибриллы обнаруживаются и между пигментированными эпителиальными клетками ресничного эпителия и вплетаются в их базальную мембрану и эластическую пластинку мембраны Бруха [721, 922].
«Передние волокна связки» распространяются до тех пор, пока не достигнут заднего края отростчатой части ресничного тела. Здесь они образуют «зонулярное сплетение», которое распространяется между ресничными отростками и прикрепляются к их боковым стенкам. Фибриллы «зонулярного сплетения» плотно присоединяются в основании ресничных гребешков, стабилизируя всю систему связок. Несколько кпереди отростчатой части ресничного тела «зонулярное сплетение» разделяется и состоит из трех пучков волокон, направляющихся к передней, экваториальной и задней капсуле хрусталика [904].
Характер преэкваториального, экваториального и заэкваториального прикрепления волокон зонулярной связки отличаются между собой (рис. 3.4.14). Преэкваториальные волокна связки относительно плотные. Они все прикрепляются на одном и том же расстоянии от
Хрусталик и ресничный поясок (зонулярный аппарат)
215

|

Рис. 3.4.14. Сканирующая электронная микроскопия экваториальной зоны хрусталика, иллюстрирующая особенности распространения цинновой связки между ресничным телом и хрусталиком и места ее прикрепления (по Bron et al., 1997):
I — экватор хрусталика; 2 — циннова связка; 3 — ресничные отростки
экватора (1,5 мм) в виде двойного ряда нитей связки шириной 5—10 мкм. Волокна связки при прикреплении суживаются и расплющиваются в плоскости капсулы хрусталика, формируя при этом «зонулярные пластинки» (пластины Бергера).
«Передние нити связки» в месте прикрепления отдают в капсулу тонкие фибриллы (от 0,07 до 0,5 мкм) на глубину 0,6—1,6 мкм. В результате этого «зонулярная пластинка» утолщается до 1,0—1,7 мкм.
Указывается на то, что число волокон «передних связок» уменьшается с возрастом. При этом вставки их смещаются к центру капсулы [1153]. Экваториальных волокон меньше. Они также как и «передние» и «задние» при прикреплении к капсуле щеткоподобно расщепляются. Фибриллы обычно шириной от 10 до 15 мкм, но могут достигать и 60 мкм.
«Задние волокна» прикрепляются двумя или тремя слоями в зоне шириной от 0,4 до 0,5 мм. Спереди они прикрепляются к заднему краю экватора хрусталика, а сзади простираются на расстояние 1,25 мм от края экватора. В месте прикрепления волокна цинновой связки погружаются в капсулу хрусталика примерно до 2 мкм.
«Постэкваториальные волокна», на первый взгляд кажутся менее развитыми, чем «передние». Это мнение ошибочно, поскольку они прикрепляются к капсуле на различных уровнях, включая вплетение в волокна передней поверхности стекловидного тела. «Стекловидные связки» являются отдельным слоем волокон, которые соединяют передний отдел стекловидного тела с плоской и отростчатой частями ресничного тела.
Streeten [1045] предполагает, что слизе-подобный характер цинновой связки является
барьером на пути распространения веществ между задней камерой глаза и стекловидным телом.
Возрастные изменения ресничного пояска (связки Цинна). В эмбриональном периоде нити связки Цинна нежные и слабо связаны между собой. Высока в них концентрация про-теогликанов. В пожилом возрасте количество волокон значительно уменьшается [165, 1153]. В первые два десятилетия жизни участки прикрепления цинновой связки в капсуле хрусталика довольно узкие. Со временем они расширяются и передвигаются к центру капсулы хрусталика, что связано с ростом хрусталика и увеличением его диаметра. При этом свободная от связки поверхность передней капсулы хрусталика уменьшается с 8 мм в возрасте 20 лет до 6,5 мм на восьмом десятилетии жизни [302, 1030]. Иногда она сужается до 5,5 мм, что существенно усложняет проведение капсулото-мии при проведении экстракапсулярной экстракции катаракты [302, 1030].
При интракапсулярной экстракции катаракты большая часть связочного комплекса отрывается от капсулы. Сохраняются только кончики передних зонулярных вставок и некоторое количество меридианальных волокон.
Циннова связка ослаблена при псевдоэксфолиации капсулы хрусталика, что может явиться причиной разрыва связок при хирургическом лечении катаракты [1010].
Роль ресничного пояска в аккомодации. Особенности функционирования аккомодирующей системы глаза до конца еще не совсем понятны. В этом процессе принимают участие многие структуры — ресничное тело, ресничный поясок, хрусталик, стекловидное тело. При этом конечный результат работы аккомодационной системы зависит от структурных и функ-
216
Глава 3. СТРОЕНИЕ ГЛАЗНОГО ЯБЛОКА

 циональных особенностей указанных структур, а также степени контроля этого процесса нервной и гуморальной системами. Более подробно о работе аккомодационной системы мы расскажем в разделе «Ресничное тело». Здесь же изложим основные принципы ее работы с уде-лением особого внимания роли в этом процессе цинновой связки.
циональных особенностей указанных структур, а также степени контроля этого процесса нервной и гуморальной системами. Более подробно о работе аккомодационной системы мы расскажем в разделе «Ресничное тело». Здесь же изложим основные принципы ее работы с уде-лением особого внимания роли в этом процессе цинновой связки.
Общепринято, что циннова связка при отсутствии сокращения ресничной мышцы натянута, что приводит к уплощению хрусталика в результате растяжения его. В процессе аккомодации сокращение ресничных мышц приводит к тому, что ресничные отростки смещаются кнут-ри. При этом циннова связка расслабляется, и хрусталик становится более сферичным благодаря его эластичности и способности к обратимой деформации. Периметр хрусталика при этом уменьшается и увеличивается относительный размер ядра хрусталика [158]. Передняя поверхность хрусталика становится более изогнутой и перемещается кпереди. Каких-либо существенных изменений кривизны задней поверхности не отмечается, что, видимо, связано с довольно высокой плотностью стекловидного тела [208].
Вышеприведенный механизм аккомодации, выдвинут еще Гельмгольцем [272] и подтвержден экспериментальными исследованиями с использованием киносъемки смещения цинновой связки и деформации хрусталика [772].
Отсутствие изменения кривизны задней поверхности хрусталика связывают с особым характером прикрепления цинновой связки к задней капсуле хрусталика. По мнению Rohen et al. [904, 911], циннова связка, направляющаяся к задней поверхности капсулы хрусталика, начинается от плоской части ресничного тела. Именно по этой причине сокращение ресничной мышцы не приводит к существенному смещению связки и, естественно, сила, прилагаемая к задней поверхности хрусталика, незначительная. Правда, ряд исследователей не поддерживают эту теорию [236, 300, 795].
Процессы, приводящие к расслаблению цинновой связки и связанные с координированным сокращением ресничной мышцы, приведены в разделе «Ресничное тело».

