




Реакция животного на новизну была впервые исследована и получила название ориентировочного рефлекса в школе И.П. Павлова. Было показано, что возникновение ориентировочного рефлекса не связано с сенсорной модальностью раздражителя, что он может быть подвергнут угашению, причем механизмом последнего является генерация внутреннего торможения, что при всем том он является врожденным, т. е. безусловным, и сохраняется у животных, лишенных коры больших полушарий, приобретая в этом случае особую стойкость и неугасимость (Н.А. Попов, 1921, 1938; С.Н. Чечулин, 1923; И.С. Розенталь, 1929; Г.П. Зеленый, 1930).
Первоначально ориентировочным рефлексом называлась только двигательная реакция животного в сторону нового или необычного раздражителя (поворот головы, движение ушами и глазами и т. д.). Впоследствии получила распространение более широкая точка зрения, согласно которой ориентировочный рефлекс представляет собой целую систему реакций, интегрированных в сложном соматове-гетативном комплексе (Е.Н. Соколов, 1958 а, б; О.С. Виноградова, 1959, 1961). Таким образом, ориентировочная реакция может быть изучена как по моторным, так и по вегетативным и электрографическим индикаторам, которые, впрочем, не всегда согласуются друг с другом (например, скорость угашения различных компонентов ориентировочной реакции может быть различна у одного и того же испытуемого).
Ориентировочная реакция может быть охарактеризована целым рядом параметров, каждый из которых имеет особое функциональное значение, по-видимому, не всегда совпадающее со значением других. Относительно каждого из них можно предполагать ту или иную степень связи с определенными особенностями нервной системы. Каковы эти параметры?
Один из них – порог ориентировочного рефлекса. Поскольку последний всегда есть результат сенсорной стимуляции, постольку возникает вопрос о том минимальном значении стимула, которое вызывает ответ в виде ориентировочной реакции. Многими авторами было установлено, что порог ориентировочного рефлекса (в основном по кожно-гальваническому и электроэнцефалографическому показателям) фактически совпадает с порогом ощущения, определяемым словесной реакцией, во всяком случае, до того, как ориентировочная реакция начинает угасать при повторном предъявлении стимула (Г.В. Гершуни, 1955; A.J. Derbyshire, J. С. Farley, 1959). Но порог ощущения (подробнее см. об этом ниже) обнаруживает связь с силой нервной системы (Б.М. Теплов, 1955; В.Д. Небылицын, 1959 а; В.И. Рождественская и др., 1960). Следовательно, порог возникновения ориентировочной реакции должен коррелировать с показателями силы нервной системы (относительно возбуждения).
К сожалению, до сих пор прямого сопоставления соответствующих индикаторов в эксперименте не было приведено, хотя, вероятно, использование этого приема принесло бы пользу при изучении соотношения между чувствительностью и силой нервной системы у животных.
В типологическом контексте может исследоваться и другой параметр ориентировочной реакции – ее величина. Определение этого параметра представляет некоторые трудности, так как величина ориентировочной реакции закономерно падает по мере повторения предъявлений. Поэтому для учета величины ориентировочного рефлекса нужно пользоваться одним из следующих приближенно отвечающих задаче показателей: 1) величиной реакции на первое предъявление нового стимула, 2) средней величиной реакции на некоторое заранее фиксированное число предъявлений стимула, наконец, 3) характеристикой крутизны кривой, отображающей на графике динамику угасания ориентировочной реакции (градиент функции). Самый простой из этих показателей – первый, и, как мы увидим впоследствии, он достаточно хорошо себя проявляет.
Наконец, третий основной параметр ориентировочной реакции – это скорость угашения ее при продолжающемся повторении стимула. Угашение может производиться до определенного, заранее намеченного критерия, например до отсутствия реакции в серии из трех или более предъявлений подряд (острое угашение) или до отсутствия реакций в нескольких следующих друг за другом опытах (хроническое угашение). Эта процедура сильно напоминает угашение условного рефлекса. И.П. Павлов предполагал, что она тоже сопровождается развитием внутреннего торможения (1951–1952, т. IV, с. 269) и, возможно, по физиологическому смыслу означает то же самое, что и угашение условной реакции. Поскольку, однако, ориентировочный рефлекс есть безусловная реакция, многие зарубежные авторы предпочитают вместо термина «угашение» пользоваться терминами «привыкание» и «адаптация».
Как уже было сказано, каждый из перечисленных основных параметров ориентировочной реакции, вероятно, имеет типологическое значение, т. е. находится в зависимости от каких-либо свойств нервной системы. К сожалению, в павловской школе – как при жизни И.П. Павлова, так и после его смерти – не были проведены сколько-нибудь систематические исследования индивидуальных особенностей ориентировочных реакций, равно как и возможной связи этих особенностей со свойствами нервной системы, хотя данные, полученные попутно некоторыми из упомянутых выше авторов, несомненно, давали основания думать, что в ряде особенностей динамики ориентировочного рефлекса отражаются и свойства нервной системы животного. Имеющиеся прямые данные по сопоставлению свойств ориентировочной реакции со свойствами нервной системы могут быть систематизированы следующим образом.
В 1933 г. Н.В. Виноградовым была описана собака слабого типа, которая, по наблюдениям автора, характеризовалась неугасимым ориентировочным рефлексом. С тех пор в литературе (М.С. Колесников, 1953) существует мнение о том, что животным слабого типа нервной системы свойственна неугасающая ориентировочная реакция на любые раздражители внешней среды. Таким образом, согласно этой точке зрения, скорость угашения ориентировки есть функция силы нервной системы.
Другая точка зрения (Л.Н. Стельмах, 1956) связывает быстроту угашения ориентировочной реакции не с силой нервной системы, а с подвижностью нервных процессов (определяемой скоростью переделки). Л.Н. Стельмах указывает, что, с одной стороны, неугасимая ориентировочная реакция может встречаться и у собак сильного типа, а с другой стороны, угашение ориентировки может быть легко достигнуто и у собак со слабой нервной системой. В то же время обнаруживается некоторая зависимость быстроты угашения от свойства подвижности (правда, с существенными исключениями). К сожалению, автор не приводит количественных значений связи между угашением ориентировки и переделкой. Существенным недостатком работы является также то, что исследование ориентировочной реакции было проведено уже после того, как у собак был определен тип нервной системы, т. е. после многомесячной работы с разнообразными внешними раздражителями.
Э.А. Варуха (1953), сопоставляя динамику ориентировочных реакций у собак с результатами определения свойств нервной системы по малому стандарту, нашел, что такой показатель, как изменение величины ориентировочного рефлекса при усилении раздражителя, может быть взят для оценки силы нервной системы (относительно возбуждения), а быстрота угашения ориентировки не связана с силой нервной системы относительно торможения.
Работы, выполненные Л.Г. Ворониным, Е.Н. Соколовым и их сотрудниками (Л.Г. Воронин, Г.И. Ширкова, 1949; Л.Г. Воронин, Е.Н. Соколов, 1955; Е.Н. Соколов и др., 1955; Л.Г. Воронин и др., 1959; У. Бао-Хуа, 1958, 1959), обратили внимание на другой аспект типологической обусловленности ориентировочных реакций, а именно на связь их с уравновешенностью нервных процессов. При этом, как уже указывалось в гл. II, хотя авторы говорят об уравновешенности по силе, анализ применяемых ими испытаний позволяет заключить, что речь идет, скорее, о том, что мы обозначаем как уравновешенность нервных процессов по динамичности. Так, в работе У. Бао-Хуа (1959) референтным показателем уравновешенности служило количество ошибочных действий при выработке элементарного двигательного стереотипа по предварительной инструкции, точнее, соотношение ошибок при предъявлении положительных и отрицательных компонентов стереотипа.
Ни это, ни другие испытания, предусматриваемые методикой Н.А. Рокотовой (1954), примененной в данном случае У. Бао-Хуа, вообще не могут дать показателей силы (выносливости) нервной системы относительно возбуждения, равно как и относительно торможения, но некоторые из них можно трактовать как отражающие уровень динамичности нервных процессов. В большинстве указанных работ речь идет о скорости угашения кожно-гальванических реакций, и предположения сводятся к тому, что быстрое угашение ориентировки по кожно-гальваническому показателю указывает на преобладание тормозного процесса, а замедленное угашение КГР свидетельствует о преобладании возбудительного процесса. То же предположение содержится в работе А. Манди-Касла и Б. Маккивера (А.С. Mundy-Castle, В. Z. МсКiеvеr, 1953), выполненной также с использованием кожно-гальванического показателя.
Итак, разные авторы связывают те или иные показатели ориентировочного рефлекса с различными свойствами нервной системы, причем, как видно, основной интерес вызывает быстрота угашения реакции. Что можно сказать по этому поводу?
Роль силы нервной системы в некоторых характеристиках ориентировочной реакции вряд ли может быть подвергнута сомнению. Мы уже говорили об этом при обсуждении вопроса о пороге возникновения ориентировки. Но величина ориентировочной реакции, по всей видимости, также не может в какой-то степени не зависеть от силы нервной системы относительно возбуждения. Поскольку сильная нервная система обладает менее высокой чувствительностью, зависимость между силой и величиной ориентировки должна быть обратной: индивиды со слабой нервной системой должны обладать более выраженной ориентировочной реакцией, особенно при использовании раздражителей слабой и средней интенсивности, которые в случае систем разной чувствительности обеспечат наибольшие различия в физиологическом эффекте. Возможно, это и является одной из причин более высокой ориентировочной активности, «неугасимого» ориентировочного рефлекса у некоторых особей слабого типа нервной системы – но, вероятно, только одной из причин, причем не самой существенной.
Что касается связи ориентировочных реакций с подвижностью нервных процессов, то имеющиеся материалы (Л.Н. Стельмах, 1956) недостаточны, чтобы сделать по этому вопросу сколько-нибудь определенные выводы. Это, конечно, не означает, что предположение о такой связи должно быть с места отвергнуто. Это означает только, что оно должно быть проверено в экспериментальном сопоставлении соответствующих индикаторов.
Наиболее обоснованными кажутся взгляды, связывающие некоторые параметры ориентировочной реакции с уравновешенностью нервных процессов (мы бы сказали, с уравновешенностью по динамичности). При этом, возможно, следует иметь в виду, что динамичность возбудительного и динамичность тормозного процессов, отражая функционально различные свойства нервного субстрата, могут оказывать различное влияние на разные стороны ориентировочного рефлекса.
Что касается скорости угашения ориентировки, то она, как можно полагать, является прямой функцией динамичности тормозного процесса. Как уже отмечалось, еще И.П. Павлов и его сотрудники указывали, что эффект угашения ориентировочного рефлекса полностью подобен эффекту угашения условного рефлекса: сходство наблюдается как в деталях самих процессов, так и в их результатах – оба они приводят к возникновению дремотного и сонного состояния, обязанного своим происхождением иррадиации вырабатываемого внутреннего торможения.
Анализ электрографических проявлений ориентировочного рефлекса позволил Е.Н. Соколову (1963) и О.С. Виноградовой (1961) выдвинуть предположение о том, что само угашение ориентировочной реакции есть не что иное, как постепенно вырабатываемый условнорефлекторный процесс, в котором условным раздражителем является начало применяемого стимула, становящееся сигналом его определенной длительности и отстутствия его в фоне.
Таким образом, угашение ориентировочного рефлекса ведет к формированию тормозной функциональной структуры так же, как и угашение условной реакции, ведущее, как предполагается, к повышению избирательной активности тормозящих синаптических аппаратов (Е.Н. Соколов, Н.П. Парамонова, 1961; П.В. Симонов, 1962). Так же как и в случае условной реакции, эта тормозная функциональная структура, видимо, складывается прежде всего в коре больших полушарий: удаление коры, согласно данным, полученным еще в школе И.П. Павлова (Г.П. Зеленый, 1930; Н.А. Попов, 1938), и данным новейших работ (М. Jouvet, 1961), приводит к устранению механизма угашения ориентировочной реакции, в результате чего, как указывает Е.Н. Соколов (1963), ориентировочный рефлекс превращается в собственно безусловный рефлекс, лишенный условнорефлекторных компонентов и не поддающийся поэтому угашению.
Основываясь на этих данных и соображениях, мы и предполагаем, что угашение ориентировочной реакции, так же как угашение условной реакции, есть функция главным образом того свойства нервной системы, которое мы обозначаем как динамичность тормозного процесса: высокий уровень динамичности торможения ведет к быстрому угашению ориентировки, при низком уровне этого свойства угашение ориентировки может оказаться весьма длительным процессом. Заметим снова, что последнее явление может, вероятно, быть следствием не только низкой динамичности тормозного процесса, но и высокой абсолютной чувствительности анализатора, воспринимающего сенсорный стимул, который, попадая на данную систему, получает большую физиологическую эффективность; высокая же чувствительность присуща слабой нервной системе.
Некоторые параметры ориентировочной реакции, возможно, зависят и от динамичности возбудительного процесса. В частности, влияние последнего можно предполагать в величине ориентировочной реакции при первых предъявлениях стимула. Действительно, если последующие его предъявления приводят к развитию условного торможения, ограничивающего возникающее возбуждение, то при первом нанесении раздражителя это ограничение еще не выработано или, во всяком случае, недостаточно. Поэтому возбуждение, возникающее при первом предъявлении сигнала, когда механизмы условного торможения еще не вступили в действие, будет, возможно, характеризоваться большей амплитудой, интенсивностью и длительностью. Отсюда у индивидов с высокой динамичностью возбудительного процесса можно ожидать более выраженных (по величине) ориентировочных реакций на первое включение стимула по сравнению с индивидами с низкой динамичностью процесса возбуждения.
По некоторым из высказанных предположений в лаборатории психофизиологии были получены определенные экспериментальные данные. Поскольку эти данные всякий раз имеют свою специфику соответственно использованной методике, рассмотрим их в нескольких разделах, посвящая каждый какой-нибудь одной из примененных методик.
Сенсорные ориентировочные реакции. Специфической чертой сенсорных ориентировочных реакций, т. е. протекающих по правилам ориентировочного рефлекса изменений порогов ощущения (в нашем случае абсолютных порогов), является то, что они кроме указанных выше параметров – порога, величины и скорости угашения – обладают еще и параметром направления: ориентировочная реакция может выражаться как в снижении, так и в повышении абсолютной чувствительности, варьируя в этом своем качестве от испытуемого к испытуемому.
Работой Л.Б. Ермолаевой-Томиной (1957, 1959) это показано с полной определенностью, чем внесены существенные поправки в материалы Л.А. Чистович (1956), отмечавшей при первоначальном действии побочных стимулов лишь повышение абсолютных порогов, и Е.Н. Соколова (1958 а), нашедшего у своих испытуемых только понижение порогов под влиянием раздражителей, вызывающих ориентировочную реакцию.
Л.Б. Ермолаева-Томина исследовала как влияние побочных световых раздражителей (мелькающий свет) на слуховые пороги, так и влияние побочных звуковых раздражителей (прерывистый звук) на зрительные пороги (подробное описание методики см. в указанных работах Л.Б. Ермолаевой-Томиной). Ориентировочный характер воздействия этих раздражителей доказывается, во-первых, тем, что указанные сдвиги угашаются при повторных предъявлениях, во-вторых, тем, что при дальнейших предъявлениях эти сдвиги приобретают противоположный по направлению и теперь уже стационарный характер, и, в-третьих, тем, что ориентировочные сдвиги порогов возникают и при выключении постоянно действующего побочного стимула, а также при изменении порядка раздражителей.
Важно заметить, что проявление найденных закономерностей, по-видимому, не зависит от исследуемого анализатора: если испытуемый имеет тенденцию к снижению слухового порога при действии пульсирующего света, то влияние прерывистого звука на зрительный порог также большей частью выразится у него в снижении измеряемого порога.
Основная корреляция, полученная Л.Б. Ермолаевой-Томиной в сопоставлении со свойствами нервной системы, заключается в зависимости направления ориентировочного сдвига чувствительности от силы нервной системы по отношению к возбуждению. Было найдено, что испытуемые с сильной нервной системой реагируют на первое и последующие (до угашения) предъявления дополнительного стимула, как правило, понижением абсолютной чувствительности, в то время как у «слабых» испытуемых в тех же условиях чувствительность в подавляющем большинстве случаев возрастает. Отдельные исключения, неизбежные при изучении неотобранных групп, лишь подтверждают общее правило.
Но влияние силы нервной системы сказывается не только в направлении сдвигов абсолютной чувствительности. Сравнение средних по группам приводит к заключению о том, что кроме направления сдвигов группы «сильных» и «слабых» испытуемых различаются еще и величиной этих сдвигов: средняя абсолютная величина изменений чувствительности у испытуемых со слабой нервной системой заметно больше, чем у испытуемых с сильной нервной системой.
Таким образом, у «сильных» испытуемых сенсорная ориентировочная реакция протекает по типу внешнего тормоза, в то время как у «слабых» индивидов ориентировочная реакция приводит к улучшению исследуемой сенсорной функции. Эти, по-видимому, парадоксальные результаты нуждаются в объяснении, в качестве которого Л.Б. Ермолаева-Томина выдвигает следующее предположение: «При слабых корковых клетках… ориентировочная реакция вызывает, очевидно, более генерализованное возбуждение, что проявляется в повышении чувствительности анализаторов. Снижение чувствительности при ориентировочной реакции у испытуемых с сильными корковыми клетками можно объяснить, вероятно, тем, что возбуждение у них очень быстро локализуется в том анализаторе, к которому непосредственно адресован экстрараздражитель» (1959, с. 102). С этим объяснением в принципе можно согласиться, если добавить к нему некоторые недостающие звенья, касающиеся главным образом физиологических механизмов указанных различий.
Можно с определенностью думать, что эти различия связаны с различием в абсолютной чувствительности сильной и слабой нервной системы. Слабая нервная система, обладая более низким порогом ощущения, вероятно, имеет и более низкий порог возбуждения неспецифической активирующей системы. Можно предполагать, что в силу этого обстоятельства слабая нервная система дольше сохраняет тонический характер генерализованной активации, обеспечиваемый мезенцефалическим отделом ретикулярной системы.
Напротив, в тех же условиях сильная нервная система с ее более высоким порогом, приводящим к относительному снижению физиологического эффекта, возможно, уже в течение интервала действия побочного раздражителя (20 – 30 с) переходит к фазической форме активации, связываемой обычно с таламической неспецифической системой. А, как известно, особенностью таламической активации является локализация ее в структурах раздражаемого анализатора (S. Sharpless, Н. Jasper, 1956; А.Ю. Гасто и др., 1957; Е.Н. Соколов, 1958 а). Можно представить себе, как это и предполагает Л.Б. Ермолаева-Томина, что в первые моменты действия побочного стимула на сильную нервную систему в ней, как и в слабой, также имеет место генерализованная активация, сопровождающаяся повышением чувствительности к тестирующему раздражителю. Поскольку, однако, она имеет очень краткий характер, экспериментатор просто не успевает измерить и зарегистрировать ее периферический эффект. Через несколько секунд, когда реакция активации уже переведена на таламический уровень и локализована в более узких границах корковых проекций, в зоне анализатора, принимающего тестирующий пороговый стимул, возможно, в силу механизмов последовательной индукции наблюдается падение возбудимости и тем самым понижение чувствительности к тестирующему раздражителю.
Разумеется, все эти соображения носят весьма гипотетический характер и нуждаются в дальнейшем экспериментальном и теоретическом обосновании.
Итак, один из параметров сенсорных ориентировочных реакций – их направление (а может быть, если иметь в виду их величину – и два) – обнаруживает достаточно определенную связь с таким свойством нервной системы, как сила ее по отношению к возбуждению. К сожалению, мы не можем сказать ничего столь же определенного относительно роли, которую играют в сенсорных ориентировочных реакциях другие свойства нервной системы, так как необходимых сопоставлений в лаборатории не производилось, а литературные данные по этому вопросу, насколько нам известно, отсутствуют. В этом отношении больший материал был получен при изучении сосудистых реакций.
Сосудистые ориентировочные реакции. Работа по изучению сосудистых (вазомоторных) ориентировочных и условнорефлекторных реакций была предпринята в лаборатории психофизиологии В.И. Рождественской (1963 б) специально с целью изучения возможностей этой методики в исследовании свойств нервной системы человека. Основная проблема, возникающая при работе с плетизмографической методикой, заключается в трудности установления у многих испытуемых так называемой нулевой плетизмографической кривой, т. е. ровного фона, лишенного спонтанных колебаний. Правда, это как будто больше относится к более чувствительной плетизмограмме пальца, а не руки (А.А. Рогов, 1963), но и в этом последнем случае может наблюдаться выраженная спонтанная волнообразность, маскирующая реакции на раздражители, используемые в опыте.
Нужно указать, однако, что сам характер исходной, фоновой кривой, как это показано В.И. Рождественской и рядом других авторов, может служить показателем такого качества, как уравновешенность возбудительного и тормозного процессов. Возникает вопрос: что это за уравновешенность? Есть ли это баланс в павловском смысле этого термина, т. е. уравновешенность нервных процессов в каких-то высших этажах нервной системы, или, может быть, волнообразность плетизмограммы отражает лишь неуравновешенность динамических сосудосуживающих и сосудорасширяющих влияний, взаимодействующих в подкорковых сосудодвигательных центрах или даже непосредственно на периферии?
Данные В.И. Рождественской свидетельствуют, скорее, в пользу первого предположения. Эти данные были получены на 25 взрослых нормальных испытуемых при регистрации пальцевой плетизмограммы. Программа опытов предусматривала: 1) испытание действия нейтральных звуковых (тон 400 Гц) раздражителей разной интенсивности, 2) испытание эффекта «безусловной» холодовой стимуляции (лед) и 3) выработку условных сосудосуживающих сосудистых реакций путем сочетания звукового раздражителя, ориентировочная реакция на которой была к этому моменту угашена, с подкрепляющим Холодовым агентом.
Таким образом, особенности фоновой кривой и процесса угашения ориентировки могли быть сопоставлены со свойствами динамичности возбудительного процесса, определяемым при помощи сосудодвига-тельной методики. Кроме того, были измерены величина и латентный период реакций на оба вида примененных раздражителей. Итак, что касается ориентировки, то здесь были подвергнуты изучению два ее параметра: величина (средняя из 10 первых предъявлений звука) и скорость угашения.
Особенностью работы было то, что все четыре примененных для угашения ориентировки интенсивности звукового раздражителя (от припороговой до очень сильной) предъявлялись вразбивку и в случайном порядке и, таким образом, можно было сравнивать ход угашения ориентировочной реакции при различных интенсив-ностях стимула. Оказалось (см. табл. 2, заимствованную из работы В.И. Рождественской, 1963 б), что громкость звука весьма существенно влияет на быстроту угашения ориентировки: при очень громком раздражителе критерий угашения (5 тормозных реакций в 5 последовательных предъявлениях данного стимула) не был достигнут у испытуемых из 25, при громком – у 7 испытуемых, при среднем и тихом – только у 1‑го.
Наиболее ясные индивидуальные различия наблюдаются при средней интенсивности раздражителя, на которую у 5 испытуемых не наблюдалось никакой реакции, а максимальное число предъявлений до угашения реакции составляло 20 (у 1‑го испытуемого более 20). По этой причине, а также потому, что условные реакции вырабатывались на раздражитель именно этой интенсивности, для определения связи между скоростью угашения ориентировки и быстротой выработки условного рефлекса мы взяли индивидуальные показатели, полученные при этой средней интенсивности.
Таблица 2
Количество предъявлений звукового раздражителя различной интенсивности до угашения ориентировочной сосудистой реакции (В.И. Рождественская, 1963 б)
| Номер | Характеристика звукового стимула | |||
| ИСП | ||||
| очень громкий | громкий | средний | тихий | |
| Не угасала | Не угасала | Не угасала | Не угасала | |
| » | » | » | ||
| » | » | |||
| » | » | |||
| » | » | |||
| » | » | |||
| » | » | |||
| » | ||||
| » | ||||
| » | ||||
| » | ||||
| » | ||||
| » | ||||
| » | ||||
| » | ||||
Подсчитанный нами коэффициент корреляции между числом предъявлений раздражителя до угашения ориентировки и количеством сочетаний до выработки условной реакции (критерий – 5 положительных реакций подряд в 5 пробах, проводившихся с отставле-нием стимула) равен ‑0,304 (р > 0,1)[28]. Эта величина свидетельствует о том, что у испытуемых с быстрым угашением ориентировочных сосудистых реакций, возможно, существует некоторая тенденция к замедленному образованию условных сосудистых реакций, и наоборот. Однако количественная мера связи между этими параметрами не достигает даже 10 % уровня значимости, и полученная величина может быть случайной. Поэтому ответ на вопрос о связи между динамичностью возбуждения и быстротой угашения ориентировочных сосудистых рефлексов должен быть, по-видимому, скорее, отрицательным. Мнение У. Риза и Р. Дикмана (W.G. Reese, R.A. Dyck-man, 1960) о том, что ориентировочные реакции являются важным индикатором способности к обусловливанию, получает в этих данных лишь весьма слабую поддержку. По-видимому, ближе к истине стоит заключение Дж. Стерна и др. (J.A. Stern et al., 1961 b), полученное, правда, на материале кожно-гальванических реакций, о том, что «реактивность организма в течение обусловливания не может быть выведена на основе учета ориентировочных реакций при предъявлении раздражителя в период, предшествующий обусловливанию» (р. 219).
Что касается другой характеристики ориентировочной реакции – ее величины, то, хотя сравнение групповых средних, обсуждаемое В.И. Рождественской, дает некоторое основание для предположения о связи между динамичностью возбуждения и средней величиной ориентировки, все же подсчет корреляции показывает, что такая связь практически отсутствует: коэффициент корреляции между быстротой выработки условной реакции (до 5 проявлений подряд) и величиной ориентировочных реакций (на громкие звуки) равен только 0,076. Это же относится и к латентному периоду ориентировочной реакции, вычислявшемуся, как и величина ее, для 10 первых предъявлений раздражителя.
Однако между двумя параметрами самой ориентировочной реакции – быстротой угашения и средней ее величиной – существует довольно высокая корреляция. Это было показано в работе В.И. Рождественской и др. (1960), в которой указанные параметры сосудодвигательной ориентировочной реакции на звук и свет, полученные от 38 испытуемых, были сопоставлены с рядом индикаторов силы нервной системы относительно возбуждения.
Коэффициенты корреляции рангов между величиной и скоростью угашения ориентировки на стимулы двух сенсорных модальностей даны в табл. 3, представляющей собой извлечение из таблицы, приведенной в указанной работе. Все корреляции, кроме одной, статистически значимы и указывают, таким образом, на существование положительной связи между числом предъявлений стимула до угашения ориентировочной реакции на него и средней величиной этой реакции. Следует заметить, что эта зависимость в силу каких-то причин выражена гораздо лучше при использовании световых раздражителей (р = 0,84; р < 0,001), чем при использовании звуковых стимулов (р = 0,43; р < 0,01).
Эта разница, однако, не помешала всем показателям, относящимся к ориентировочной реакции, войти при факторной обработке материала в один и тот же фактор, причем с достаточно высокими факторными весами.
Вместе с тем факторный анализ, в согласии с исходным распределением интеркорреляций, показал, что индикаторы ориентировочной реакции не обнаруживают никакой связи с показателями силы нервной системы относительно возбуждения. Это было видно уже из матрицы интеркорреляций, которые бывают то положительными, то отрицательными и во всех случаях, кроме одного-двух, не достигают граничного 5‑процентного уровня значимости. При факторной обработке факторы, представленные показателями силы нервной системы и показателями ориентировочной реакции, оказались различными. Этот второй фактор был обозначен тогда предположительно как уравновешенность нервных процессов; сейчас он, вероятно, был бы идентифицирован более точно, а именно как уравновешенность нервных процессов по динамичности.
Таблица 3
Коэффициенты корреляции рангов между показателями ориентировочных сосудистых реакций (В.И. Рождественская и др., 1960)
| Показатели | ||||
| 1. Средняя скорость угашения ориентировки на зрительные раздражители | 65** | 47** | ||
| 2. Средняя величина ориентировки на зрительные раздражители | 51** | |||
| 3. Средняя скорость угашения ориентировки на звуковые раздражители | 43** | |||
| 4. Средняя величина ориентировки на звуковые раздражители |
Примечание. ** р < 0,01; ***р < 0,001. 106
Итак, подытоживая материалы, полученные при исследовании сосудистых ориентировочных реакций, можно заключить следующее:
1) такие параметры ориентировочной сосудодвигательной реакции, как ее величина и скорость ее угашения, коррелируют между собой;
2) скорость угашения ориентировочной сосудодвигательной реакции обнаруживает лишь весьма слабую связь со скоростью выработки условной сосудодвигательной реакции и тем самым с динамичностью возбудительного процесса, проявляющейся в данной вегетативной функции;
3) ни один из исследованных показателей ориентировочной реакции практически не коррелирует с индикаторами силы нервной системы по отношению к возбуждению.
Кожно-гальванические ориентировочные реакции. Как уже отмечалось выше, кожно-гальванические реакции в силу относительной легкости их регистрации стали одним из излюбленных объектов анализа при изучении высшей нервной деятельности. Особенностью КГР является то, что они во всех случаях возникают только как компонент эмоционально-ориентировочной реакции и являются, таким образом, «наиболее неспецифическими» из всех проявлений ориентировочного рефлекса (Е.Н. Соколов, 1958 а; О.С. Виноградова, 1961). Будучи в качестве такового широко изученным целым рядом авторов (Г.В. Гершуни, 1949; С.С. Мусящикова, 1950; Е.Н. Соколов, 1959; А.С. Mundy-Castle, В. Z. МсКiеvеr, 1953; R. А. Dykman et al., 1959; G. Winokur, 1962; J.D. Montagu, 1963), которые большей частью отмечали весьма значительные индивидуальные различия в величине и скорости его угашения, ориентировочная КГР тем не менее весьма мало исследовалась как функция основных свойств нервной системы. Даже в работах В.С. Мерлина и его сотрудников, широко использующих КГР в качестве основной методики изучения свойств нервной системы, эта реакция исследуется главным образом как условная, а собственно ориентировочная КГР в качестве индикатора каких-либо свойств нервной системы почти не используется.
Попытки связать один из параметров ориентировочной КГР, именно скорость ее угашения, с балансом нервных процессов содержатся лишь в статьях А. Манди-Касла и Б. Мак Кивера (А.С. Mundy-Castle, В. Z. МсКiеvеr, 1953), Л.Г. Воронина, Е.Н. Соколова и У. Бао-Хуа (1959). В первой из этих статей такое заключение делается в порядке предположения, во второй оно обосновывается сопоставлением с результатами выполнения испытуемым относительно трудной двигательной задачи. Авторы обеих работ соотносят замедленное угашение КГР с преобладанием возбуждения, а быстрое угашение – с преобладанием торможения.
Л.Б. Ермолаева-Томина в лаборатории Б.М. Теплова провела несколько циклов работы по изучению возможностей кожно-галь-ванической методики в исследовании свойств нервной системы чело века. В числе изученных индикаторов находилась и скорость угашения ориентировочной КГР, а также величина этой реакции на первое предъявление звукового стимула. Было показано, в согласии с авторами предшествующих работ, что индивидуальность испытуемых сильнейшим образом влияет на скорость угашения ориентировочных КГР. Достижение установленного критерия (три тормозные реакции подряд) происходило у одних испытуемых с первой же пробы, у других растягивалось на много десятков предъявлений.

Рис. 14. Динамика угашения ориентировочной кожно-гальванической реакции (в среднем для всей выборки). Ось абсцисс – порядковый номер предъявления звукового раздражителя; ось ординат – процент испытуемых, реагировавших положительной реакцией.
Из кривой на рис. 14 видно, что на первые предъявления стимула реагируют примерно три четверти испытуемых, затем число реакций в группе уменьшается, но даже на третьем десятке предъявлений приблизительно 10–12 % испытуемых реагируют на стимул положительно (запись по Тарханову с кожи ладони в условиях звукоизолированной камеры). Столь же сильно варьирует от испытуемого к испытуемому и величина реакции на первое предъявление нейтрального стимула. Между собой эти два параметра коррелируют на хорошем уровне значимости (г = 0,683; р < 0,001 для выборки из 24 испытуемых; для более широкой выборки (40 испытуемых) впоследствии были получены значения г = 0,501; р < 0,005).
Сопоставление величины первой реакции и скорости угашения ориентировочной КГР с индикаторами некоторых свойств нервной системы, проведенное на 24 испытуемых в рамках совместной работы по сравнению ряда «коротких» методик (В.Д. Небылицын и др., 1965), приводит к выводам, не лишенным интереса в связи с проблемой структуры основных свойств нервной системы (данные получены и статистически обработаны Л.Б. Ермолаевой-Томиной).
Внутри кожно-гальванической методики мы располагали в указанной работе одним индикатором динамичности возбудительного процесса – скоростью выработки условной КГР (при подкреплении мышечным сжатием) – и одним индикатором динамичности тормозного процесса – скоростью выработки дифференцировки. Сопоставляя параметры ориентировочной реакции с этими референтными индикаторами динамичности, мы находим следующее (см. табл. 4):
Таблица 4
Интеркорреляции показателей кожно-гальванической реакции (Л.Б. Ермолаева-Томина, В.Д. Небылицын и др., 1965)
| Показатели | -3 | |||||
| 1. Число предъявлений звука для угашения ориентировки | –279 | 652** | 683** | |||
| 2. Число сочетаний до выработки условной реакции | –305 | –385 | –039 | –218 | ||
| 3. Число предъявлений до выработки диффе-ренцировки | 597* | |||||
| 4. Величина 1‑й реакции на звук | ||||||
| 5. Величина «безусловной» реакции | ||||||
| 6. Число колебаний потенциала в фоне в среднем за 1 мин |
Примечание. • р < 0,01; ** р < 0,001.
1) скорость угашения ориентировочной КГР на звук не коррелирует значимо со скоростью выработки условной КГР (г = -0,279; р > 0,1), но зато имеет высокозначимую положительную связь с быстротой образования дифференцировки (r = 0,683; р <0,001); таким образом, угашение ориентировки, по-видимому, весьма мало зависит от динамичности возбуждения, но обнаруживает отчетливую зависимость от динамичности торможения;
2) величина (амплитуда) реакции на первое предъявление звукового раздражителя включена в более сложную систему зависимостей: она коррелирует положительно и значимо (г = 0,597; р < 0,01) с числом сочетаний до выработки дифференцировки, а также, как уже указывалось, с числом предъявлений до угашения ориентировки (г = 0,683; р < 0,001); кроме того, величина первой реакции имеет приближающуюся к значимой отрицательную корреляцию с числом сочетаний до выработки условной КГР (г = ‑0,385 р < 0,1). Таким образом, величина первой ориентировочной КГР определяется как динамичностью торможения (отрицательная связь), так и – в некоторой степени – динамичностью возбудительного процесса (положительная связь).
Примерно те же соотношения были получены Л.Б. Ермолаевой-Томиной позже на более широкой группе испытуемых (40 человек), включившей и тех 24 испытуемых, которые участвовали в работе по сопоставлению «коротких» методик (1965). Дополнительные данные, которые здесь получены, касаются отношений между параметрами ориентировочной КГР и показателями баланса по динамичности. О балансе судили по разности между индексами скорости выработки условной реакции и скорости образования дифференцировки (индексы измерялись числом сочетаний): максимальным положительным значениям разности соответствует преобладание торможения, максимальным отрицательным – преобладание возбуждения. Сравнивая показатели кожно-гальванической ориентировки и баланса по динамичности, мы находим, что испытуемые с преобладанием возбуждения (с быстрой выработкой положительной и медленной – тормозной реакции) имеют тенденцию к замедленному угашению ориентировочной реакции (г = 0,474; р < 0,01), а также к большей величине реакции на первое предъявление раздражителя (г = 0,358; р < 0,05). Однако численные значения корреляции не очень велики; это указывает на то, что роль баланса по динамичности в указанных параметрах ориентировочной КГР перекрывается действием других, «первичных» факторов высшей нервной деятельности, в частности фактора динамичности торможения.
Некоторые из отмеченных соотношений подтверждаются материалами других авторов. Так, Дж. Стерн и др. (J.A. Stern et al., 1961 b), как уже отмечалось, на нормальных испытуемых получили отсутствие корреляции между числом реакций при угашении («адаптации») ориентировочной КГР и числом реакций при обусловливании. К сожалению, они не исследовали выработку дифференцировочных реакций, но у них имеется другой показатель динамичности торможения – число реакций при угашении условной КГР. В отличие от только что изложенных данных этот показатель не коррелирует с «адаптацией» к тону. Мы полагаем, что это расхождение можно объяснить тем, что угашение условной реакции проводилось ими не после достижения определенного критерия выработки, а после фиксированного, для всех одинакового числа сочетаний; таким образом, лица с лучшим эффектом обусловливания, естественно, будут показывать большее сопротивление угашению, а корреляция между угашением условной и ориентировочной реакций, естественно, будет близка к корреляции между угашением ориентировки и обусловливанием.
Подытоживая сказанное относительно кожно-гальванической ориентировки, можно сделать следующие выводы:
1) величина (амплитуда) КГР на первое применение звукового раздражителя значимо коррелирует с быстротой угашения кожно-гальванической реакции при повторных предъявлениях этого раздражителя;
2) быстрота угашения ориентировочной КГР обнаруживает высокую положительную корреляцию с быстротой образования дифференцировки и лишь слабую отрицательную корреляцию с быстротой выработки положительной условной реакции. Таким образом, угашение ориентировочной реакции есть функция главным образом динамичности тормозного процесса;
3) величина реакции на первое предъявление раздражителя, напротив, отрицательно связана с быстротой образования дифференцировки, а также обнаруживает некоторую положительную связь со скоростью формирования положительной условной реакции. Таким образом, величина первой реакции связана обратными отношениями с динамичностью тормозного процесса, но при этом обнаруживает некоторую зависимость и от динамичности процесса возбуждения;
4) сравнение обоих показателей ориентировочной КГР с оценкой баланса нервных процессов по динамичности указывает на существование у лиц с преобладанием возбуждения тенденции к замедленному угашению ориентировочной КГР и к более высокой амплитуде первой реакции, а у лиц с преобладанием торможения – обратной тенденции.
Электрокорковые ориентировочные реакции. Основной ЭЭГ компонент ориентировочной реакции – подавление альфа-ритма, реакция активации, – есть непосредственное следствие возбуждения ретикулярной системы среднего мозга с ее генерализованными, сенсорно-неспецифическими влияниями на кору больших полушарий. Хотя ретикулярные структуры среднего мозга, несомненно, принимают участие в интеграции и многих других вегетативных компонентов ориентировочной реакции, электрографическое выражение последней в виде реакции активации есть прерогатива исключительно ретикулярных образований среднего мозга. Этот момент приобретает особый интерес в свете наших психофизиологических построений, отводящих функциям ретикулярной формации существенное место в структуре основных свойств нервной системы (В.Д. Небылицын, 1964 б). Действительно, изучая индивидуальные различия в динамике ЭЭГ ориентировочных реакций, мы можем косвенным образом судить об интенсивности, длительности и лабильности активирующих влияний, генерируемых ретикулярными механизмами среднего мозга.
В соответствии с общим принципом сенсорной неспецифичности ретикулярных активаций последние могут быть вызваны раздражителями любой сенсорной модальности. В наших опытах мы пользовались звуковыми и световыми раздражителями. Оба эти вида стимулов могут вызвать реакцию активации (блокады, подавления альфа-ритма), однако между ними имеется и существенное различие, заключающееся в том, что реакция на звук достаточно быстро подвергается угашению, в то время как при реакции на свет угашению подвергается лишь следовая, десинхронизация, а реакция во время действия светового раздражителя при достаточной его интенсивности может сохраняться неопределенно долго. Это последнее обстоятельство несколько выделяет «световую» активацию из класса ориентировочных реакций, что молча признается большинством авторов, однако не ставит ее особняком настолько, чтобы вовсе исключить ее из этой категории. Действительно, как мы сейчас увидим, разница между ЭЭГ ориентировочными реакциями на звук и на свет имеет скорее количественный, чем качественный характер.
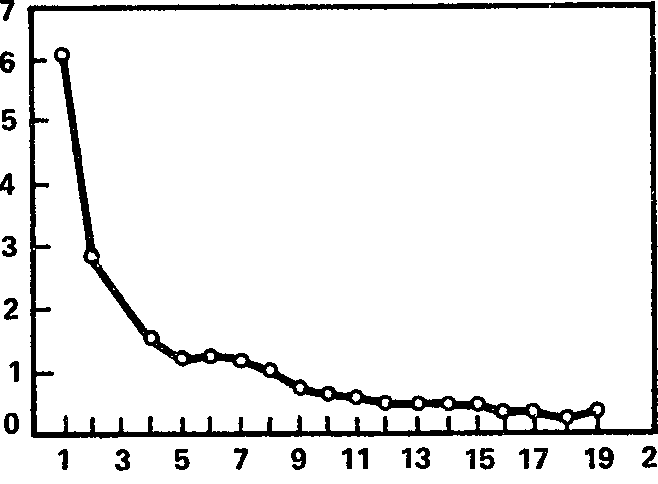
Рис. 15. Динамика угашения ориентировочной электрокорковой реакции на звук (в среднем для всей выборки). Ось абсцисс – порядковый номер предъявления звукового раздражителя; ось ординат – длительность реакции десинхронизации (в с).
Общее представление о динамике ориентировочной реакции на звук можно получить из рис. 15, на котором изображена средняя для выборки из 47 испытуемых длительность блокады альфа-ритма в ответ на последовательные предъявления звукового раздражителя (характеристики раздражителей указаны в гл. 3). Видно, что на первое предъявление звука десинхронизация длится в среднем более 6 с, т. е. к наличной реакции добавляется и следовая (длительность стимула равна 4 с); но уже второе предъявление раздражителя вызывает подавление альфа-ритма, заканчивающееся в среднем уже в течение 3 с, т. е. еще до завершения звукового раздражителя. Примерно с 8 – 9‑го применения стимула длительность реакции является минимальной, составляя в среднем около 0,5 с и менее.
В целом кривая угашения ЭЭГ ориентировки весьма напоминает гиперболу (следует иметь в виду, что она, как и некоторые предыдущие, построена в предположении нулевых реакций у тех испытуемых, которые достигли критерия раньше, чем за 20 предъявлений звукового стимула). Однако эта почти правильная функция слагается из индивидуальных кривых, которые демонстрируют весьма широкий диапазон различий от испытуемого к испытуемому и от реакции к реакции. По нашим данным, угашение ЭЭГ ориентировки на звук требует от одного до нескольких десятков применений раздражителя (в среднем около 9), а длительность реакции на первое предъявление стимула составляет от нуля до 26 с (в среднем около 6 с).
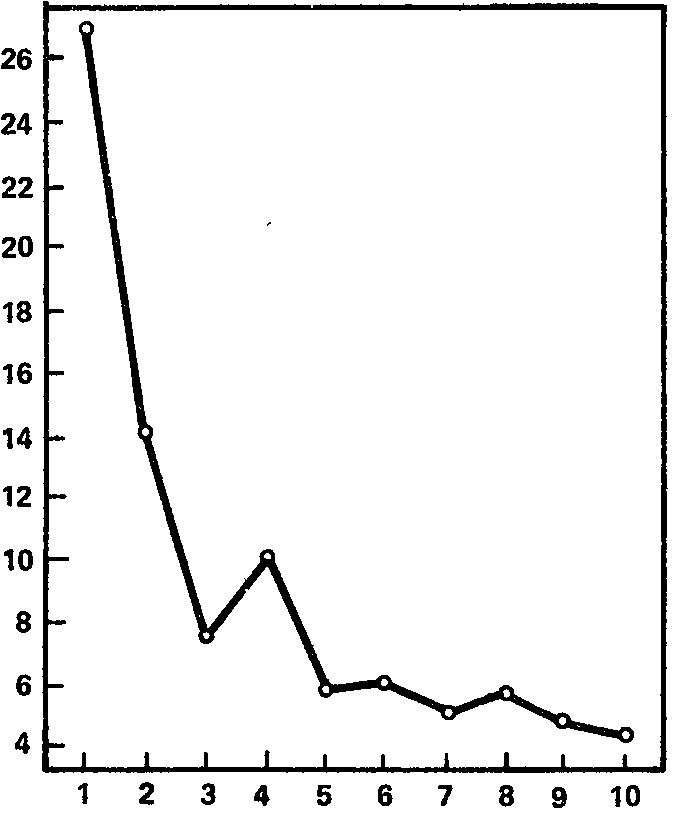
Рис. 16. Динамика ориентировочной электрокорковой реакции на свет (в среднем для всей выборки). Ось абсцисс – порядковый номер светового раздражителя; ось ординат – длительность реакции десинх-ронизации (в с).
На рис. 16 показана динамика ориентировочной реакции биотоков мозга на раздражитель другой модальности, именно на световой стимул значительной яркости (1000 лк при измерении освещенности в непосредственной близости к поверхности излучателя). Измерялось следовое действие раздражителя, т. е. длительность блокады альфа-ритма с момента прекращения светового стимула и до восстановления альфа-ритма (группы альфа-волн длительностью не менее 1 с). Как видно из графика, построенного по данным 20 испытуемых, длительность десинхронизации при первом предъявлении света составляет около 27 с, что примерно в 4 раза превышает длительность реакции на первый звук; подобное же соотношение сохраняется и для каждого из последующих 9 применений светового раздражителя. Однако по своей форме кривая сокращения реакции на свет очень близко напоминает кривую, полученную для звукового раздражителя; это и дает основания думать, что, по крайней мере, следовая десинхронизация на световой раздражитель является по своим механизмам собственно ориентировочной реакцией.
Что касается индивидуальных особенностей реакции ЭЭГ на световой стимул, то они так же разнообразны, как и при действии звукового раздражителя. На первое применение света продолжительность следовой блокады альфа-ритма, по полученным нами данным, составляет от 0,6 до 89,0 с, средняя длительность следовой блокады из 10 предъявлений варьирует у разных испытуемых от 0,8 до 31,1 с, составляя в среднем 8,9 с. Индивидуальные кривые, отражающие динамику сокращения следовой блокады альфа-ритма, показывают, что процесс этот совершается подчас с весьма значительными колебаниями эффекта от стимула к стимулу.
По мнению некоторых авторов (А. Юс, К. Юс, 1954; S. Sharpless, Н. Jasper, 1956; Е.Н. Соколов, 1958 а; О.С. Виноградова, 1961), сокращение активации до длительностей, меньших, чем продолжительность действия раздражителя, связано с переводом активирующих механизмов с мезэнцефалического на таламический уровень: первый обеспечивает длительную, но быстро угашаемую тоническую форму реакции, а второй – более короткую, но и медленнее угашаемую фазическую форму реакции активации.
Признание этой точки зрения необходимо ведет к следующим заключениям: 1) во-первых, нужно согласиться с тем, что активация того или другого уровня, по-видимому, зависит от сенсорной модальности раздражителя, так как при звуковом стимуле переход от тонической активации к фазической совершается (в среднем) уже со второго предъявления (А. Юс, К. Юс, 1958), а при световой стимуляции такого перехода может не наблюдаться даже после многих десятков предъявлений; 2) во-вторых, анализируя индивидуальные данные, следует, очевидно, допустить, что различные индивиды могут характеризоваться различным соотношением двух указанных уровней активации: ведь некоторые из них с первого же предъявления стимула демонстрируют фазическую реакцию, у других же даже при звуковом раздражителе трансформация тонической реакции в фазическую требует нескольких предъявлений.
К сожалению, мы не располагаем материалом, который позволил бы подтвердить или опровергнуть оба заключения; между тем, по крайней мере, второе из них, несомненно, представляет интерес для психофизиологии индивидуальных различий, давая возможность в качестве гипотезы выделить лиц с явно таламическим характером неспецифических активирующих воздействий и лиц, нервная система которых имеет склонность к более генерализованной (топографически и во времени) активации, имеющей преимущественно мезенцефалическое происхождение.
Итак, мы располагаем рядом показателей ЭЭГ ориентировки, полученных в двух сериях экспериментов. Каковы соотношения этих показателей между собой? В табл. 44, помещенной в гл. XI и обсуждаемой там по другому поводу, представлены интеркорреляции данных второй серии (20 испытуемых). Данные этой таблицы свидетельствуют о существовании следующих значимых соотношений: между длительностью первой реакции на звук и длительностью угашения ориентировки на звук (r = 0,493; р < 0,05), а также средней длительностью реакции на свет (в 10 предъявлениях; r = 0,447; р < 0,05); между длительностью угашения ориентировки на звук и длительностью реакции на первое предъявление света (r = 0,665; р < 0,01), а также средней длительностью реакции на световой стимул (r = 0,700; р < 0,001); между длительностью первой реакции на свет и средней длительностью реакции на световой раздражитель (в 10 предъявлениях; r = 0,855; р < 0,001). Иными словами, все соотношения между показателями ЭЭГ ориентировки (кроме одного: между длительностями первых реакций на звук и на свет) оказались статистически значимыми, но заметим, одни – на более высоких, другие – на гораздо более низких уровнях вероятностей нулевой гипотезы. Последнее заставляет предполагать наряду с возможным существованием общего фактора, связанного с протеканием ориентировочных реакций наличие некоторых более дифференцированных влияний на различные параметры ЭЭГ ориентировки, связанных как с модальностью применяемых раздражителей, так и с изучаемой функцией.
Это предположение становится еще более обоснованным, когда мы начинаем сравнивать параметры ориентировочной реакции с некоторыми референтными индикаторами свойств нервной системы. В наших сопоставлениях (В.Д. Небылицын, 1963 б, 1964 в; В.Д. Небылицын и др., 1965) мы имели возможность сравнить показатели ЭЭГ ориентировки, во-первых, с некоторыми индикаторами динамичности, полученными внутри той же электроэнцефалографической методики, а во-вторых, с рядом показателей силы нервной системы по отношению к возбуждению. Заметим сразу, что сколько-нибудь значимой связи между параметрами ориентировки и силой нервной системы нам найти не удалось. Правда, в некоторых случаях длительность угашения ориентировки на звук обнаруживала слабую положительную связь с показателями силы (r = 0,20 – 0,30), но корреляции ни разу не достигали хотя бы 10‑процентного уровня значимости.
Что же касается связи параметров ЭЭГ ориентировки с показателями динамичности, то здесь заслуживают быть отмеченными следующие моменты (табл. 44):
1) длительность реакции десинхронизации на первое предъявление звукового стимула высоко коррелирует со средним условно-рефлекторным эффектом, полученным при «нейтральном» подкреплении (r = 0,709; р < 0,001), в то время как коэффициенты корреляции этого показателя с другими ЭЭГ индикаторами не превышают 0,45. Это – данные второй нашей электроэнцефалографической серии. В первой серии соотношение коэффициентов было примерно таким же, хотя некоторые коэффициенты были несколько выше; корреляция рангов между длительностью первой реакции и средними условнорефлекторным эффектом составила 0,72 (р < 0,001), а для средней длительности условий реакции, выработанной на активирующем подкреплении, – 0,62 (р < 0,01);
2) скорость угашения ориентировки (точнее, число применений стимула до достижения критерия) имеет значимую корреляцию со скоростью угашения условной реакции, выработанной на активирующем подкреплении (r = 0,600; р < 0,01); в то же время корреляция этого показателя со средним условнорефлекторным эффектом не достигает даже 10 %-ного уровня значимости (r = 0,358; р > 0,1). В первой серии соответствующие корреляции были значительно выше – соответственно 0,76 (р < 0,001) и 0,66 (р < 0,01), но между угашением ориентировки и средней длительностью условной реакции, образованной на активирующем подкреплении, корреляция была близка к нулю (0,04);
3) средняя (из 10 предъявлений светового раздражителя) длительность следового подавления альфа-ритма, которую можно рассматривать как индикатор, в определенной степени отражающий длительность угашения ориентировки на свет (чем больше средняя следовая десинхронизация, тем дольше, следовательно, длится процесс привыкания к стимулу), высоко коррелирует с длительностью угашения условной десинхронизации (г = 0,687; р < 0,001); корреляция этого же показателя со средним условным эффектом гораздо ниже и находится на грани значимости (г = 0,449; р < 0,05). В первой серии аналогичный показатель отсутствовал;
4) наконец, длительность блокады альфа-ритма при первом предъявлении света тоже имеет значимую корреляцию с показателем угашения условной реакции (г = 0,622; р < 0,01) и гораздо более низкую – с показателем среднего условного эффекта (r = 0,334; р > 0,1). В первой серии аналогичные корреляции были значимыми (соответственно 0,58, р < 0,01 и 0,69, р < 0,01).
Приведенные соотношения по своему характеру и смыслу близко напоминают те, которые были вскрыты при рассмотрении данных кожно-гальванической методики. Действительно, главный вывод, следующий из анализа таблицы интеркорреляций, заключается в данном контексте в том, что быстрота угашения ориентировочной ЭЭГ реакции на звук и на свет очень хорошо коррелирует с референтным показателем динамичности тормозного процесса – скоростью угашения условной реакции, но довольно слабо (в одном случае – незначимо) коррелирует с референтным показателем динамичности возбуждения – средним условнорефлекторным эффектом. Что же касается длительностей реакции десинхронизации на первые предъявления звукового и светового раздражителей, то их соотношения, как и в случае КГР, выглядят более сложно обусловленными, а кроме того, как видно, зависят от сенсорной модальности стимулов. Если реакция на первое предъявление звука, являющегося нейтральным стимулом для ЭЭГ, высоко коррелирует со средним условным эффектом и слабо – с длительностью угашения условной реакции, то для реакции на первое предъявление света, являющегося специфическим депрессором альфа-ритма, эти соотношения являются, скорее, обратными. Ниже, в гл. XI, при изложении результатов факторного анализа интеркорреляций, представленных в табл. 44, возможные причины этих расхождений будут обсуждены более подробно.
Завершая рассмотрение изложенного в данной главе материала, мы приходим к выводу о том, что параметры любого по физиологической модальности ориентировочного рефлекса организованы в достаточно сложную функциональную структуру, отдельные элементы которой испытывают влияния со стороны различных свойств нервной системы. Так, можно с большой долей уверенности утверждать, что быстрота угашения ориентировочных реакций определяется главным образом динамичностью тормозного процесса как свойством, связанным вообще с быстротой генерации потенциала торможения.
Об этом говорят особенно убедительно результаты, полученные кожно-гальванической и электроэнцефалографической методиками, – высокие корреляции между показателями угашения ориентировки на нейтральный звуковой раздражитель и показателями выработки дифференцировки и угашения условной реакции. Это заключение подтверждается и негативными результатами – слабыми и незначительными корреляциями между угашением ориентировки и образованием положительных условных реакций, зависящим от динамичности возбудительного процесса.
Можно, далее, заключить, что величина реакции на первое предъявление стимула испытывает на себе двойственное влияние как со стороны динамичности возбуждения, так и главным образом со стороны динамичности торможения: первое свойство, видимо, определяет размах, амплитуду, длительность реакции, однако без вмешательства тормозного процесса реакция, возможно, продолжалась бы неограниченно долго. Тормозный процесс выступает в роли ограничителя реакции, и момент его введения в качестве такового определяется как раз уровнем его динамичности. Таким образом, взаимодействие тормозных влияний с возбудительными формирует функцию, обладающую свойством автостабилизации, при этом быстрота развития стабилизирующего эффекта зависит от динамичности тормозного процесса. Конечно, эта схема носит достаточно умозрительный характер, практически же отделить влияние динамичности возбуждения от динамичности торможения в протекании каждой отдельной ориентировочной реакции невозможно.
Далее, в некоторых параметрах ориентировочной реакции можно предполагать влияние силы нервной системы (относительно возбуждения). Это влияние экспериментально показано при изучении индивидуальных различий в направлении сенсорных ориентировочных реакций, и его можно также с определенностью предполагать в величине порога ориентировочной реакции. Нам не удалось выявить отчетливого влияния силы нервной системы в амплитуде ориентировочной реакции, хотя некоторые указания на существование такой зависимости в сенсорных ориентировочных реакциях все же были получены. Скорее всего, влияние силы (чувствительности), если оно имеется, перекрывается влиянием других факторов деятельности нервной системы, более эффективных в отношении данной функции, именно динамичности торможения и динамичности возбуждения. Эти два свойства нервной системы, особенно первое, видимо, являются главными детерминантами основных динамических параметров ориентировочного рефлекса – его величины и скорости его угашения.

