




Методы испытания работоспособности нервной системы, разработанные в школе И.П. Павлова, как известно, не могут быть прямо перенесены на человека. Этому препятствуют этические соображения, не позволяющие экспериментатору ставить на человеке опыты по определению максимально переносимой дозы кофеина или по исследованию эффекта действия неожиданного, сверхсильного и устрашающего раздражителя. Именно по этой причине разработка методик для определения силы нервной системы у человека со времени смерти И.П. Павлова встречала наибольшие затруднения и была самым\узким местом всей проблемы «типов» в ее приложении к человеку.
Отдельные попытки разрешить эти затруднения с помощью методик, внешне напоминающих некоторые приемы, разработанные в школе И.П. Павлова (например, многократное повторение раздражителя в модифицированной методике речевого подкрепления – Н.А. Рокотова (1954), успеха не имели именно потому, что они лишь внешне, с формальной стороны имитировали эти приемы, обладая совершенно различным внутренним физиологическим содержанием. Между тем методики определения силы, предполагаемые для применения их к человеку, внешне совсем не обязательно должны иметь сходство с приемами определения силы у животных, но они непременно должны сохранять ту нейрофизиологическую суть, которая только и обеспечивает их адекватность поставленной задаче. Создание методик, удовлетворяющих этому требованию, должно основываться не на имитации формы, а на исследовании закономерностей и творческом использовании вытекающих из них следствий.
Примером такого творческого подхода к созданию новых, по форме ничем не напоминающих традиционные методик определения силы, основанных, однако, на использовании ранее установленных закономерностей, является разработка нескольких вариантов методик определения силы нервной системы по особенностям концентрации и иррадиации процесса возбуждения в зрительном анализаторе, осуществленная Б.М. Тепловым и В.И. Рождественской (1955). Речь идет о методике, получившей впоследствии название индукционной.
Индукционная методика
Закономерностью, использованной в этой методике, явилось правило, сформулированное И.П. Павловым на основе многочисленных экспериментальных данных его школы и касающееся поведения возбудительного процесса при нарастании интенсивности стимула. Это правило гласит: «При слабом раздражительном процессе происходит иррадиация, при среднем – концентрация, при очень сильном – опять иррадиация» (1951–1952, т. III, кн. 2, с. 329). В школе И.П. Павлова это правило реализовалось в работе по условноре-флекторной методике, но оно, несомненно, имеет и более широкую сферу применения. В частности, Б.М. Тепловым еще в 30‑x гг. в согласии с этой закономерностью были истолкованы факты, относящиеся к динамике зрительного ощущения как функции интенсивности дополнительного, побочного светового раздражителя (1937, 1941).
Б.М. Тепловым было экспериментально установлено, что допол нительный точечный световой раздражитель, находящийся на определенном расстоянии от тестирующего (тоже точечного) стимула, влияет на порог ощущения этого последнего стимула, причем степень и направление влияния дополнительного раздражителя зависят от его яркости. При яркости, лишь в несколько раз превышающей индивидуальный порог, измеренный в отсутствие побочной стимуляции, наблюдается повышение чувствительности к тестирующему раздражителю, в наибольшей степени – при яркости побочного раздражителя приблизительно в 5 раз больше пороговой. Чувствительность повышается и при подпороговых яркостях дополнительного стимула, вплоть до 1/2 порога. При увеличении яркости побочного раздражителя его положительное влияние ослабляется, и при яркостях, в 9 – 15 раз превышающих порог, оно исчезает. При дальнейшем повышении интенсивности побочного стимула наблюдается уже понижение чувствительности к тестирующему раздражителю.
Согласно предположению Б.М. Теплова, описанная динамика изменения порогов при изменении яркости стимула, находящегося на незначительном угловом расстоянии от тестирующего, весьма точно соответствует первым двум третям приведенного выше павловского правила. Повышение чувствительности при малых интен-сивностях дополнительного раздражителя может быть объяснено иррадиацией возбуждения из пункта корковой проекции того участка сетчатки, на который падает лучистая энергия побочного стимула. Снижение же чувствительности при больших яркостях последнего обязано своим происхождением переходу этих яркостей в разряд «средних» и возникновению вокруг соответствующего пункта корковой проекции зоны отрицательной индукции.
Правдоподобность этой весьма убедительной схемы еще более возрастает, если учесть нейроанатомические данные относительно организации системы «сетчатка – зрительная кора». Согласно этим данным, организация зрительных отделов коры топографически достаточно точно соответствует организации центральных и периферических участков сетчатки (Г. Хартридж, 1952), так что пространственные индукционные отношения между раздражителями, использованные в описываемой методике, видимо, представляют собой реальные пространственные организованные индукционные отношения близко лежащих очагов возбуждения, локализованных, видимо, в области борозды птичьей шпоры.
Однако для того, чтобы обнаруженные соотношения между точечными световыми стимулами могли быть использованы для разработки методики определения верхнего порога функции нервной ткани, необходимо было найти прием, позволяющий зарегистрировать такие закономерные изменения порогов при изменении яркости побочного стимула, которые соответствовали бы третьей части указанного павловского правила. Этим приемом послужило в работе В.И. Рождественской введение различных доз кофеина – вещества, широко использованного в павловской школе для стимуляции возбудительного процесса и для дифференциации на этой основе сильной нервной системы от слабой.
Поскольку кофеин усиливает – и притом дифференцированно, в зависимости от силы нервной системы, – возбудительный процесс, то его эффект теоретически должен заключаться в том, чтобы у слабой нервной системы средний очаг возбуждения превратить в очень сильный, оставив без изменения или с незначительными изменениями средний очаг возбуждения у сильной нервной системы. Применительно к индукционной методике конкретный эффект кофеина должен был выразиться в случае сильной нервной системы в отсутствии всякого действия, а в случае слабой нервной системы – в возникновении вокруг очага среднего побочного стимула, ставшего теперь очень сильным, сверхсильным, зоны иррадиации возбудительного процесса (вместо отрицательной индукции) и соответственно в повышении чувствительности к тестовому раздражителю в присутствии побочного стимула (вместо понижения ее). При этом инверсии должно подвергнуться также действие всех других интенсивностей стимула, которые теперь по своей функциональной интенсивности вынуждены передвинуться на ступеньку выше. Наконец, все эти эффекты, очевидно, зависят от такого существенного фактора, как применяемая доза фармакологического воздействия.
Эксперимент по влиянию точечных световых раздражителей на пороги, поставленный В.И. Рождественской с использованием нескольких доз кофеина, принес именно те результаты, которых можно было ожидать, исходя из предположений о механизмах влияния различных доз кофеина на различные по своей силе нервные клетки. Действительно, наблюдались значительные индивидуальные различия во влиянии каждой из примененных доз кофеина на кривую индуктивных изменений зрительной чувствительности. Действительно, в условиях применения кофеина происходят закономерные сдвиги функционального значения применяемых интенсивностей дополнительного стимула. И наконец, как дифференциация испытуемых, так и сдвиги функционального эффекта в сильнейшей степени зависят от количества принятого испытуемым перед опытом кофеина.
Не останавливаясь на описании деталей экспериментальной установки, процедуры и обработки полученных в опыте данных (В.И. Рождественская, 1955), приведем для иллюстрации обнаруженных фактов несколько рисунков, отражающих индивидуальные особенности изменения индукционных кривых в зависимости от принятой испытуемым дозы кофеина.

Рис. 26. Отсутствие влияния кофеина на форму индукционной кривой. Ось абсцисс – логарифм яркости побочного раздажителя; ось ординат – логарифм отношения величины зрительной чувствительности при дополнительном раздражителе к «фоновой» величине чувствительности (В.И. Рождественская, 1955).
На рис. 26 показан случай, когда ни малая, ни средняя из примененных доз (Coffeinum natr.-benz. соответственно 0,05 и 0,2 г) практически не изменили формы «фоновых» кривых.
На рис. 27 представлен другой случай: малая доза не оказывает влияния на индукционную кривую, а средняя влияет в сторону повышения концентрации и дальнейшего снижения чувствительности при действии сильного побочного раздражителя (интенсивностью около 2,2 лог. ед. от индивидуального порога). Это, так сказать, промежуточный случай.
Наконец, на рис. 28 отражен критический момент: доза 0,2 г вызывает инверсию влияния максимального раздражителя, приводящего теперь к повышению чувствительности, в то время как меньшие интенсивности побочного стимула, без кофеина вызывавшие увеличение чувствительности к тестовому стимулу, теперь вызывают эффект, свойственный ранее только максимальным интенсивно-стям, – приводят к снижению чувствительности.
Есть все основания думать, что индивидуальные особенности «кофеиновой» динамики индукционных взаимоотношений, отображенные на рис. 26–28, представляют собой индивидуальные особенности реализации приведенного выше закона И.П. Павлова на конкретном материале пространственного взаимодействия зрительных ощущений. Если рисунок 26 отражает отсутствие влияния кофеина, характерное – согласно всему опыту павловской школы – для индивидов с сильной нервной системой, то рисунок 28 демонстрирует результаты такого опыта, в котором совершенно отчетливо зарегистрирован момент перехода раздражителя по своей физиологической интенсивности от среднего, вызывающего отрицательную индукцию, к очень сильному, вызывающему иррадиацию, – момент перехода за «порог иррадиации возбуждения» (термин, предложенный Б.М. Тепловым, 1959). Такого рода инверсия действия побочного раздражителя является несомненным доказательством слабости нервной системы, поскольку она может быть определена при помощи кофеиновой пробы.

Рис. 27. Натриобензойный кофеин в дозе 0,2 г увеличивает отрицательную индукцию.
Обозначения те же, что на рис. 26 (В.И. Рождественская, 1955).
Таким образом, кофеин в «докритической» для данного субъекта дозе способен усилить очаг, возбуждения от побочного стимула настолько, что место «первичной» иррадиации от слабых раздражителей займет концентрация, и этот побочный раздражитель при всех своих интенсивностях будет вызывать снижение чувствительности; критическая же для данного субъекта доза вызывает дальнейшее усиление очага возбуждения, так что при значительных интенсивностях побочного стимула возникает «вторичная» иррадиация, свойственная, согласно И.П. Павлову, очень сильным раздражителям и выражающаяся в данном случае в повышении чувствительности к тестовому стимулу. Что же произойдет при дальнейшем допустимом увеличении дозы кофеина до «сверхкритической»? Можно ожидать, что наметившаяся закономерность получит свое дальнейшее развитие: оно должно выразиться в приобретении качеств сверхсильных раздражителей и теми интенсивностями стимула, которые при критической дозе кофеина вели себя как средние. Это предположение полностью оправдывается в опыте: прием чистого кофеина в дозе 0,3 г приводит к тому, что очаги возбуждения от всех интен-сивностей дополнительного раздражителя (даже подпороговых!) становятся функционально сверхсильными, обусловливая одну только иррадиацию и, следовательно, повышение чувствительности независимо от физической яркости этого дополнительного стимула. Примером такого эффекта может служить график на рис. 29, на котором штрих-пунктирная кривая вся проходит выше нулевой линии, указывая на то, что все интенсивности побочной стимуляции вызывают повышение чувствительности к тестовому раздражителю.

Рис. 28. Натриобензойный кофеин в дозе 0,05 г. вызывает увеличение отрицательной индукции, в дозе 0,2 г приводит к появлению иррадиации при высоких яркостях дополнительного стимула.
Обозначения те же, что на рис. 26. (В.И. Рождественская, 1955).

Рис. 29. Чистый кофеин в дозе 0,3 г вызывает иррадиацию возбуждения от дополнительных раздражителей всех примененных яркостей. Сплошная линия – исходная кривая. Пунктирная линия соответствует дозе чистого кофеина 0,05 г., штриховая – 0,1 г, штрихпунктирная – 0,3 г. Значения абсцисс и ординат те же, что на рис. 26 (В.Д. Небылицын, 1956).
Рисунок 29 может служить показательной схемой динамики индукционных кривых под влиянием различных доз кофеина у индивидов со слабой нервной системой. Однако эта схема, конечно, не является стандартной; возможны различные вариации динамики кривых при разных дозах кофеина, и эти вариации, очевидно, будут прежде всего указывать на различные степени слабости нервных клеток. На рис. 27 уже был приведен случай, по-видимому, промежуточного уровня силы нервной системы, когда доза 0,05 г. оказывается неэффективной, а доза 0,2 г приводит лишь к увеличению концентрации в очаге побочного стимула. Но в опыте наблюдаются и в некотором смысле противоположные случаи, когда уже малая доза кофеина вызывает значительный эффект, сопровождающийся инверсией действия побочного стимула и вторичной иррадиацией в зоне больших его интенсивностей, а средняя доза действует так же, как большая у других испытуемых, – выравнивающим образом, приводя к иррадиации от всех яркостей дополнительного раздражителя (рис. 30). Исходя из всей логики действия кофеина как индикатора силы – слабости, можно, очевидно, предположить у данного испытуемого случай весьма выраженной, может быть, крайней слабости нервной системы.
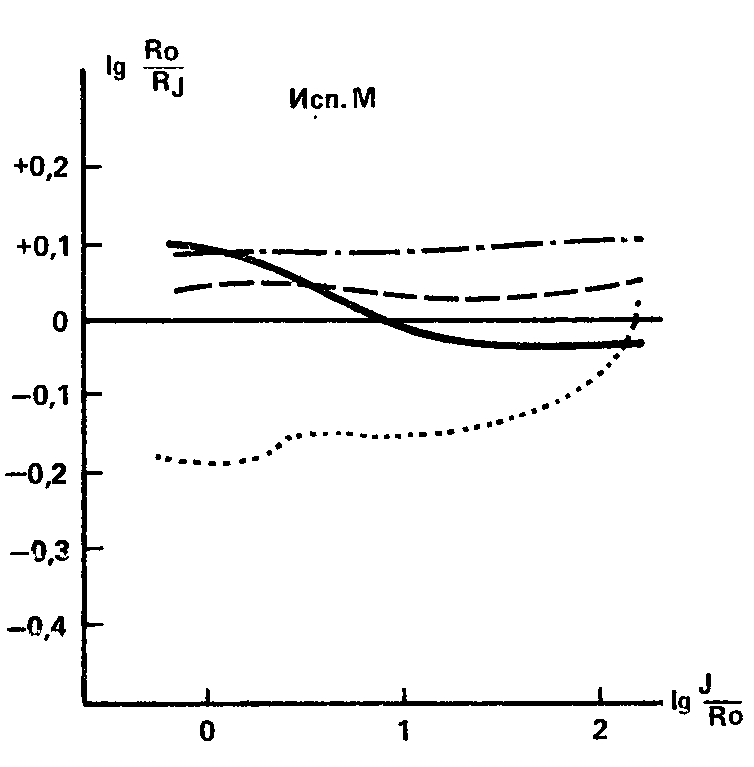
Рис. 30. Иррадиация возбуждения от всех яркостей дополнительного раздражителя вызывается не только максимальной (0,3 г), но и средней (0,1 г) дозой кофеина. Обозначения те же, что на рис. 29 (В Д. Небылицын, 1956).
Все эти данные говорят о том, что определение динамики индукционных кривых в условиях действия различных доз кофеина дает эффективные результаты, позволяя не только квалифицировать испытуемых как «сильных» или «слабых», но и намечать более тонкие градации по параметру силы нервной системы.
Однако применение кофеина, особенно в значительных дозах, не всегда желательно по медицинским соображениям; кроме того, оно удлиняет опыт, а при использовании нескольких различных доз возникает необходимость в организации целой серии экспериментов с обязательными интервалами между опытными днями. Эти соображения побудили В.И. Рождественскую разработать новые варианты индукционной методики, не связанные с необходимостью использования кофеина (1959б).
Один из этих вариантов, так же как и с применением кофеина, основан на усилении очага возбуждения, соответствующего побочному раздражителю, только усиление это достигается более простым и экспериментально более удобным способом: путем многократного предъявления побочного стимула с одновременным измерением чувствительности к тестовому раздражителю. В.И. Рождественская предположила, что повторение дополнительного раздражителя с короткими (порядка 1 мин) интервалами приведет к постепенной суммации возбуждения в клетках зрительного анализатора, соответствующих этому раздражителю, и что у лиц со слабой нервной системой такое накопление возбуждения будет иметь следствием тенденцию к переходу за порог вторичной иррадиации в общем по тому же механизму, что и в случае применения кофеина. Таким образом, многократное измерение порога в присутствии дополнительного раздражителя должно результировать у лиц со слабой нервной системой в постепенной ликвидации эффекта отрицательной индукции от побочного раздражителя и в росте чувствительности к тестовому стимулу в присутствии дополнительного; в то же время у лиц с сильной нервной системой эти эффекты не должны наблюдаться, и падение чувствительности к тестовому раздражителю в присутствии побочного в течение всего опыта должно сохраняться по всей величине приблизительно одним и тем же.
Опыты, в которых чувствительность измерялась 20 раз подряд с интервалами в 1 мин между измерениями, полностью подтвердили эти предположения. В этих опытах в качестве монотонно повторяющегося дополнительного раздражителя служил точечный световой стимул яркостью около двух логарифмических единиц от индивидуального порога. Из предыдущего изложения понятно, что такая интенсивность побочного раздражителя была избрана потому, что при ней скорее, чем при меньших яркостях, можно наблюдать инверсию действия побочного стимула и переход ко вторичной иррадиации. Динамику такого перехода можно видеть на рис. 31.

Рис. 31. Изменения световой чувствительности при многократном предъявлении дополнительного раздражителя.
Сплошная линия указывает на падение индукционного эффекта в течение первых предъявлений дополнительного стимула. Штриховая линия отражает процесс падения зрительной чувствительности при ее измерениях в отсутствие побочной стимуляции.
Ось абсцисс – порядковый номер предъявления стимула; ось ординат – чувствительность (в %) к исходному уровню (В.И. Рождественская, 1959б).
Из приводимых графиков видно, что начиная с первых же измерений порога в присутствии побочного стимула у некоторых испытуемых появляется тенденция к снижению индукционного эффекта, которая достигает максимума обычно в пределах первых же 10 мин опыта (сплошные линии на графиках). При этом можно наблюдать, в сущности, ту же самую инверсию действия дополнительного раздражителя, что и в опытах с кофеином: дополнительный стимул в течение одного, двух или более измерений оказывает положительное влияние на чувствительность к тестовому раздражителю, повышая ее по сравнению с фоном. Вслед за этим, однако, большей частью имеет место процесс постепенного, более или менее заметного увеличения отрицательного индукционного эффекта (снижение сплошных кривых на рис. 31). Причиной этого явления можно, по-ьиди-мому, считать рост утомления, возникающего при частых измерениях чувствительности и интерферирующего с основным процессом – ростом вторичной иррадиации. Доказательством этого предположения могут служить графики, отражающие динамику изменении об-солютной чувствительности при столь же частом ее измерении, но без дополнительного раздражителя. Из этих графиков, пунктирными линиями изображенных на рис. 31, видно, что чувствительность действительно падает при частом повторении измерений и, таким образом, основной процесс развития вторичной иррадиации, вероятно, действительно испытывает, влияние со стороны развивающегося сенсорного утомления.
У другой части испытуемых 20‑кратное измерение порога в присутствии побочного стимула практически никак не влияет на величину индукционного эффекта, а если и влияет, то, скорее, в сторону его возрастания. Иллюстрацией этого могут служить графики на 'рис. 32. Вероятно, и у этих испытуемых можно добиться подъема кривой, но для этого, во-первых, нужно, по-видимому, больше чем 20 измерений (или они должны быть более частыми); во-вторых, возможное снижение индукционного эффекта, очевидно, гак же, как и у испытуемых первой группы, перекрывается влиянием утомления.

Рис. 32. Отсутствие значительных изменений индукционного эффекта при многократном измерении зрительной чувствительности в условиях одновременного предъявления дополнительного раздражителя.
Значения абсцисс и ординат те же, что на рис. 31 (В.И. Рождественская, 1959б).
Таким образом, вариант «повторение» индукционной методики так же успешно дифференцирует испытуемых по некоему качеству нервной деятельности, как и вариант «кофеин». Предварительные теоретические соображения дали В.И. Рождественской существенные основания для того, чтобы трактовать эти различия как обусловленные силой нервной системы по отношению к возбуждению. Позже мы увидим, что такая трактовка приема «повторение» получила неоднократное экспериментальное подтверждение в непосредственных сопоставлениях этого приема с другими методиками определения силы нервной системы.
Физиологические механизмы варианта «повторение» индукционной методики представляются достаточно ясными. Однако нельзя того же сказать о третьем варианте индукционной методики, получившем условное название «утомление» (В.И. Рождественская, 19596). Сущность его вкратце заключается в следующем. У испытуемого многократно (20 раз) измеряется абсолютная зрительная чувствительность, при этом интервалы между измерениями составляют 1 мин вместо обычных 2 – 3 мин. Поскольку тестовым стимулом является точечный раздражитель и деятельность по его обнаружению является достаточно напряженной, вскоре возникает состояние сенсорного утомления, которое сказывается в падении абсолютной чувствительности (примеры этого явления были показаны на рис. 31 пунктирными линиями).
В.И. Рождественская предположила, что состояние утомления, возникающее в соответствующих нервных клетках, должно действовать в направлении снижения предела их работоспособности. Тогда применение индуцирующего раздражителя по окончании серии частых измерений порога должно дифференцировать испытуемых соответственно достигнутой степени этого снижения так же, как в вариантах «кофеин» и «повторение»: у одних испытуемых эффект отрицательной индукции и снижения чувствительности при предъявлении дополнительного стимула должен сохраниться, а у других, менее выносливых, исчезнуть или даже смениться на противоположный.
Опыт показывает, что это предсказание оправдывается и указанные эффекты действительно имеют место, разделяя испытуемых согласно величине и знаку конечного индукционного влияния со стороны побочного стимула. Некоторая неясность остается только в толковании физиологических механизмов происходящих при этом процессов.
Автор методики считает, что в ходе частого применения тестового раздражителя и развития утомления одновременно падает чувствительность и повышается «восприимчивость» тех нервных клеток, которые принимают энергию тестового стимула; в силу этого у лиц слабого типа, более склонных к развитию утомления, предъявление побочного стимула на фоне утомления вызывает переход за порог вторичной иррадиации с положительным влиянием на чувствительность. Другими словами, ответственность за происходящие явления возлагаются целиком на процессы, которые разыгрываются в очаге возбуждения, соответствующем тестовому стимулу, причем толкование этих процессов, как мы полагаем, не свободно от противоречий.
Возможно, более простой и менее противоречивой может быть следующая схема динамики физиологических процессов, лежащих в основе варианта «утомление» индукционной методики. В ходе частых измерений чувствительности в группе клеток, принимающих энергию тестового стимула, развивается состояние торможения (возможно, сходного по своей природе с запредельным). Это торможение в пределах принятой серии измерений тем интенсивнее, чем слабее нервная система, и оно обусловливает большее или меньшее падение чувствительности. Согласно закону отрицательной индукции, в пространстве, прилегающем к очагу развивающегося торможения, возникает зона возбудительного процесса, которая, учитывая близость тестового и побочного раздражителей (45 угл. мин), захватывает и очаг возбуждения от побочного стимула. Интенсивность индуктивного возбуждения тем выше, чем больше интенсивность развивающегося торможения, а значит, тем выше, чем слабее нервная система. В этих условиях суммация возбуждений – наличного, от дополнительного стимула, и латентного, индукционного – приводит в случае слабой нервной системы к переходу очага возбуждения, соответствующего побочному стимулу, за порог вторичной иррадиации и к повышению чувствительности, а в случае сильной нервной системы суммированное возбуждение не достигает порога иррадиации и повышения чувствительности не происходит.
Таким образом, изложенная схема объясняет дифференцирующий эффект варианта «утомление» не повышением восприимчивости (и снижением предела работоспособности) нервных элементов, соответствующих тестовому раздражителю, а главным образом процессами нарастания и последующей суммации возбуждения в очаге дополнительного стимула, с вытекающими отсюда последствиями. Тем самым теория варианта «утомление» сближается с объяснениями физиологических механизмов других вариантов индукционной методики, в которых эффект достигается усилением возбуждения в очаге побочного раздражителя.
Все варианты индукционной методики были неоднократно сопоставлены друг с другом, а также – о чем речь будет идти дальше – с другими методами испытания силы нервной системы. Сопоставления внутри индукционной методики показали, что в общем имеется вполне удовлетворительное совпадение результатов, полученных разными приемами. Первоначальное сопоставление, проведенное в ходе разработки вариантов методики (В.И. Рождественская, 1959б), было выполнено методом сравнивания групповых результатов: испытуемые были разбиты на три группы («сильных», «слабых» и «промежуточных») согласно результатам испытания по одному из вариантов, и оказалось, что с этим делением в общем совпали результаты испытаний и но другим вариантам.
В указанном сопоставлении, кроме трех вариантов, участвовал еще один показатель, получаемый попутно, в ходе определения индукционной кривой –«фоновая» величина снижения чувствительности при действии дополнительного раздражителя интенсивностью в 2 лог. ед. от индивидуального порога. Было установлено, что этот показатель также может служить индикатором силы нервной системы: большему уровню силы соответствует тенденция к большей отрицательной индукции.
В следующих сопоставлениях, проведенных с использованием математико-статистических методов обработки материала, между показателями индукционной методики была найдена довольно высокая корреляция. Так, в работе В.И. Рождественской с соавторами (1960) были получены для 38 испытуемых следующие интеркорреляции (табл. 20).
Показатель «форма кривой» здесь означает соотношение площадей отрицательной индукции и иррадиации, очерчиваемых индукционной кривой; он в общем аналогичен показателю фоновой индукции при раздражителе в 2 лог. ед. порога. Как видим, он коррелирует с другими показателями примерно на том же уровне, что и эти другие показатели между собой. Следовательно, уже фоновые особенности индукционной кривой могут характеризовать силу нервной системы данного индивида.
Таблица 20
Интеркорреляция рангов показателей индукционной методики (В.И. Рождественская и др., 1960)
| Показатели | ||||
| 1. Форма кривой | 55* | 52* | 59* | |
| 2. Утомление | 55* | 73* | ||
| 3. Повторение | 53* | |||
| 4. Кофеин |
Примечание. • р < 0,001.
В работе З. Г. Туровской (19636) в сопоставление различных показателей были включены два варианта индукционной методики: повторение и утомление. Они дали между собой корреляцию рангов, равную 0,78 (р < 0,001). Оба эти варианта были также сопоставлены в работе М.Н. Борисовой (1965). Корреляция рангов снова была очень высока (р = 0,76; р < 0,001).
Таким образом, индукционная методика дает ряд теоретически обоснованных и хорошо коррелирующих между собой индикаторов силы нервной системы по отношению к возбуждению и может считаться одним из самых надежных референтных методов испытания этого свойства нервной системы у человека.
Угашение с подкреплением
Как отмечают Б.М. Теплов (1956) и В.И. Рождественская (1959а), характеристика силы нервной системы может быть получена путем использования двух основных групп показателей. Одна из них включает индикаторы, испытывающие выносливость нервной системы по отношению к кратковременному (однократному) действию сильных и сверхсильных раздражителей. В другую входят индикаторы, основанные не на интенсивности стимуляции, а на ее продолжительности, на многократном предъявлении раздражителей обычно средней интенсивности.
Хотя некоторые варианты индукционной методики связаны с многократным нанесением стимула, ее, по-видимому, все же нельзя отнести ко второй категории силовых методик: ведь основным критическим испытанием в этой методике является как раз однократное предъявление раздражителя, становящегося в определенных условиях сверхсильным для той, очевидно, ограниченной группы нервных клеток, куда приходит соответствующая афферентная импульсация. Многократное же нанесение раздражителя в вариантах «повторение» и «утомление» только создает условия, в которых побочный раздражитель для определенной части испытуемых становится сверхсильным и вызывает иррадиацию возбуждения.
В отличие от индукционной методики угашение с подкреплением условнорефлекторных реакций прямо измеряет силу нервной системы путем многократного предъявления сочетаний условного и подкрепляющего раздражителей – процедуры, которая сама по себе оказывает истощающее (в функциональном смысле) влияние на нервные элементы и которая сама же, без каких-либо дополнительных методических моментов, дает необходимый количественный индикатор.
Сущность этого метода состоит в предъявлении испытуемому серии сочетаний, в которой интервалы между сочетаниями сокращены по сравнению с обычным опытом. Предполагается, что сближение сочетаний во времени приведет к аккумулированию возбуждения в нервных структурах, являющихся звеньями условной связи, и обусловит более быстрое или более глубокое падение условной реакции у тех индивидов, нервная система которых имеет относительно более низкий порог запредельного торможения.
Эффект угашения с подкреплением в зависимости от организации опыта может быть выражен количественно двояким образом. Когда тест заключается в предъявлении заранее фиксированного числа сочетаний, мерой работоспособности нервных клеток служит отношение величины условной реакции, измеренной по окончании теста, к величине условной реакции, измеряемой перед началом испытания: если это отношение близко к единице или даже превышает ее, то можно говорить о высоком уровне силы нервной системы; если это отношение значительно меньше единицы, то, очевидно, такую нервную систему следует признать слабой. Когда же тест заключается в повторении сочетаний до получения заранее обусловленного числа нулевых условных реакций, то мерой выносливости нервной системы служит число таких сочетаний: чем оно больше, тем нервная система сильнее.
В работах с применением угашения с подкреплением, проведенных в лаборатории Б.М. Теплова, использовался первый из этих двух вариантов.
Подробный анализ условий, влияющих на динамику условных реакций в ходе угашения с подкреплением, и детальный обзор относящейся сюда литературы читатель найдет в статье В.И. Рождественской (1959а), которая экспериментально обосновала возможность использования этого методического приема с целью испытания силы нервной системы по отношению к возбуждению.
В.И. Рождественская указывает, что в свое время этот прием в качестве испытания силы был принят в павловских лабораториях, но затем, после работ М.К. Петровой (1934), в ряде случаев не получившей совпадения его результатов с другими силовыми индикаторами, он был оставлен и в «стандартах» больше не фигурировал.
В.К. Федоров (1944), проанализировав возможные – причины падения условных реакций в ходе угашения с подкреплением, нашел, что оно может быть обусловлено: 1) истощением нервных клеток, 2) гипнотизирующим действием однообразных раздражителей, 3) затруднением нервной деятельности вследствие перехода от сложной системы к повторению только одной комбинации раздражителей. Соответственно этому в падении условных реакций могут сказываться как сила нервной системы по отношению к возбуждению, так и сила нервной системы по отношению к торможению (гипнотизирующее влияние), а также инертность нервных процессов (переход от одной системы раздражителей к другой). Следовательно, задача состоит в том, чтобы построить такую систему испытаний, которая была бы свободна от влияния второго и третьего из указанных факторов и по возможности целиком отражала бы действие первого фактора – силы нервной системы по отношению к возбуждению.
В.И. Рождественская указывает, что этого можно добиться при соблюдении следующих условий:
«1) не применять очень слабых раздражителей, которые преимущественно являются причиной гипнотизации;
2) не применять больших интервалов между раздражителями, так как они способствуют гипнотизации;
3) применять раздражители разной интенсивности, а также вызвать усиление создаваемого ими очага возбуждения при помощи кофеина;
4) при снижении величины рефлекса на данный раздражитель проверять величину рефлексов на другие раздражители;
5) опыты с применением одного раздражителя проводить не менее двух раз» (1959а, с. 26).
Соблюдение этих правил позволило В.И. Рождественской разработать и внедрить в экспериментальную практику лаборатории психофизиологии такую весьма эффективную и теоретически безупречно обоснованную методику определения силы нервной системы по отношению к возбуждению, как угашение с подкреплением фотохимических условных реакций.
«Фотохимический» вариант угашения с подкреплением. В методической серии опытов, проведенных В.И. Рождественской, участвовали только 7 испытуемых, однако этого было достаточно, чтобы показать, что исследуемая методика вполне четко дифференцирует испытуемых и что существуют все основания приписать полученные различия влиянию силы нервной системы относительно возбуждения.
В этих опытах условнорефлекторное понижение абсолютной чувствительности вырабатывалось вначале на тихий тон 500 Гц и вместе с этим – для большей концентрации возбуждения в очаге условного раздражителя – вырабатывалась дифференцировка на тон 350 Гц. После того как устанавливалось наличие достаточно прочной условной реакции, проводился первый опыт по угашению с подкреплением.
Этот опыт складывается из следующих моментов: пробы наличия условной реакции путем предъявления изолированного условного сигнала, 10‑кратного повторения сочетаний условного и подкрепляющего раздражителей с короткими интервалами между сочетаниями (2 мин вместо обычных 4 – 6 мин), измерения фоновой чувствительности и, наконец, новой пробы наличия условной реакции по отношению к только что измеренному «фону», которой иногда после процедуры опыта устанавливается на новом (обычно более низком) уровне.
После проведения этого опыта громкость условного и дифференцировочного раздражителей значительно повышалась и происходила новая выработка положительной и тормозной условных реакций. По ее окончании снова следовал опыт на угашение с подкреплением, по своей структуре полностью повторявший только что описанный. Затем были проведены третий и четвертый опыты, отличавшиеся от второго только тем, что в них за полчаса до начала теста испытуемые получали кофеин (в третьем опыте – 0,1 г чистого кофеина в растворе, в четвертом – 0,2 г).
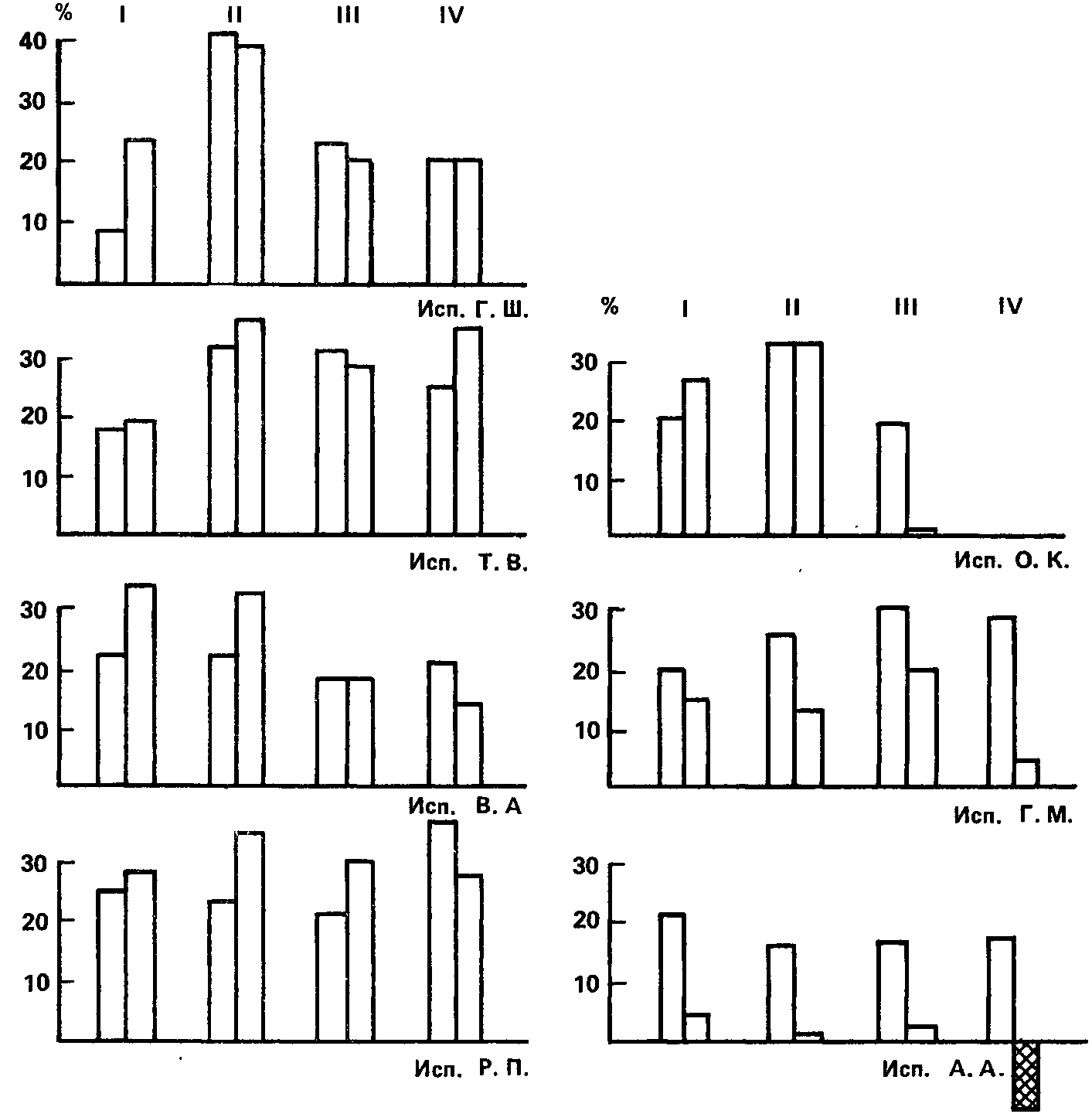
Рис. 33. Угашение с подкреплением условных фотохимических реакций. Римскими цифрами обозначены номера контрольных опытов. Высота каждого столбца означает величину условной реакции (в % снижения чувствительности). Первый столбец в паре показывает величину реакций в начале данного контрольного опыта, второй – в конце его, после десятикратного подкрепления с сокращенными интервалами между сочетаниями.
Результаты этого эксперимента показаны на рис. 33. Здесь итоги четырех опытов с каждым испытуемым (кроме одного) представлены в виде четырех пар столбиков, из которых левый обозначает величину условной реакции в начале опыта, а правый – по окончании серии сочетаний. Можно видеть, что у испытуемых, чьи данные показаны в левой части рисунка, величина условной реакции (в % снижения чувствительности относительно фона) лишь в отдельных случаях снижается после угашения с подкреплением, причем это снижение настолько мало, что лежит в пределах ошибки измерений (10 %), и к тому же, заметим, приходится в трех случаях из четырех на опыты с кофеином. Что же касается остальных трех испытуемых (правая часть рисунка), то у них, особенно у Г.М. и А.А., процедура угашения с подкреплением приводит к падению величины условных реакций, и тем большему, чем более форсируются условия опыта. Последнее обстоятельство явно свидетельствует о том, что данной формой испытания определяется именно сила, а не какое-нибудь другое свойство нервной системы.
Таким образом, в изложенной работе угашение с подкреплением было достаточно убедительно обосновано как испытание силы нервной системы по отношению к накапливающемуся концентрированному возбуждению. С тех пор эта методика была использована в целом ряде работ, выполненных в лаборатории Б.М. Теплова (Л.Б. Ермолаева-Томина, 1959, 1960; В.И. Рождественская и др., 1960; В.Д. Небылицын, 1957а, 1959а).
Проведенные на гораздо большем числе испытуемых, эти работы уточнили некоторые детали методики и обогатили наши представления о ее возможностях и об отдельных ее сторонах. Так, было найдено, что применение в качестве условных раздражителей стимулов различной сенсорной модальности приводит к характеристике силы нервных клеток именно того анализатора, к которому адресуется условный раздражитель; это открывает возможность исследования силы как парциального качества нервной системы (В.Д. Небылицын, 1957а). Этому вопросу ниже будет посвящена специальная глава (см. гл. XII). Из других моментов можно отметить значительное число случаев, когда у лиц со слабой нервной системой угашение с подкреплением не только результировало в падении рефлекса до нуля, до уровня фона, но было более глубоким, проявляясь в повышении абсолютной чувствительности, т. е. в «реакции обратного знака» (В.И. Рождественская и др., 1960). Это явление, очевидно, указывает на особенную глубину развивающегося запредельного торможения.
Фотохимическая методика, обладая рядом достоинств, в то же время не лишена и ряда существенных недостатков, среди которых, как указывает Б.М. Теплов (1956), большая трудоемкость метода и невозможность непосредственного наблюдения эффекта условно-рефлекторного раздражителя, особенно динамики его развития во времени. К этому можно добавить еще и отсутствие объективной регистрации условного эффекта – изменения зрительных порогов. Поэтому важной задачей было разработать экспериментальную технику, основанную на том же принципе, т. е. исследующую способность нервных клеток выдерживать длительное концентрированное возбуждение, но лишенную указанных недостатков. Опыт или цикл опытов должен занимать минимальное время, должно быть обеспечено непосредственное наблюдение и измерение условнорефлекторного эффекта и его динамики во времени, и, наконец, должна производиться объективная регистрация всего хода опыта.
Этим условиям, как мы полагаем, вполне удовлетворяет техника угашения с подкреплением условнорефлекторной депрессии альфа-ритма электроэнцефалограммы человека (В.Д. Небылицын, 1961 д, 19656).
Электроэнцефалографический вариант угашения с подкреплением. При разработке этой методики пришлось преодолеть принципиальную трудность, заключающуюся в чрезмерной нестойкости и лабильности условнорефлекторной блокады альфа-ритма.
Условная десинхронизация, будучи у многих (но отнюдь не у всех!) лиц легко вырабатываемой, в то же время и очень легко исчезает. Причина этого лежит в индифферентности светового стимула, обычно применяемого в качестве подкрепления, в отсутствии у него сигнального, содержательного значения. Эта же причина обусловливает и случаи необразования электрокорковых условных рефлексов, когда условная реакция депрессии альфа-ритма не может быть получена, несмотря на большое число сочетаний.
Нужно было, однако, иметь гарантию того, что исчезновение условной реакции в ходе угашения с подкреплением не является простым следствием утери подкрепляющим раздражителем того сигнального значения, которое он в силу своей новизны имеет в начале опыта. Выход здесь был найден в том, что испытуемым в качестве подкрепления предъявлялись не простые световые раздражители, а различные – чаще всего сюжетные – изображения, с инструкцией запомнить показанное и быть готовым ответить на вопросы после опыта (В.Д. Небылицын, 1961 г.).
При использовании этого подкрепления, названного нами активирующим, условнорефлекторная блокада альфа-ритма у подавляющего большинства испытуемых оказывалась очень стойкой. Это дало нам основания полагать, что изменения в величине условно-рефлекторных реакций, наблюдаемые в ходе угашения с подкреплением, не являются артефактом методики, а происходят действительно как следствие закономерного процесса перехода нервных клеток в состояние запредельного торможения, обусловленного истощением функциональных клеточных ресурсов.
Техника угашения с подкреплением в ее электроэнцефалографическом варианте состоит в следующем. У испытуемого вырабатывается условный рефлекс, заключающийся в том, что после ряда сочетаний условного раздражителя, ориентировочное действие которого было предварительно угашено, с подкрепляющим стимулом одно изолированное действие условного стимула вызывает депрессию альфа-ритма.
В наших опытах условным раздражителем служит прерывистый звук 500 Гц средней громкости, частотой в 10 посылок в секунду и длительностью 4 с, подаваемый через динамик от фотофоностимуля-тора ФФС‑01. Испытуемый лежал с открытыми глазами в звуконепроницаемой камере в темноте. Перед глазами у него был укреплен белый экран, на который через окно в стенке камеры проецировались кадры кинопленки. В течение всех 50 сочетаний на экран предъявлялось одно и то же изображение. Инструкция требовала от испытуемого быть чрезвычайно внимательным до самого конца опыта и стараться запомнить мельчайшие детали изображения, с тем чтобы по окончании опыта ответить на вопросы экспериментатора. Применялось биполярное затылочно-височное отведение. Регистрация биотоков производилась при помощи четырехканального чернильно-пишущего электроэнцефалографа. В опытах участвовали 22 испытуемых в возрасте от 20 до 26 лет.
Применение активирующего подкрепления приводило в подавляющем большинстве случаев к весьма быстрому (после 2 – 5 сочетаний) образованию условного рефлекса и к стойкому его сохранению в опытах, предшествовавших угашению с подкреплением. После выработки дифференцировки на прерывистый тон 250 Гц ставились опыты на угашение с подкреплением. В этих опытах испытуемым предъявлялось 50 сочетаний условного и подкрепляющего стимулов, с интервалами в 4 – 5 с между сочетаниями (в обычном опыте этот интервал составляет 15 – 20 с).
Для наблюдения за динамикой условной реакции после 3, 7, 10, 15, 20 и т. д. сочетаний предъявлялся звук без подкрепления – всего, таким образом, 11 раз, причем пропуски подкрепления делались не точно через 5 сочетаний, а через 4 – 6, во избежание образования стереотипа. Вся процедура занимает около 15 мин.
В следующем опыте этот тест был повторен с применением более громкого (на 20 дБ) звукового раздражителя. Использование более интенсивного условного раздражителя должно было углубить эффект угашения с подкреплением и тем самым подтвердить ценность этого испытания как приема определения силы нервной системы.
Третий опыт являлся точной копией первого; он ставился с целью получения коэффициента надежности данной пробы, т. е. меры постоянства результата при повторении испытания.
Наконец, в четвертом опыте испытуемые за 30 мин до начала эксперимента получали 0,2 г чистого кофеина в водном растворе. Применение кофеина должно было, как и применение более громкого звукового раздражителя, увеличить эффект угашения с подкреплением.
Обработка результатов осуществлялась следующим образом. Показателем величины условной реакции служила длина отрезка депрессии альфа-ритма в секундах в период действия условного раздражителя; следовое действие стимуляции не учитывалось. Поскольку величина условных рефлексов под влиянием различных причин подчас сильно меняется от измерения к измерению, было признано целесообразным использовать для обработки не индивидуальные измерения, а средние из нескольких (трех) проб, идущих подряд, именно из 1 – 3, 4–6, 7–9 и 10 – 12 проб, причем первое измерение производилось до начала угашения с подкреплением. О величине изменения условного рефлекса в ходе опыта мы судили, вычисляя процентное отношение каждой из средних за 4 – 6, 7–9 и 10 – 12 измерений к средней за первые три измерения.
Полученные таким образом индивидуальные кривые представлены на рис. 34, где отражены результаты первого опыта.
Из этого рисунка видно, что у группы испытуемых величина условной реакции практически не изменилась или только немного снизилась к концу опыта. Это дало нам основание, имея в виду опыт работы с фотохимическими рефлексами, отнести всех этих испытуемых к сильному типу нервной системы. Сюда же, очевидно, должен быть отнесен и тот испытуемый, который показал значительный прирост величины условной реакции к концу опыта. Группа других испытуемых показала выраженное падение условных рефлексов. Эти испытуемые должны быть отнесены к слабому типу. Наконец, шестеро испытуемых, занявших какие-то средние места, рассматриваются нами как промежуточные по силе нервной системы. Таким образом, было допущено, что повышение, сохранение или падение условной реакции меньше чем до 70 % от фона есть признак достаточно высокой силы нервной системы. Падение реакции больше чем на 50 % от фона было признано свидетельством слабости нервной системы по отношению к возбуждению.

Рис. 34. Индивидуальные графики электроэнцефалографического варианта угашения с подкреплением. Ось абсцисс – порядковый номер проб изолированного условного раздражителя, сгруппированных по три; ось ординат – процентное отношение средних величин условных реакций в течение опыта к средней величине условного рефлекса за 1 – 3 измерения.

Рис. 35. Динамика условного эффекта (в процентном выражении) в среднем для всей выборки (жирная кривая) и для групп «сильных» (квадраты) и «слабых» (кружки) испытуемых при угашении с подкреплением (ЭЭГ вариант).
Значения абсцисс и ординат те же, что на рис. 34.
Из рисунка видно, что графики части испытуемых представляют собой ломаные линии. Это указывает на характерную волнообразность процесса перехода клеток в тормозное состояние. Однако такая волнообразность свойственна не всем испытуемым, а главным образом тем, которые обладают сильной нервной системой; испытуемые со слабой нервной системой сразу дают снижение условных реакций, которое почти во всех случаях прогрессирует по мере углубления возникшего тормозного состояния. По-видимому, это отличие не случайно, оно указывает на большую функциональную стойкость сильной нервной системы, которая способна после периода некоторого снижения функции вновь возвращаться к исходному (или близкому к нему) состоянию и которая, таким образом, обладает большим сопротивлением переходу в состояние запредельного торможения.
На рис. 35 показана динамика условного эффекта в его процентном выражении в среднем для выделенных групп «сильных» и «слабых» испытуемых, а на рис. 36 дана динамика опыта с представлением условного рефлекса в абсолютных значениях.
Из рис. 36 видно, что средние кривые первого опыта, обозначенные кружками, начинаются довольно близко от другой (у «слабых» испытуемых средняя величина условной реакции за первые три измерения даже несколько больше, чем у «сильных»). Однако если средняя кривая испытуемых с сильной нервной системой остается после десятков сочетаний практически на исходном уровне или даже имеет тенденцию к повышению, то кривая группы испытуемых со слабой нервной системой сразу и круто идет вниз, а к концу опыта средняя величина реакции для «сильных» испытуемых в 2,55 раза больше, чем для «слабых».

Рис. 36. Динамика условного эффекта (в абсолютном выражении) для групп «сильных» (сплошные кривые) и «слабых» (штриховые кривые) испытуемых при угашении с подкреплением (ЭЭГ вариант).
Кружками обозначен первый опыт, треугольниками – повторный опыт. Ось абсцисс – порядковый номер предъявлений условного стимула, сгруппированных по три; ось ординат – значения длительности условной реакции (в с).
На том же рисунке даны средние кривые третьего опыта, проводившегося, как указывалось, для определения надежности испытания; эти кривые обозначены треугольниками. Можно видеть, что эти кривые весьма близки к соответствующим кривым первого опыта и проходят почти по тем же самым ординатам. Коэффициент корреляции, обычно используемый в качестве меры надежности испытания, равен для первого и третьего опытов +0,661 (р < 0,001). Это говорит о близком сходстве результатов обоих опытов и дает нам право построить по данным этих опытов общие кривые, которые мы и будем далее сопоставлять с кривыми других экспериментов.
Результаты опытов с громким условным раздражителем представлены на рис. 37 кривыми с квадратами. Как уже указывалось, применение более интенсивного условного стимула должно по логике опыта увеличить эффективность пробы, т. е. привести к большему падению условных рефлексов, чем это имеет место в обычном опыте. В таком направлении действовало повышение громкости условного раздражителя в опытах В.И. Рождественской, и то же самое наблюдали и мы в нашем эксперименте (рис. 37).
Таблица 21 Индивидуальные показатели величины условной реакции испытуемых с сильной нервной системой в конце первого опыта и опыта с громким условным раздражителем, с
| Первый опыт | Опыт с громким условным раздражителем |
| U | 2,2 |
| 2,8 | 2,0 |
| 2,0 | 1,4 |
| 2,6 | 1,9 |
| 1,7 | 1,3 |
| 2,4 | 1,1 |
| 2,6 | 2,1 |
| 2,9 | 1,3 |
| 2,2 | 1,9 |
Примечание. x\ = 2,32; х2 = 1,69; t = 3,103; р < 0,01.

Рис. 37. Угашение с подкреплением (ЭЭГ вариант) в условиях действия громкого условного раздражителя (кривая с квадратами) и чистого кофеина в дозе 0,2 г (кривая с крестиками) по сравнению с исходными данными (кривая с кружками).
Обозначения те же, что на рис. 36.
Кривая группы испытуемых слабого типа во второй половине опыта проходит ниже двух кривых, полученных в обычных опытах, что означает уменьшение величины условных рефлексов, очевидно, в результате более мощного энергетического воздействия.
Интересна кривая «сильной» группы. Она имеет значительный излом, указывающий на усиление волнообразности процесса угашения с подкреплением, свойственной испытуемым с сильной нервной системой. После подъема в предпоследней точке кривая круто идет вниз, отражая резкое уменьшение величины реакции в конце опыта. Сравнивая индивидуальные показатели испытуемых, полученные в этом опыте, с показателями предшествующего опыта (для данной точки кривой), обнаруживаем, что снижение условной реакции имеет место у всех «сильных» испытуемых, кроме одного, и что разность соответствующих средних статистически значима (табл. 21).
Разность между средними столь же значима, если сравнивать не абсолютные значения длительности условной реакции, а ее процентные отношения к длительности реакции, измеренной в начале опыта (t = 3,150; р < 0,01). Это указывает на правильность предположения о большей эффективности более интенсивного условного раздражителя и, стало быть, подтверждает «валидность» угашения с подкреплением как приема для определения силы нервной системы.
Интересно, что, хотя у «слабых» испытуемых повышение громкости тоже привело к уменьшению величины условных реакций, это уменьшение выражено значительно менее ясно и статистически незначимо. Вероятно, это можно объяснить тем, что у индивидов со слабой нервной системой реакция была близкой к пределу уже в предшествующем опыте, в условиях менее интенсивного условного раздражителя, так что повышение громкости стимула попросту не смогло у этих испытуемых оказать существенного добавочного влияния на процесс перехода клеток в состояние запредельного торможения. Таким образом, увеличение интенсивности условного стимула приблизило данную функцию к ее физиологическому пределу, но диапазон движения оказался различным в силу различного отстояния функций сильных и слабых нервных систем от предела при исходной интенсивности стимуляции.
Что касается введения кофеина, то предполагалось, что в ЭЭГ варианте угашения с подкреплением, как и в «фотохимическом», кофеин, возможно, приведет к некоторому снижению условного эффекта к концу испытания у «сильных» индивидов и вызовет более раннее и более значительное падение условных реакций у «слабых» испытуемых.
Оба эти предположения не оправдались. Кофеин практически не повлиял на динамику угашения с подкреплением у лиц с сильной нервной системой (рис. 37, сплошная кривая с крестиками) и значительно повысил во второй половине опыта условные реакции у лиц со слабой нервной системой, хотя общая тенденция к снижению эффекта у них осталась (рис. 37, пунктирная кривая с крестиками).
Этот факт, по видимости, противоречит общей концепции снижения порога запредельного торможения под влиянием кофеина, однако он может быть объяснен, если учесть, что кофеин обладает специфическим действием на биотоки мозга, выражающимся в снижении медленной и усилении быстрой активности (А.В. Вальдман, 1963; В.В. Руссев, 1960; Т.Ш. Хапажев, 1962; П.И. Шпильберг, С.И. Субботник, 1960). Это действие, возможно, является результатом подавляющего влияния кофеина на некоторые таламические структуры (P. Krupp et al., 1959; А.В. Вальдман, 1963), в норме оказывающие тормозящее действие на кору больших полушарий.
Если кофеин способствует росту десинхронизации, то не исключено, что это его специфическое действие перекрывает его обычный эффект, выражающийся в снижении порога запредельного торможения. Тогда угашение с подкреплением на фоне кофеина вместо уменьшения условного эффекта будет либо оставлять его на уровне, сходном с «предкофеиновым», либо даже повышать этот эффект.
Именно это мы и наблюдали в наших опытах (рис. 37). При факторной обработке корреляций угашения с подкреплением на фоне кофеина с другими индикаторами силы нервной системы оказалось, что этот тест совершенно не коррелировал с фактором, интерпретированным как сила нервной системы, а вошел в другой фактор, состоявший из показателей десинхронизации ЭЭГ при действии специально световых раздражителей и обозначенный как «альфа-реактивность» (В.Д. Небылицын, 19636). Это подтверждает только что высказанное предположение о характере действия кофеина в ЭЭГ варианте угашения с подкреплением и заставляет, таким образом, отказаться от применения в этой методике кофеина в качестве дополнительного индикатора силы нервной системы.
Весь приведенный материал в целом свидетельствует о возможности использования угашения с подкреплением условных электрокорковых реакций для исследования силы нервной системы по отношению к возбуждению. Однако специфика электроэнцефалографических условных реакций – в смысле их склонности к исчезновению – такова, что даже использование активирующего подкрепления не снимает полностью сомнений в том, что изменения условно-рефлекторного эффекта зависят именно от силы нервной системы, нельзя исключить того, что здесь может играть определенную роль и постепенное снижение ориентировочной активности – ведь испытуемому в качестве подкрепления показывался все время один и тот же кадр.
Поэтому во втором цикле работы по исследованию ЭЭГ варианта угашения с подкреплением, проведенном на 20 новых испытуемых, в методику были внесены некоторые изменения, которые должны были способствовать поддержанию ориентировочной активности испытуемого в течение всего опыта на одном и том же уровне. Именно кадры кинопленки, служившие подкреплением, теперь сменялись через каждые 10 сочетаний, с инструкцией испытуемому запомнить последовательность кадров и содержание каждого из них. В связи с этим общее число сочетаний, в предвидении более замедленного процесса падения условного эффекта, было увеличено до 80. Кроме того, условным раздражителем служил теперь не прерывистый, а сплошной тон 500 Гц. В остальном методика осталась без изменений.

Рис. 38. Изменение длительности условной реакции десинхронизации в ходе угашения с подкреплением (ЭЭГ вариант).
Рисунок 38 иллюстрирует конкретную динамику условного эффекта у одного из наших испытуемых при применении этого варианта угашения с подкреплением.
Подобно тому как это было сделано в работе первого цикла, по окончательным результатам – процентному отношению средней реакции в последних пяти пробах к средней реакции в первых трех пробах изолированного условного раздражителя – среди испытуемых были выделены «сильные» (с увеличением условного эффекта или небольшим его снижением – не более чем до 80 % от фона) и «слабые» (с окончательным эффектом, меньшим чем 70 % от фона). В первую группу вошли 8, во вторую – 9 испытуемых.
На рис. 39 приведены графики, иллюстрирующие динамику изменения процентного показателя в течение опыта в среднем для выделенных групп, а также для всей выборки испытуемых в целом. Общий график (жирная линия) указывает на то, что в среднем и при данной форме опыта наблюдается некоторое снижение условного эффекта, происходящее равномерно в течение всего опыта. Таким же равномерным по своему характеру является падение условного эффекта, имеющее место у испытуемых со слабой нервной системой (кривая с кружками), у «сильных» же испытуемых снова наблюдается значительная волнообразность процесса угашения с подкреплением (кривая с квадратиками).
Стрелками обозначены начало и конец действия изолированного условного звукового раздражителя. Цифры у образцов записи – порядковые номера предъявлений условного стимула, распределенные в фиксированном порядке среди 80 сочетаний с активирующим подкреплением.

Рис. 39. Динамика условного эффекта при угашении с подкреплением (ЭЭГ вариант; вторая серия опытов). Обозначения те же, что на рис. 35.
Отличие этих кривых от соответствующих кривых, приведенных на рис. 35, заключается в более высоком сохранении условного эффекта как в среднем по всей выборке, так и в отдельности у «слабых» и у «сильных» испытуемых. Несомненно, это отличие связано с видоизменением подкрепления, вызвавшим общее повышение ориентировочной активности и внимания испытуемых и несколько ограничившим развитие тормозного состояния. Заметим, однако, что это ограничение коснулось примерно в одной и той же степени как «сильных», так и «слабых» испытуемых, и поэтому можно считать, что модификация подкрепления не внесла существенных изменений в методику и, следовательно, не имеет заметных преимуществ перед вариантом, описанным выше.
Единственным ограничением электроэнцефалографического варианта угашения с подкреплением является невозможность его применения к тем лицам, у которых альфа-ритм в ЭЭГ отсутствует или характеризуется малым индексом. Эти лица должны быть исследованы при помощи других экспериментальных методик.
Итак, лаборатория психофизиологии располагает сейчас двумя достаточно проверенными методами прямого испытания работоспособности нервных клеток – качества, составляющего главное содержание такого свойства нервной системы, как сила ее по отношению к возбуждению.
В лаборатории получены также данные о соотношении этих методов – индукционного и угашения с подкреплением – между собой. Выяснению этого вопроса была, в частности, посвящена уже неоднократно упоминавшаяся коллективная работа В.И. Рождественской, В.Д. Небылицына, М.Н. Борисовой и Л.Б. Ермолаевой-Томиной (1960), в которой четыре индикатора, извлекаемые из индукционной методики, были сопоставлены в числе прочих с четырьмя индикаторами угашения с подкреплением фотохимической условной реакции (см. также: V.D. Nebylizin, 1962).
Таблица 22
Коэффициенты корреляции рангов между показателями угашения с подкреплением и индукционной методики (В.И. Рождественская и др., 1960)
| Показатели угашения с подкреплением | Индукционная методика | |||
| Форма кривой | Утомление | Повторение | Кофеин | |
| Угашение с подкреплением фотохимического условного рефлекса при зрительном условном сигнале | 54*** | 45** | 56*** | 5Q** |
| То же с применением кофеина | 4д** | 45** | 31* | 31* |
| Угашение с подкреплением фотохимического условного рефлекса при слуховом условном сигнале | ||||
| То же с применением кофеина |
Применение. • р < 0,1; ** р < 0,01; ***р < 0,001.
Соответствующие коэффициенты корреляции приводятся в табл. 22, представляющей собой извлечение из корреляционной матрицы, приводимой авторами. Распределение коэффициентов в этой таблице свидетельствует о том, что, когда оба испытания относятся к одному и тому же (в данном случае зрительному) анализатору, связь между ними достаточно высока. Но и относительно невысокие коэффициенты корреляции «слуховых» вариантов угашения с подкреплением не помешали им при обработке матрицы методом факторного анализа войти с достаточно высокими весами в фактор, обозначенный как сила нервной системы, куда вошли также и почти все другие взятые для сопоставления силовые индикаторы.
Этот последний момент является весьма знаменательным. Он позволил авторам сделать вывод об «однозначности и равной пригодности» этих методик «в качестве методов исследования нервной системы» и «о том, что в основе всех проявлений силы нервной системы лежит единое свойство нервного субстрата» (В.И. Рождественская и др., 1960, с. 47). Далее мы увидим, что этот ответственный и теоретически важный вывод не раз подтвержден в последовавших затем сопоставлениях различных индикаторов силы нервной системы (3.Г.Туровская, 19636; В.Д. Небылицын, 19636; В.Д. Небылицын и др., 1965).
Наличие надежных и адекватных методик сделало возможным проведение целого ряда работ по исследованию как самой природы свойства силы нервной системы, так и той роли, которую это свойство играет в различных функциях и проявлениях организма. Сопоставление с референтными методиками позволило также разработать и ввести в лабораторную практику некоторые более простые, отнимающие меньше времени и более удобные в практическом отношении способы

