




Исследование реакции биотоков мозга на воздействие ритмического светового раздражителя (реакции навязывания ритма) таит в себе большие возможности для получения результатов в дифференциально-психофизиологическом аспекте. Эта реакция наблюдается практически у всех испытуемых даже при визуальной оценке эффекта, хотя детальное ее изучение предпочтительнее вести с помощью частотных анализаторов и интеграторов, позволяющих регистрировать и количественно оценивать динамику отдельных участков спектра электроэнцефалограммы.
В реакции навязывания ритма наблюдается весьма значительный диапазон индивидуальных различий. Он одинаково велик как при изучении частотных характеристик реакций, так и при определении зависимости ее от интенсивности, цветности или иных параметров предъявляемого раздражителя (Н.Н. Данилова, 1963; В.Н. Ильянок, 1961; А.Г. Поваринский, 1960; G.A. Ulett, L. С. Johnson, 1958). В отдельных работах, главным образом зарубежных, эти индивидуальные вариации сопоставлялись с некоторыми особенностями личности (А.С. Mundy-Castle, 1953; G.A. Ulett et al., 1953; С. Shagass, 1955). В других работах изучалась зависимость реакции навязывания от различных состояний объекта, человека или животного (Л.Я. Голубева, 1959; Н.Н. Данилова, 1958, 1959; А.Г. Копылов, 1956, 1960; А. Крейндлер, 1960; Д.А. Фарбер, 1960; А.О. Долин, Д.А. Фарбер, 1963; Н.Н. Зислина, А.А. Новикова, 1962).
Первым опытом исследования реакции навязывания ритма в связи со свойствами нервной системы, наколько нам известно, явилась работа Э.А. Голубевой (1963), выполненная в лаборатории Б.М. Теплова и имевшая целью сопоставление некоторых показателей реакции навязывания ритма с характеристикой баланса нервных процессов (последняя была определена В.И. Рождественской по соотношению скоростей образования положительных и тормозных фотохимических условных реакций и, таким образом, представляет собой характеристику баланса по динамичности; кроме того, о ней судили по кожно-гальваническому показателю).
В этой работе было найдено, – правда, на малом числе испытуемых, – что индивиды с преобладанием возбуждения имеют определенную тенденцию к лучшему навязыванию низких частот стимуляции – от 1,5 до 7 имп/с. Этот факт был предположительно объяснен Э.А. Голубевой тем известным обстоятельством, что некоторые индивиды с преобладанием возбуждения имеют склонность быстро впадать в тормозное, сонное состояние, для которого характерно как раз более выраженное навязывание медленных частот.
Вслед за этой работой в лаборатории психофизиологии была выполнена целая группа исследований, посвященных определению роли свойств нервной системы в различных параметрах реакции навязывания ритма (В.Д. Небылицын, 1964 в, г; Э.А. Голубева, 1965; Э.А. Голубева, Л.А. Шварц, 1965). Одним из таких исследований был поставленный нами комплексный эксперимент, в котором подвергались сопоставлению показатели: 1) навязывания ритмов в диапазоне от 5 до 22 имп/с и при различных интенсивностях светового раздражителя, 2) ориентировочной и условнорефлекторной блокады альфа-ритма при воздействии звуковых и световых раздражителей и их сочетаний и 3) «фоновых» особенностей электроэнцефалограммы, точнее, ее составляющих, выделенных с помощью анализатора частот.
Остановимся на тех итогах количественной обработки указанных сопоставлений, которые имеют отношение к проблеме динамичности нервных процессов. Те аспекты реакции навязывания ритма, которые обнаружили связь с проблемой силы нервной системы, будут обсуждены в гл. 9.
Кратко опишем методическую сторону эксперимента.
Испытуемый находился в звукоизолированной камере в положении лежа с открытыми глазами в темноте. Регистрировалось биполярное височно-затылочное отведение справа; выбор отведения диктовался желанием получить максимальную выраженность альфа-ритма для более четкого определения его фоновых особенностей и реакций. Применение полосовых фильтров, выделявших из ЭЭГ частоты 4 – 7, 8–13 и 14 – 25 Гц (В.А. Кожевников, 1954), позволило регистрировать одновременно с электроэнцефалограммой на трех других перьях ее составляющие: тета-, альфа- и бета-активность; включение в схему интегратора (В.А. Кожевников, В.И. Сороко, 1959) давало возможность количественной оценки электрической активности (энергии колебаний) в полосах каждого из выделяемых ритмов. Источником световых импульсов в опытах с усвоением ритма служил фотостимулятор ЭФС‑01; длительность светового импульса оставалась постоянной, составляя 10 мс, частота и интенсивность варьировались соответственно конкретной программе опыта.
Методика получения показателей навязывания ритма несколько отличается у нас от той, которая используется большинством авторов. Мы стремились получить точные количественные индикаторы эффекта навязывания и применили с этой целью прием, позволяющий учесть «чистый» эффект навязывания ритма. Этот прием заключается в следующем. Прежде чем перейти к предъявлению мелькающего света, мы измеряли энергию колебаний данной частотной полосы при действии непрерывного света той же длительности (10 с) и тех же интенсивностей, что и ритмический свет. После этого мы приступали к измерению энергии в данной полосе при действии мелькающего света соответствующей частоты (в полосе тета-ритма при стимуляции частотой 5, 6 и 7 имп/с, в полосе альфа-ритма при стимуляции 9, 10, 11 и 12 имп/с и в полосе бета-ритма при стимуляции 16, 18, 20 и 22 имп/с). Индекс навязывания ритма выводился как разность между вторым и первым показателями и означает, таким образом, прирост суммарной энергии колебаний в данной частотной полосе при действии мелькающего света по сравнению с энергией, измеренной при действии непрерывного света; он означает, следовательно, «чистый» эффект воздействия ритмического светового раздражителя. Окончательный индекс для каждой из указанных частот вычислялся путем суммирования индексов для шести применявшихся интенсивностей стимуляции (25, 50, 100, 200, 500 и 1000 лк). Он обозначается нами как энергетический индекс навязывания ритма (ЭИНР). Кроме вычисления индексов для отдельных частот производился (путем простого» сложения) подсчет суммарных ЭИНР для частотных полос в целом, а также общего индекса, суммарного для всех применявшихся частот.
Индивидуальные индексы навязывания сопоставлялись с рядом ориентировочных и условно-ориентировочных показателей реакции активации; эти показатели, полученные у тех испытуемых, которые обладали достаточно выраженным альфа-ритмом (20 из 25 человек), перечислены в табл. 6. Что касается показателей фона, то к ним относятся частота, суммарная энергия и индекс тета-, альфа- и бета-ритма. Частота и индекс ритмов подсчитывались в одном из опытов ближе к концу экспериментальной серии; суммарная энергия каждой из частотных полос измерялась в каждом из трех опытов по навязыванию ритма, и затем выводились средние.
Обзор полученных данных мы начнем с изложения результатов сопоставления ЭИНР для различных частот стимуляции.
Индивидуальные индексы навязывания для каждой из частот, а также для трех ритмов в целом и суммарные ЭИНР для всех частот приведены в табл. 9. В этой таблице встречаются отрицательные значения. Они обязаны своим происхождением тому, что в некоторых случаях значения энергии колебаний при действии непрерывного света превышают таковые, получаемые при действии мелькающего света; мелькающий свет даже тех частот, которые соответствуют данному ритму, приводит к большему его угнетению, чем непрерывный. Имеет место как бы «отрицательное навязывание», наблюдаемое, впрочем, довольно редко и при сильных интенсивностях света реже, чем при слабых.
Что касается сравнительного эффекта стимуляции, соответствующей разным ритмам, то, как можно видеть из таблицы, наибольшие абсолютные значения принадлежат, как правило, частотам альфа-полосы, во многих случаях – частотам бета-полосы и у отдельных испытуемых – частотам, соответствующим тета-ритму. Наибольший средний эффект наблюдается на частоте 12 имп/с, затем на частоте 11 имп/с, наименьший эффект дает частота 5 имп/с. Для большей наглядности эти отношения на рис. 17 представлены в виде гистограммы.

Рис. 17. Навязывание ритма как функция частоты ритмической световой стимуляции (средние данные). Ось абсцисс – частота стимуляции (имп/с); ось ординат – «чистый» эффект назязывания (усл. ед.).
Таким образом, у подавляющего большинства испытуемых навязывание ритма имеет место на всех примененных частотах стимуляции, в том числе и частотах, соответствующих тета-ритму, в чем наши данные согласуются с результатами Э.А. Голубевой (1963) и некоторых других авторов.
В таблице интеркорреляций (табл. 10) даны значения коэффициентов корреляции ЭИНР для каждой частоты с ЭИНР для каждой другой частоты. Рассмотрение этой таблицы приводит к любопытным выводам. Один из них заключается в том, что показатели навязывания для самых низких частот – 5, 6 и 7 имп/ с – на высоком уровне значимости (р < 0,01; р < 0,001) коррелируют между собой, но почти не коррелируют с показателями навязывания других частот, особенно высоких. В то же время, аналогичные показатели для частот, лежащих в зоне бета-ритма (16, 18, 20, 22 имп/с), дают чрезвычайно высокие взаимные корреляции (р < 0,001), но их корреляции с показателями всех других частот практически равны нулю. Что же касается ЭИНР частот, входящих в полосу альфа-ритма (9, 10, И и 12 имп/с), то они не только не коррелируют с остальными показателями, но и между собой находятся в весьма своеобразных отношениях, именно коррелируют показатели навязывания только соседних частот: 9 и 10, 10 и 11, 11 и 12 имп/с, причем заметим, первая и третья пары коррелируют на уровне р < 0,001, а вторая – только на минимальном уровне значимости р < 0,05. При сопоставлении ЭИНР частот этой полосы «через одну» (9 и 11, 10 и 12 имп/с) или «через две» (9 и 12 имп/с) корреляции оказываются статистически незначимыми или вообще близкими к нулевым.
Таблица 9
Энергетические индексы навязывания ритмов (В.Д. Небылицын, 1964в)
| Номер испытуемого | Частоты, имп/с | Ритмы | Общий | ||||||||||||
| тета | альфа | бета | |||||||||||||
| –26 | –23 | ||||||||||||||
| –10 | –20 | –23 | |||||||||||||
| –13 | –1 | ||||||||||||||
| –2 | –13 | –17 | –32 | –19 | –81 | ||||||||||
| –1 | –19 | ||||||||||||||
| –24 | –15 | ||||||||||||||
| –1 | –3 | ||||||||||||||
| –1 | |||||||||||||||
| –2 | ' 415 | ||||||||||||||
| Средние | 11,2 | 20,9 | 37,6 | 26^ | 65,5 | 116,0 | 128,2 | 33,7 | 55,2 | 55,4 | 22,6 | 69,8 | 336,0 | 167,1 | 572,3 |
Примечание. Индексы навязывания даны в усл. ед., которые с помощью специальной формулы могут быть пересчитаны в мкВ/с.
На рис. 18–21 приводятся примеры того, как у разных испытуемых по-разному осуществляется реакция навязывания ритма на различных частотах стимуляции.
Все это означает, что навязывание частот группы тета определяется одним фактором работы нервной системы, навязывание частот группы бета – другим фактором, притом, судя по характеру корреляций, чрезвычайно резко ограниченным от остальных, а в альфа-полосе действуют, возможно, даже два фактора (хотя, как видно, и связанные между собой), один из которых определяет эффект навязывания ритмической стимуляции, соответствующей низкочастотному альфа-ритму, а другой вступает в действие при стимуляции, соответствующей более высокочастотному альфа-ритму.

Рис. 18. Мелькающий свет частотой 6 имп/с вызывает отчетливо выраженную реакцию навязывания ритма в тета- и отчасти в альфа-полосе (вторая гармоника).
Обозначения: 1 – отметка действия раздражителя, 2 – тета-ритм, 3 – альфа-ритм, 4 – бета-ритм, 5 – неанализиро-ванная ЭЭГ, 6 – отметка времени.

Рис. 19. Мелькающий свет частотой 10 имп/с приводит к появлению навязанного ритма в альфа-полосе. Обозначения те же, что на рис. 18.
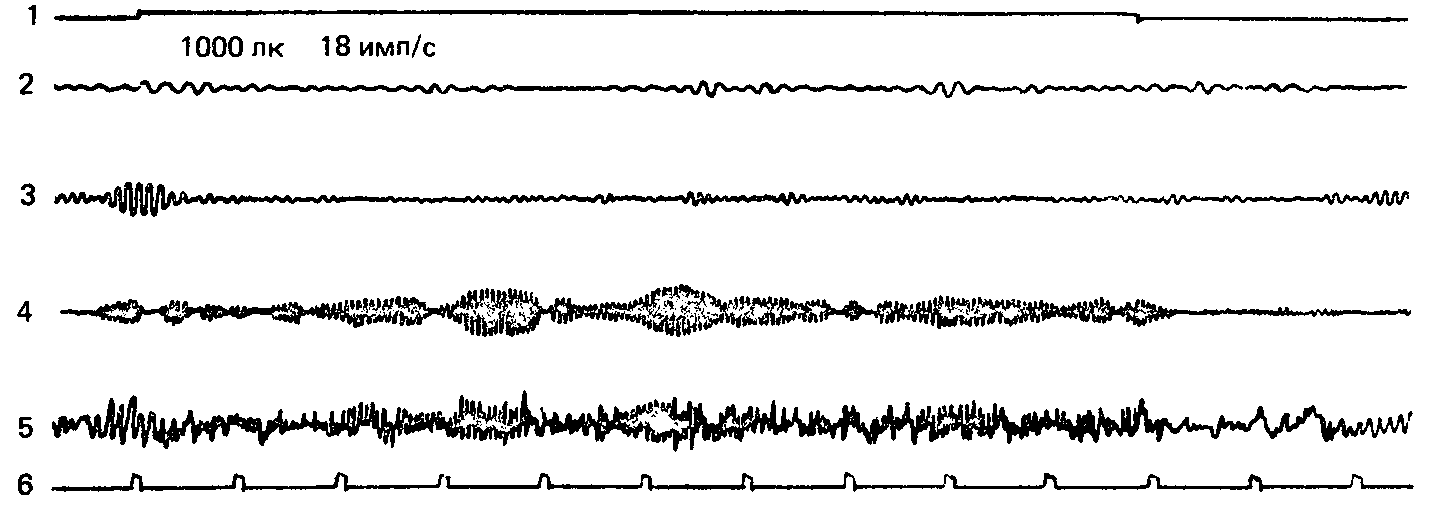
Рис. 20. Мелькающий свет частотой 18 имп/с вызывает ярко выраженную реакцию навязывания ритма в бета-полосе. Обозначения те же, что на рис. 18.
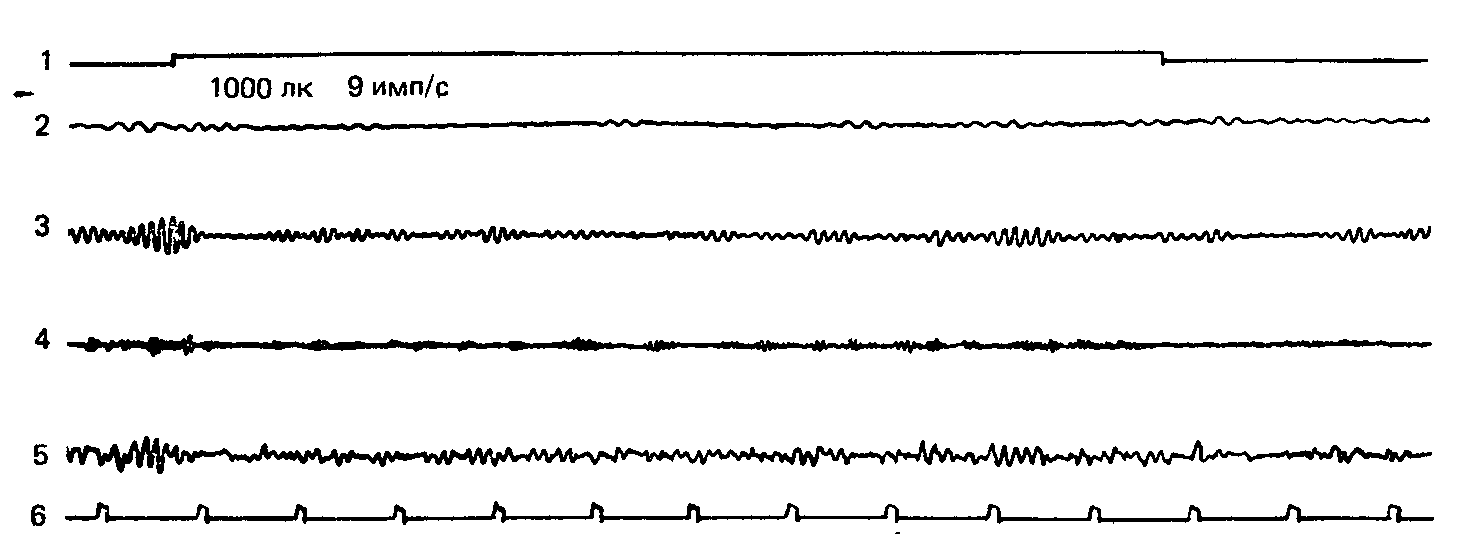
Рис. 21. Мелькающий свет частотой 9 имп/с не вызывает реакции навязывания. Обозначения те же, что на рис. 18.
Таблица 10
Интеркорреляции индексов навязывания ритма для различных частот (В.Д. Небылицын, 1964 в)
| Частоты, имп/с | Частоты, имп/с | ||||||||||
| 562** | 568*** | –020 | -212 | -086 | –0,76 | ||||||
| 784*** | –017 | –002 | |||||||||
| 475* | |||||||||||
| 686*** | –069 | -064 | -175 | ||||||||
| 429* | -216 | ||||||||||
| 788*** | |||||||||||
| -002 | -018 | ||||||||||
| 807*** | взо*** | 699*** | |||||||||
| 904*** | 728*** | ||||||||||
| 768*** | |||||||||||
Примечание. • р < 0,5; ** р < 0,01; ***р < 0,001.
Таким образом, можно думать, что нейрофизиологические механизмы формирования реакции навязывания различны при различных частотах световой стимуляции.
Возможно, однако, что эти очень четко разграниченные частотные группировки обязаны своим возникновением причинам более внешнего свойства, а именно тому, что сам ряд частот световой стимуляции, использованный в эксперименте, имел перерывы как раз в тех интервалах, которые лежат на границах физиологических ритмов: не применялись частоты 8, 13, 14 и 15 имп/с. Если бы эти частоты были использованы, то, может быть, и грани между ритмами, выявившиеся в эксперименте, не были бы столь отчетливыми. На эту мысль наводят результаты, полученные в исследовании реакции навязывания ритма Э.А. Голубевой (1965), которая использовала гораздо более дробный и широкий логарифмический ряд частот от 2 до 80 имп/с.
Следует заметить, что методика работы Э.А. Голубевой довольно значительно отличалась от нашей: ею был использован узкополосный анализатор частот, позволявший выделить из ЭЭГ весьма узкий участок спектра, соответствующий частоте наносимого в данный момент раздражения, и счетчик импульсов, сосчитывавший число волн данной частоты с амплитудой выше 10 мкВ (1963). Одно из отличий этого способа от примененного нами заключается, таким образом, в том, что колебания с амплитудой менее 10 мкВ не учитывались вовсе, а с амплитудой более 10 мкВ рассматривались как одинаковые независимо от их действительной величины, в то время как применение индикатора позволяет учитывать энергию колебаний любой амплитуды. Другое отличие заключается в способе подсчета эффекта ритмической стимуляции: индексом навязывания, применяемым Э.А. Голубевой, служило число колебаний (указанной амплитуды за данный отрезок времени) как таковое, в то время как у нас индексом навязывания являлась разность между показателями интегратора при мелькающем свете и при непрерывной световой стимуляции.
Не приходится сомневаться, что разница в методике приводит и к определенным различиям в результатах.
Таблица 11
Интеркорреляции коэффициентов (Э.А. Голубева, 1965)
| Частота, имп/с | Частота, имп/с | |||||||||||||||||||||||||
| 2,5 | 3,5 | 4,5 | ||||||||||||||||||||||||
| 481** | 387* | –107 | –038 | –008 | –141 | -125 | ||||||||||||||||||||
| 2,5 | 397* | 486** | –082 | –047 | ||||||||||||||||||||||
| 456** | 370** | |||||||||||||||||||||||||
| 3,5 | 804*** | 636*** | 523** | 396* | 333* | 430** | 455** | 561** | 472** | 433** | 377* | 343* | ||||||||||||||
| 785*** | 694*** | 566*** | 536*** | 364* | 477** | 361* | 541*** | 692*** | 574*** | 509** | 473** | 337* | 346* | |||||||||||||
| 4,5 | 847*** | 705*** | 539*** | 555*** | 482** | 367* | 339* | 388* | 582*** | 727*** | 613*** | 566*** | 501** | 378* | ||||||||||||
| 760*** | 630*** | 570*** | 346* | 369* | 509** | 425** | 474** | 419* | ||||||||||||||||||
| 786*** | 077*** | 527*** | 452** | 351* | 383* | 532*** | 493** | 404* | 345* | –044 | –097 | –049 | ||||||||||||||
| 763*** | 616*** | 539*** | 470** | 477** | 389* | 466** | 436** | 443** | 495** | 332* | 350* | 359* | ||||||||||||||
| 555*** | 531*** | 422* | 399* | 393* | 356* | |||||||||||||||||||||
| 523** | 458** | 337* | 483** | 514** | 485** | 512** | 503** | 469** | 430** | 379* | ||||||||||||||||
| 750*** | 637*** | 459** | 486** | 476** | 336* | 332* | 361* | 366* | 340* | 404* | ||||||||||||||||
| 817*** | 640*** | 481** | 422* | 433** | 509** | 495** | 521** | 414* | 437** | 346* | ||||||||||||||||
| 726*** | 558*** | 551*** | 488** | 471** | 457** | 465** | 396* | 444** | ||||||||||||||||||
| 789*** | 696*** | 499** | 425** | 393* | ||||||||||||||||||||||
| 820*** | 588*** | 400* | –032 | –112 | –023 | |||||||||||||||||||||
| 653*** | 470** | 419* | –066 | |||||||||||||||||||||||
| 841*** | 686*** | 553*** | 425** | 542*** | 333* | |||||||||||||||||||||
| 865*** | 718*** | 658*** | 558*** | 532*** | 448** | 466** | ||||||||||||||||||||
| 807*** | 764*** | 574*** | 581*** | 605*** | 597*** | |||||||||||||||||||||
| 742*** | 653*** | 717*** | 691*** | 717*** | ||||||||||||||||||||||
| 696*** | 625*** | 693*** | 687*** | |||||||||||||||||||||||
| 806*** | 633 *** | 670 *** | ||||||||||||||||||||||||
| 836*** | 845*** | |||||||||||||||||||||||||
| 936*** | ||||||||||||||||||||||||||
Примечание. • р <* 0,5\ **р <* 0,01; ***р < 0,001.
Данные Э.А. Голубевой представлены в виде интеркорреляций в табл. 11. По распределению коэффициентов эта таблица в некоторых чертах напоминает матрицу, полученную нами (табл. 10), в других – довольно сильно отличается от последней. Определенное сходство можно усмотреть в наличии некоторых группировок коэффициентов согласно физиологическим ритмам. Так, индексы навязывания частот зоны дельта почти не коррелируют с индексами навязывания других частот, и то же можно сказать о навязывании наиболее высоких из примененных частот (более 50 имп/с), хотя и те и другие, в особенности последние, имеют высокие внутренние корреляции. Индексы зоны тета коррелируют между собой и с индексами соседних частот, относящихся к низкочастотному альфа-ритму, но слабо коррелируют с индексами частот, соответствующих более высокочастотному альфа-ритму (10 и 12 кол/с). Что касается различий в распределении корреляций, то главное из них заключается в том, что индексы зоны тета, а также (в меньшей степени) зоны альфа обнаруживают довольно высокие положительные корреляции с индексами навязывания частот, соответствующих низкочастотному бета-ритму (до 30 – 35 кол/с).
Нужно предположить, следовательно, что при данном методическом подходе к изучению реакции навязывания в основе проявления эффектов низкочастотной (3,5 – 7 имп/с) и средней частоты (16 – 35 имп/с) стимуляции лежат, по-видимому, общие мозговые механизмы, природа которых, впрочем, в настоящий момент отнюдь не представляется ясной. Возможно, что эта общность обусловлена теми же причинами, в силу которых и тета и бета-активность часто рассматриваются как имеющие отношение к эмоциональным проявлениям индивида.
Заканчивая анализ проблемы соотношения между эффектами навязывания ритма при различных частотах стимуляции, можно, как мы полагаем, независимо от методических различий сделать некоторые общие выводы. Вероятно, резких граней между частотными группировками, выявляющимися при изучении реакции навязывания, и соответствующими отдельными физиологическими ритмами не существует, так же как не существует резких граней между самими физиологическими ритмами; в этом убеждает хотя бы факт наличия весьма высоких коэффициентов корреляции между индексами навязывания всех рядом лежащих частот (табл. 11). Тем не менее сами эти частотные группировки, по-видимому, выявляются с достаточной отчетливостью, соответствуя, в общем, основным физиологическим ритмам.
Этот вывод, основанный на анализе данных статистической обработки, перекликается с заключениями, к которым приходят некоторые авторы в результате непосредственного физиологического эксперимента. Так, А.Я. Супин (1963), анализируя механизмы формирования потенциалов при ритмическом световом воздействии, отмечает, что ответы коры на частые и редкие вспышки «не вполне идентичны», так как возникают на различном уровне возбудимости коры, обусловленном самой частотой вспышек. На различные механизмы формирования (у кроликов) навязанных ритмов при стимуляции реже и чаще 6 – 7 имп/с указывают В.И. Гусельников и А.Я. Супин (1962). По данным Р.М. Мещерского и Г.Д. Смирнова (1961), реактивные потенциалы, возникающие при различных частотах световой стимуляции, имеют отличия в своей структуре и, возможно, связаны с возбуждением различных элементов коры и с различной степенью участия неспецифических таламических ядер. Японские авторы (Z. Kaneko et al., 1961) наблюдали на кошках противоположные сдвиги навязывания более медленных и более частых ритмов в условиях стимуляции ретикулярных образований.
Все эти данные, как мы полагаем, идут в том же направлении, что и результаты, только что рассмотренные нами. Они свидетельствуют о том, что электрические реакции, формирующиеся в коре при разных частотах стимуляции сенсорного аппарата, не однозначны и, как видно, зависят от различных структурных, функциональных и конституционных факторов.
Обратимся теперь к рассмотрению зависимостей между реакцией навязывания и показателями электроэнцефалогического фона, под которыми мы подразумеваем частоту ритмов, доминирующих в выделенных с помощью фильтров полосах 4 – 7, 8–13 и 14 – 25 Гц, суммарную энергию, измеренную в указанных частотных полосах за 10‑секундные отрезки покоя, и индексы тета-, альфа- и бета-ритмов в частотных полосах, т. е. процент времени, занятого на кривой соответствующим ритмом.
Изучая корреляции между ЭИНР и этими показателями (табл. 12), можно заметить любопытную зависимость, заключающуюся в том, что количество значимых коэффициентов корреляции между навязыванием и показателями фона возрастает по мере понижения частоты ритмической световой стимуляции.
Действительно, среди корреляций фоновых показателей с индексами навязывания в бета-полосе (частоты 16, 18, 20 и 22 имп/с) нет ни одной значимой даже на уровне р < 0,05.
Таблица 12
Коэффициенты корреляции между энергетическими индексами навязывания ритма и показателями ЭЭГ покоя
| эинр, имп/с | Частота | Суммарная энергия | Индекс | ||||||
| тета | альфа | бета | тета | альфа | бета – | тета | альфа | бета | |
| –009 | 329* | –018 | |||||||
| –076 | 429** | 619*** | 507*** | 468** | |||||
| –181 | 429** | 376* | –007 | ||||||
| –117 | –208 | –116 | –066 | –269 | –016 | ||||
| –110 | –124 | –212 | –064 | –067 | |||||
| 500*** | 328* | ||||||||
| –004 | 385** | –055 | 471** | ||||||
| –214 | 354* | ||||||||
| –037 | –078 | ||||||||
| –052 | –064 | ||||||||
| –018 | 331* |
Примечание. • р < 0,1; ** р < 0,05; ***р < 0,01.
ЭИНР двух частот альфа-полосы (11 и 12 имп/с) имеют две значимые положительные корреляции с таким показателем фона, как суммарная энергия альфа-ритма.
Наибольшую связь с показателями фона ЭЭГ обнаруживают ЭИНР частот, лежащих в зоне тета-ритма (5, 6 и 7 имп/с), особенно частоты 6 имп/с. Навязывание этой частоты связано положительной значимой корреляцией с показателями суммарной энергии всех трех ритмов, причем в двух случаях – с энергией тета-ритма и альфа-ритма – эти корреляции (0,619; 0,507) значимы на уровне р < 0,01. Навязывание частот 5 и 7 имп/с также положительно коррелирует с показателями суммарной энергии всех трех ритмов.
Следует также заметить, что, хотя корреляции индексов навязывания каждой из трех групп частот с показателями суммарной энергии соответствующих им ритмов большей частью (кроме корреляций для зоны тета) статистически незначимы, они все же в основном устойчиво положительны и, стало быть, указывают на проявление одной и той же тенденции. Она заключается в наличии положительной связи между эффектом ритмической световой стимуляции и величиной корковой электрической активности. Особенно отчетливо эта тенденция выявляется при сопоставлении показателей корковой активности с индексами навязывания тех частот, которые соответствуют полосе тета-ритма (5, 6, 7 кол/с).
Исходя из эиих данных, можно предполагать, что и величина электрической активности коры, и навязывание медленных ритмов, по-видимому, хотя бы частично определяются одним и тем же фактором деятельности нервной системы, действующим в направлении положительной связи между двумя указанными феноменами.
В этом последнем пункте наши данные, видимо, расходятся с материалами Э.А. Голубевой (1965), включившей в свое сопоставление один из показателей фона ЭЭГ – альфа-индекс – и получившей отрицательные корреляции между этим показателем и эффектом навязывания медленных частот (2 – 8 имп/с), хотя/ эти корреляции и не всегда значимы (табл. 11). В качестве оценки альфа-индекса Э.А. Голубевой было взято число альфа-волн частотой 10 кол/с, выделенных из исходной ЭЭГ при помощи узкополосного анализатора и подсчитанных посредством счетчика импульсов за определенный промежуток времени. В принципе такой показатель, как альфа-индекс, в определенном смысле (но, конечно, не полностью) подобен суммарной электрической активности альфа-полосы, поэтому корреляции обоих этих показателей с третьим должны были бы иметь одинаковый знак. Однако в двух сопоставлениях знаки оказались противоположными.
Причины этого расхождения до сих пор не выяснены, но можно предполагать, что здесь сыграли свою роль отмеченные выше различия методического характера в способе определения эффекта навязывания, а также особенности метода измерения альфа-индекса, применяемого Э.А. Голубевой. Не исключено также, что это расхождение, возможно, только кажущееся, так как, согласно упомянутому выше предположению Э.А. Голубевой (1963), лучшее навязывание медленных частот у испытуемых с преобладанием возбуждения имеет место постольку, поскольку эти испытуемые имеют склонность к быстрому развитию сонного торможения, а развитие торможения сопровождается улучшением навязывания как раз в области медленных ритмов (Н.Н. Данилова, 1959; Н.Н. Зислина, 1955).
Рассмотрим теперь реакцию навязывания ритма в ее сопоставлении с электрографическими показателями динамичности нервных процессов. Общее представление о характере действующих здесь зависимостей можно получить из графиков, изображенных на рис. 22 и 23, и из табл. 13 и 14. На рис. 22 показаны две гистограммы, построенные по средним ЭИНР для групп с высокой (сплошная линия) и с низкой (пунктирная линия) динамичностью возбудительного процесса, а на рис. 23 приводятся соответствующие гистограммы для групп, различающихся динамичностью торможения (сплошная линия – высокая динамичность, пунктирная – низкая динамичность тормозного процесса). Здесь обращают на себя внимание следующие моменты.

Рис. 22. Реакция навязывания ритма как функция частоты стимуляции в среднем по группам с высокой (сплошная линия) и низкой (пунктирная линия) динамичностью возбудительного процесса.
Ось абсцисс – частота стимуляции (имп/с); ось ординат – эффект навязывания (усл. ед).

Рис. 23. Реакция навязывания ритма как функция частоты стимуляции в среднем по группам с высокой (сплошная линия) и низкой (пунктирная линия) динамичностью тормозного процесса.
Ось абсцисс – частота стимуляции (имп/с); ось ординат – эффект навязывания (усл. ед).
Кривые для групп, различающихся по динамичности возбуждения (рис. 22), проходят почти на всем их протяжении довольно близко одна к другой. Однако на некоторых участках они все же заметно расходятся. Это расхождение очевидно в зоне частот стимуляции, соответствующих бета-ритму, где навязывание оказывается лучше выраженным у лиц с пониженной динамичностью возбуждения. Подобное же различие наблюдается в зоне частот, прилегающих к бета-полосе, – 11 и 12 кол/с, а также частот, входящих в полосу тета-ритма. Из этого правила выделяются лишь частоты 9 и 10 кол/с, навязывание которых имеет (может быть, случайно) противоположный характер. Таким образом, в целом испытуемым с высокой динамичностью возбудительного процесса оказываются свойственны несколько худшие индексы навязывания ритма, чем испытуемым с низкой динамичностью возбуждения.
Значительно более отчетливый характер имеют различия, вскрывающиеся при сопоставлении групповых графиков, построенных для групп с различной динамичностью тормозного процесса (рис. 23). Эти различия касаются только частот, соответствующих тета- и альфа-ритму, в полосе которых очевидно лучшие ЭИНР принадлежат лицам с высокой динамичностью тормозного процесса. В полосе бета-ритма эти различия практически отсутствуют.
Таблица 13

