




Ефектори — структури, утворені закінченням аксонів нейроци-тів, які бувають рухові та секреторні. Нервово-м'язові закінчення у скелетних м'язах утворені терміналями аксонів нейроцитів, рухових ядер спинного та моторних — головного мозку. При наближенні до м'язового волокна мієлінове волокно втрачає мієлінову оболонку, його осьовий циліндр розгалужується і разом із саркоплазмою міосимплас-та занурюється у м'язове волокно (рис. 81).
Роздгл 3
Загальна гістологія

Рис. 80. Схема будови нервово-м'язового веретена:
1-моторні бляшки екстрафузальних м'язових волокон; 2-моторні бляш-ки інтрафузальних м'язових волокон; 3, 4-ядерна сумка (скупчення ядер); 5-м'язовий нерв.
У цій ділянці утворюється аксом 'язовий синапс, де аксолема віді-грає роль пресинаптичної мембрани, а сарколема м'язового волокна є постсинаптичною мембраною (рис. 82). Медіатор цих синапсів — аце-тилхолін, який при збудженні вивільняється і через пресинаптичну мембрану проникає у синаптичну гцілину На поверхні постсинаптич-ної мембрани ацетилхолін зв'язується із холінорецепторами і стає хі-мічним сигналом для збудження м'язового волокна.
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія

12 13 9
Рис. 81. Схема будови моторної бляшки:
1-цитоплазма нейролемоцита; 2-ядро; 3-нейролема; 4-осьовий циліндр; 5-сар-колема; 6-кінцеві гілочки нервового волокна в повздовжньому та поперечно-му розрізах; 7-мітохондрії аксоплазми; 8-первинний синаптичний простір; 9-саркосоми; 10-вторинний синаптичний простір; 11-синаптичні пухирці; 12-пресинаптична мембрана; 13-постсинаптична мембрана; 14-ядро моторної бляшанки; 15-міофібрила.

Рис. 82. Ефекторне нервове закінчення (моторна бляшка):
А — вигляд у профіль: 1,2 — терміналі мієлінового нервового волокна; 3-ядра м'язових волокон; 4-м'язове волокно; 5-мієлінове волокно; Б — вигляд звер-ху: 1- мієлінове волокно; 2- безмієлінове волокно; 3-ультратермінальне во-локно (те, що входить до моторної бляшки і закінчується на іншій).
Розділ 3
Загальна гістологія
Еферентні закінчення у гладенькій м'язовій тканині мають прості-шу будову. Окремі нервові закінчення на поверхні гладеньких міоци-тів утворюють варикозні терміналі. Медіатор цих закінчень — ацетил-холін, або адреналін.
Розділ 4
Спеціальнапстолопя
Органи нервової системи
Загальна характеристика. Нервова система виконує координуючу функцію і об'єднує у єдине ціле усі структури організму, а також здій-снює взаємозв'язок із навколишнім середовищем. Це означає, що мор-фофункціональні зміни в одній системі чи органі сприймаються спеці-алізованими структурами нервової системи і передаються у спинний та головний мозок, а звідти нервові імпульси спрямовуються в інші органи та тканини, викликаючи у них реакцію на дане подразнення. Зовнішні фактори навколишнього середовища, такі як світло, тепло, холод, звук також сприймаються нервовими закінченнями і через не-рвові центри передаються до відповідних тканин чи органів. Нерво-ва система відіграє і об'єднуючу роль в організмі за допомогою інших систем і, зокрема, гуморальної. Остання являє собою один із механіз-мів регуляції функції організму, що відбуваються через його рідинне середовище (кров, лімфу, тканинну рідину), в яких знаходяться про-дукти тканинного обміну а також гормони, які здійснюють свій вплив на роботу окремих органів і організму в цілому. А оскільки й органи секреції підпорядковані нервовій системі (у їх будові є нервові струк-тури), то остання і відіграє головну роль в об'єднанні і цілісності орга-нізму Морфологічно така роль нервової системи проявляється у тому, що кожний орган в своєму складі має свої нервові елементи. Отже, як нервова система впливає на морфофункціональні зміни в клітинах, тканинах, органах і системах, так і останні своїми продуктами життєді-яльності здійснюють зворотний вплив на нервову систему — в організ-мі здійснюється нейрогуморальна регуляція систем і органів. Нервова система побудована з нервової тканини, до складу якої входять нейро-ни та нейроглія, а також відростки нейронів, які мають певні шляхи в рефлекторних дугах. Морфологічним субстратом рефлекторної ді-
Розділ 4
Спеціальна гістологія
яльності нервової системи є рефлекторні дуги, які об'єднують ланцюг нейронів, різні частини якого лежать як в периферичних нервових вуз-лах, так і в сірій речовині центральної частини нервової системи.
Отже, нервова система поділяється на: центральну частину, до якої відносять спинний та головний мозок, та периферійну — нервові закінчення, сплетення, ганглії та периферійні нерви, умовне і потрібне лише виходячи з методичних підходів до систематизації викладу мате-ріалу, її топографії та будови.
Залежно від характеру іннервації органів і тканин, нервову систе-му умовно ще поділяють на соматичний та вегетативний відділи. Со-матичний відділ регулює, головним чином, довільний рух і зв'язок з зовнішнім середовищем; вегетативний, в основному — процеси обміну речовин (іннервує внутрішні органи, залози, судини). Морфологічно повністю розділити нервову систему на соматичний і вегетативний від-діли неможливо. В центральній частині нервової системи знаходяться зв'язані між собою центри як соматичних, таіс і вегетативних функцій. В периферійній частині нервової системи, в складі нервів, іннервую-чих сому, знаходяться і вегетативні нервові волокна, а у вегетативних стовбурах проходять соматичні волокна.
Основною функцією нервової системи є рефлекс. Рефлексом на-зивається відповідь (реакція) організму на подразнення рецепторів (нервових закінчень), розміщених як на поверхні тіла, так і всередині його, яка здійснюється через центральну нервову систему. Шлях, по якому проходить нервовий імпульс, що забезпечує відповідь на по-дразнення, називається рефлекторною дугою (рис. 83).
Рефлекторна дуга починається рецептором, від якого імпульси йдуть по доцентровому (еферентному) нерву до чутливих нервових вуз-лів (гангліїв) і далі по мозку, де відбувається трансформація чутливого імпульсу в руховий. Останній по відцентрових (аферентних) нервах до-ходить до м'язів або залоз і викликає відповідну реакцію. Залежно від того, через скільки нейронів пройшло збудження, рефлекторна дуга може бути дво-, три-, чотиринейронною. У більшості рефлекторні дуги вищих хребетних тварин у своєму складі містять ще значну кількість асоціативних нейронів, розміщених між аферентними нейронами.
Методика гістологічного дослідження нервової системи надзвичайно складна, оскільки нервові елементи в тканинах виявити звичайними ме-тодами фарбування повністю неможливо. Для вивчення нервової сис-
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
теми рядом вчених А.С. Догелем, А.Є. Смирновим, Кампосом, Гольджі, Більшовським, Рамон-і-Кохалем запропоновано спеціальні методики. У нейрогістології широко розвинене біологічне моделювання, яке грунту-ється на явищі дегенерації. Це дає змогу вивчати зв'язки між нейронами, тобто описувати рефлекторні дуги. Тепер у дослідженнях нервової сис-теми застосовують нові методи імпрегнації сріблом, свинцювання, різні гістохімічні та біохімічні аналізи, електронну мікроскопію, імунологічні методи, культуру тканини, лазер, авторадіографію.

Рис. 83. Схема рефлекторної дуги простого рефлексу
(за Догелем):
а-чутливі закінчення в шкірі; б-чутливі волокна; в-чутливі клітини; г-спи-нальні ганглії; д-дорсальний корінець; е-рухливі клітини; є-синапси на них волокон чутливих клітин; ж-вентральний корінець; з-рухове волокно; и-ефе-рентні закінчення в поперечносмугастих м'язових волокнах; і-центральна гілка неврита чутливої клітини.
Онтофілогенез нервової системи. Виникнення нервової системи -це результат складного розвитку організму у процесі еволюції. У про
Розділ 4
Спвціальна гістологія
цесі філогенетичного розвитку нервова система зазнала значних змін в бік ускладнення її будови і функції. На перших етапах розвитку бага-токлітинних організмів кожна клітина була здатна сприймати зовніш-ні подразнення і реагувати на них. Спеціальні клітини, які сприймають зовнішні подразнення, вперше з'явилися у кишковопорожнинних. Піз-ніше виникли спеціальні клітини з відростками, які, розгалужуючись, утворюють нервові сплетення. Таку примітивну нервову систему нази-вають дифузною. У індивидів з двобічною симетрією чутливі клітини концентруються і утворюють нервові вузли (ганглії), які з'єднуються між собою за допомогою нервових стовбурів. Таку нервову систему на-зивають гангліозною.
У хребетних органи нервової системи розвиваються із нервової трубки і гангліозної пластинки. Із краніальної частини нервової труб-ки диференціюються головний мозок і органи чуття, а із тулубового відділу її та гангліозної пластинки утворюються спинний мозок, веге-тативні та спинномозкові ганглії, а також хромофільна тканина орга-нізму. Великі півкулі у філогенезі розглядаються як частини головного мозку, що з'явилися пізніше. Вперше великі півкулі, які являють со-бою смугасті тіла, з'являються у риб. Дещо більшого розвитку вони досягають у амфібій, причому у них з'являються перші зачатки кори великих півкуль. У рептилій і птахів останні розвинені дещо краще, а кора являє собою тонкий шар нервових клітин. Значного розвитку кора досягає у ссавців, де вона вкриває всю поверхню мозку, а смугасті тіла, навпаки, займають невелику його частину. Найвищого розвитку кора досягає у людини (95% всієї поверхні головного мозку). Це зу-мовлено тим, що у вищих ссавців, у зв'язку з ускладненнями їх функ-цій, підвищується інтегруюча роль нервової системи, встановлюється тісний її зв'язок з іншими системами організму.
В онтогенезі у ссавців на ранніх стадіях з нейроектодермального зачатка диференціюється нервова пластинка, яка утворює нервовий жолобок, а потім нервову трубку. При замиканні нервового жолобка в нервову трубку, формується гангліозна пластинка, яка розміщується вздовж спинного мозку. Спочатку вона розчленовується на сегменти, у яких диференціюються нейро- та спонгіобласти. 3 нейробластів фор-муються нейрони, із спонгіобластів — нейроглія. Інтенсивно розвиваю-чись, бокові відділи нервової трубки формують дорсальну — крилову пластинку й вентральну — основну. На цій стадії розвитку в бокових
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
стінках нервової трубки розрізняють три зони: внутрішню, яка ви-стеляє спинномозковий канал — епендиму, плащовий шар, із якого в подальшому розвивається сіра речовина, і крайову вуаль, що формує білу речовину спинного мозку. Нейробласти вентральної зони плащо-вого шару — передніх стовпів — диференціюються у рухливі нейрони ядер вентральних рогів, а їх аксони утворюють вентральні корінці. Не-йрити чутливих нейронів відходять у бік дорсальних відділів спинного мозку, утворюючи чутливі дорсальні корінці спинномозкових нервів. Просвіт нервової трубки значно звужується, перетворюючись у цен-тральний спинномозковий канал. 3 переднього (розширеного) кінця нервової трубки розвивається головний мозок. Спочатку утворюються три первинних мозкових міхурі — передній, середній і задній. Передній і задній, в свою чергу, діляться надвоє, у результаті чого утворюються п'ять мозкових міхурців, з яких розвиваються усі відділи головного мозку. 3 двох передніх мозкових міхурців утворюються великі півкулі мозку і проміжний мозок, із середнього розвивається середній мозок, а з двох задніх мозкових міхурців — мозочок, вароліїв міст і довгас-тий мозок. Після утворення трьох вигинів стінки мозкових міхурців посилено розростаються з подальшою диференціацією нейробластів і спонгіобластів у нейрони та нейроглію.
Спинний мозок
Спинний мозок розміщений в спинномозковому каналі і складаєть-ся із двох симетричних половин, розділених вентральною серединною борозною і дорсальною серединною перегородкою. Обидві половини з'єднуються між собою за допомогою сірої комісури, де знаходиться центральний канал спинного мозку (рис. 84).
Спинний мозок, як і головний, оточений трьома сполучнотканин-ними оболонками: твердою, павутинною і м 'якою, які утворюють щі-линоподібні підоболонкові простори (субдуральний і субарахноідаль-ний). Каудальний кінець спинного мозку переходить в кінцеву нитку, а краніальний — безпосередньо в довгастий мозок. Центральну части-ну спинного мозку становить сіра речовина, що містить мультиполяр-ні нервові клітини, які утворюють ядра спинного мозку Периферійна частина його-біла речовина, утворена сукупністю нервових волокон, що входять до складу різних провідних шляхів центральної нервової системи.
Розділ 4
Спеціальна гістологія
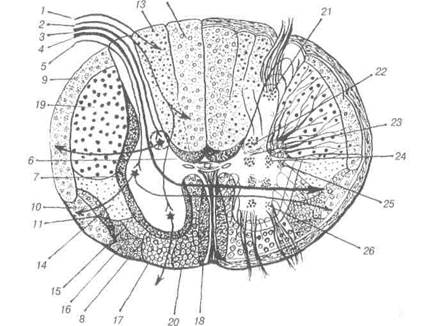
Рис. 84. Схематичний розріз спинного мозку:
1-2 — рефлекторні шляхи свідомих пропріоцептивних відчуттів та дотику; 3-4 — рефлекторні шляхи пропріоцептивних імпульсів; 5 — рефлекторні шляхи температурної та больової чутливості; 6-дорсальний власний пучок; 7-латеральний власний пучок; 8-вентральний власний пучок; 9-дорсальний та 10— вентральний спиномозочкові шляхи; 11-спинно-таламічний шлях; 12-ніжний пучок; 13-клиноподібний пучок; 14-руброспинальний шлях; 15-таламо-спинальний шлях; 16-весгибуло-слинальний шлях; 17-ретикуло-спинальний (пірамідний) латеральний шлях; 20-кортико-спинальний (піра-мідний) вентральний шлях; 21-власне ядро дорсального рогу; 22-грудне ядро Кларка; 23,24 — ядра проміжної зони; 25-латеральне ядро (симпатичне); 26-ядра вентрального рогу.
Сіра речовина має вигляд метелика з розправленими крилами, се-редню її частину називають сірою спайкою (комісурою). Кожна поло-вина сірої речовини має дорсальні та вентральні роги, в грудних і по-перекових сегментах спинного мозку можна виділити верхньобоковий відділ вентральних рогів, як латеральні роги. Сіра речовина утворена
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
мультиполярними нейронами, мієліновими (м'якотними) і безмієлі-новими (безм'якотними) нервовими волокнами і нейроглією. Групи нервових клітин з однаковим функціональним значенням утворюють ядра сірої речовини.
За морфологічними ознаками локалізації, участю в передачі нер-вового імпульсу в складі сірої речовини спинного мозку можна виді-лити такі види нейронів: корінцеві — нейрити цих клітин виходять за межі спинного мозку в складі його вентральних корінців; внутрішні-клітини-нейрити останніх утворюють синапси на клітинах сірої речо-вини спинного мозку; пучкові нейрони — їх нейрити формують у білій речовині особливі пучки, які проводять нервові імпульси від певних ядер спинного мозку в його інші сегменти, або у відповідні відділи го-ловного мозку, утворюючи провідні шляхи центральної нервової систе-ми. В сірій речовині виділяють ділянки, які відрізняються нейтальним складом, а також характером волокон і нейроглії. Так, в дорсальних ро-гах сірої речовини виділяють губчастий шар, желатинозну речовину, власне ядро дорсального рога, його дорсальне ядро, або ядро Кларка. Губчастий шар дорсального рога містить дрібні пучкові нейрони, які занурені в широкопетлистий гліальний остов. Желатинозна речови-на утворена переважно елементами глії, у якій містяться, в незначній кількості, дрібні пучкові клітини. В складі дорсального рога розміщене власне ядро дорсального рога (грудне ядро) і значна кількість дифуз-но розсіяних дрібних мультиполярних вставних нейронів. Власне ядро дорсального рога містить пучкові клітини, аксони яких через передню білу спайку переходять на протилежний бік спинного мозку в боковий канатик білої речовини, де вони утворюють вентральний спинномо-зочковий та спинноталамічний шлях.
Дорсальні роги містять значну кількість дрібних мультиполярних асоціативних і комісуральних нейронів, нейрити яких утворюють си-напси на клітинах сірої речовини спинного мозку свого боку (асоці-ативні), або протилежного (комісуальні). Грудне ядро (ядро Кларка) утворене великими нейроцитами, а їх аксони входять до складу боко-вого канатика білої речовини свого боку і в складі дорсального спин-номозочкового шляху (Флексига) входять в мозочок. Нервові клітини проміжної зони сірої речовини утворюють два ядра: медіальне проміж-не, нейрити якого приєднуються до волокон вентрального спинномо-зочкового шляху (Говерса) свого боку, і латеральне проміжне, в якому
Розділ 4
Спеціальна гістологія
знаходяться асоціативні клітини симпатичної частини нервової систе-ми. Аксони цих клітин через вентральні спинномозкові корінці зали-шають спинний мозок і формують білі сполучні гілки симпатичного стовбура. Ядра вентральних рогів сірої речовини утворені найбільши-ми нейронами спинного мозку (100-150 мкм в діаметрі). їх нейрити утворюють основну масу нервових волокон вентральних корінців.
Через змішані спинномозкові центри вони входять у скелетні м'язи і закінчуються в моторних нервових закінченнях. У вентральних рогах сірої речовини спинного мозку розрізняють дві групи мотонейронів: медіальну, що іннервує м'язи тулуба і латеральну, характерну для ді-лянки шийного і поперекового потовщення спинного мозку. Латераль-не ядро вентральних рогів містить нейроцити, що іннервують м'язи кінцівок.
У сірій речовині розсіяні нервові клітини. Аксони їх у білій речовині діляться на довшу висхідну і коротшу низхідну гілочки. Ці волокна утворюють власні (основні) пучки білої речовини (їх у спинному мозку три пари), які прилягають безпосередньо до сірої речовини, утворюючи багато колатералей, що закінчуються синап-сами на моторних нейронах передніх рогів 4-6 суміжних сегментів спинного мозку.
Біла речовина спинного мозку складається із мієлінових нерво-вих волокон та опорного нейрогліального остова. Нервові волокна в білій речовині утворюють провідні шляхи (комплекси волокон) — ділянки певних рефлекторних дуг. До складу провідних шляхів слід віднести: власні шляхи рефлекторного апарату спинного мозку; з'єднуючі шляхи центрів спинного і головного мозку; висхідні (афе-рентні); низхідні (еферентні). Ті провідні шляхи, що не виходять за межі спинного мозку і служать для з'єднання різних його сегментів, називають короткими, а ті, що з'єднують окремі сегменти спинного мозку з ядрами головного — довгими. Короткі провідні шляхи бе-руть участь в утворенні двонейронних рефлекторних дуг. їх чутливі нейрони являють собою псевдоуніполярні нейроцити, розміщені в спинальних гангліях дорсальних спинномозкових корінців. Усі не-рвові волокна провідних шляхів мають однакову будову, їх можна розрізнити лише шляхом застосування біологічного експерименту. Центральний канал спинного мозку вкритий епендимою, клітини якої у зародка мають війки.
14-8-305
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
Головний мозок
Головний мозок складається із сірої та білої речовини. Сіра речо-вина розміщується на поверхні великих півкуль і утворює кору велико-го мозку та кору мозочка. Крім того, вона входить до складу численних ядер стовбурової частини мозку. У головному мозку зосереджуються центри вищої нервової діяльності, а також інших основних життєвих функцій організму. Стовбурову частину мозку становлять довгастий мозок, Вароліїв міст, середній, проміжний мозок та базальна части-на кінцевого мозку. Стовбур має внутрішній апарат, який забезпечує зв'язок між його основними частинами. До його складу входять ядра, які перемикають нервові імпульси, висхідні до кори півкуль і мозочка, а також низхідні від кори на стовбур і далі в певні сегменти спинного мозку. Мультиполярні нейрони ядер сірої речовини стовбура мозку за функціональною характеристикою представлені трьома групами: мо-торними, чутливими та асоціативними. Моторні нейрони аналогічні клітинам вентральних рогів спинного мозку, вони формують рухливі і змішані ядра черепномозкових нервів. Чутливі ядра стовбура анало-гічні нейронам дорсальних рогів спинного мозку, на них закінчуються аксони клітин спинальних гангліїв, які проходять в складі дорсального канатика спинного мозку, або чутливих вузлів голови (півмісяцевого, колінчастого, каменистого, яремного й верхнього пучкового). Велика кількість ядер забезпечує перемикання нервових імпульсів від спин-ного мозку та стовбура на кору і від кори на апарат стовбура й спинно-го мозку (зорові горби, зубчасте ядро, оливи та ін.).
Довгастий мозок містить аферентні та еферентні ядра власного апарата черепномозкових нервів, розміщених, переважно, в ділянці дна четвертого мозкового шлуночка; рухові (еферентні) ядра-меді-альніше, а чутливі (аферентні) — займають периферійну частину дна шлуночка (ядра тонкого та клиноподібного пучків, (Голля і Бурдаха) та ін.). У довгастому мозку локалізуються ядра, що перемикають не-рвові імпульси на інші відділи головного мозку, до них відносять ниж-ні оливи. Вони містять великі мультиполярні нейрони, а їх нейрити утворюють синапси на клітинах мозочка та зорового горба. В нижні оливи вступають нервові волокна від мозочка, червоного ядра, ретику-лярної формації та спинного мозку.
Важливим координуючим апаратом головного мозку еретикуляр-на формація, яка розміщується в центрі довгастого мозку, а в сплетен-
Розділ 4
Спеціальна гістологія
нях нервових волокон містяться невеликі групи мультиполярних не-йроцитів. Ретикулярна формація поширюється по центральній части-ні стовбура до довгастого мозку і є складним рефлекторним центром. Нейрони ретикулярної формації вмикаються у рефлекторні дуги як асоціативні ланцюги, які впливають на центри великих півкуль і мо-зочка, гальмуючи, а також на зворотному шляху одержують вплив від цих органів. Одним із прикладів механізму такого впливу є участь ре-тикулярної формації у сновидіннях.
Біла речовина довгастого мозку розміщується, переважно, вен-трально. Основними пучками нервових волокон є кортикоспиналь-ні-піраміди довгастого мозку, що лежать у його вентральній частині. Спинномозочкові шляхи утворюють вірьовчасті тіла, а відростки не-йронів ядер клиноподібного та тонкого пучків, які спрямовуються у мозочок, перехрещуються по середній лінії, утворюючи шов, а звідти направляються до зорового горба.
Вароліїв міст складається з великої кількості поперечно-розміще-них нервових волокон, між якими є нейроцити різних розмірів.
Середній мозок складається із сірої речовини чотиригорбикового тіла, покришки та ніжок великого мозку-великої кількості мієлінових нервових волокон, що йдуть від кори великих півкуль. У центральній частині покришки, її сірої речовини, розміщуються великі мультипо-лярні та дрібні веретеноподібні нейрони, а також нервові волокна.
Проміжний мозок. Основну його масу становить зоровий горб. Вен-трально від нього розміщується гіпоталамічна (підгорбикова) ділянка. Зоровий горб містить ядра, розмежовані одне від одного білою речо-виною. У вентральних ядрах таламічної ділянки закінчуються висхід-ні аферентні шляхи, від яких нервові імпульси передаються до кОри великих півкуль. Гіпоталамічна ділянка — це основний вегетативний центр головного мозку, який регулює температуру тіла, кров'яний тиск, водний і жировий обмін.
Мозочок — це частина головного мозку, яка рефлекторно регу-лює координацію руху та рівновагу тіла, а також стан м'язового то-нусу. Його аферентні та еферентні (або ефекторні) провідні шляхи з'єднують півкулі мозочка з варолієвим мостом, довгастим мозком та чотиригорбиковим тілом.
Сіра речовина утворює кору мозочка і ядра, що локалізуються у бі-лій речовині органу. Нейрони мозочка розміщуються шарами: зовніш-
14*
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
ній — молекулярний; середній — гангліозний і внутрішній — зернистий (рис 85, 86).

Рис. 85. Схема будови мозочка:
1-грушоподібні клітини; 2-зірчасті клітини; 3-кошикові клітини; 4-клітини-зерна; 5-великі клітини-зерна (клітини Гольджі); 6-дендрити грушоподібних клітин; 7-повзучі волокна; 8-мохоподібні волокна; 9-кошик волокон; 10-клу-бочки мозочка; 11-біла речовина мозочка; 12-горизонтальні клітини (за Кла-ром).
Молекулярний шар кори мозочка містить дрібні нейрони (корзин-часті та зірчасті), які передають гальмівні імпульси на клітини ганглі-озного шару. Кошикові нейрони мають невеликі розміри перикаріонів (10-25 мкм). їх численні тонкі дендрити, спрямовані до поверхні мо-зочка — аксони розміщуються горизонтально поперек звивин, утворю-ючи ряд бокових відгалужень, які над тілами грушоподібних нейронів нижнього шару формують синаптичний апарат, обплітаючи їх перика-ріони у вигляді «кошиків«.
Численні зірчасті нейрони молекулярного шару мозочка представ-лені двома видами великих і дрібних клітин. Дрібні нейрони мають короткі дендрити, а аксони утворюють синапси на дендритах грушо-подібних клітин. Великі зірчасті нейрони мають довгі розгалужені не-йрити та дендрити. Термінальні гілочки дендритів зірчастих нейронів контактують як з дендритами, так і з тілами грушоподібних клітин,
Розділ 4
Спеціальна гістологія
входячи до складу їх перицелюлярних синаптичних апаратів (кор-зинок). Еферентні (ефекторні) нейрони кори мозочка — це клітини (Пуркін'є) грушоподібної форми, які розміщуються в один ряд і утво-рюють гангліозний шар мозочка. Від перикаріонів цих клітин у моле-кулярний шар відходить 2-3 дендрити, які розгалужуючись в площині впоперек звивин, проходять через всю товщу молекулярного шару. Від протилежного полюса клітин Пуркін'є відходять нейрити, які в складі мієлінових волокон білої речовини спрямовуються до ядер мозочка. В межах зернистого шару нейрити грушоподібних клітин віддають кола-тералі, які, повертаючись у гангліозний шар, вступають в синаптичні зв'язки з сусідніми клітинами Пуркін'є.
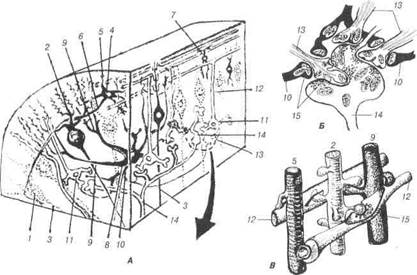
Рис. 86. Схема синаптичних зв'язків нейронів у корі мозочка:
А — звивини мозочка (зазначені пунктиром на поздовжньому зрізі рисунків Б і В); Б — клубочок зернистого шару; В — синапси в молекулярному шарі (у чорний колір зафарбовані термінальні нейрони; за Сантогатай): 1-грушопо-дібна клітина (клітина Пуркіньє); 2-дендрити грушоподібних клітин; 3-не-йрит грушоподібних клітин; 4-кошикові клітини; 5-дендрити; 6-аксон коши-кової клітини; 7- зірчаста клітина; 8-кошикові клітини-зерна (клітини Голь-джі); 9-дендрити клітин Гольджі; 11-клітини-зерна; 12-аксон клітин-зерен; 13-дендрити клітин-зерен; 14-мохоподібні волокна; 15-синаптичні пухирці.
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
Зернистий шар складається з великої кількості нейронів — зерен і двох видів клітин Гольджі (з короткими й довгими нейритами). Кліти-ни — зерна малі за об'ємом, містять бідний цитоплазмою перикаріон з великим круглим ядром. Характерні для них 3-4 коротких дендри-ти закінчуються розгалуженням у вигляді пташиної лапки, які утво-рюють синапси на закінченнях збуджуючих мохоподібних волокон, що є в корі мозочка (це зайшлі волокна). Нейрити клітин — зерен в молекулярному шарі кори Т-подібного діляться на дві гілки, які роз-міщуються вздовж звивин паралельно поверхні. Ці нейрити дуже до-вгі і перетинають зони розгалуження кількох рядів клітин Пуркіньє, утворюючи на них, а також на дендритах кошикових і грушоподібних клітин численні синапси, які розміщуються в молекулярному шарі. Нейрити клітин-зерен проводять збудження, яке до них надходить від мохоподібних волокон і передають його по паралельних волокнах (розгалуженнях своїх дендритів) на значну відстань до грушоподібних клітин (Пуркіньє), утворюючи синапси на їх дендритах.
Зірчасті клітини Гольджі є двох видів: з короткими й довгими нейри-тами. 3 короткими відростками клітини лежать ближче до гангліоз-ного шару, а їх дендрити в молекулярному шарі утворюють синапси з паралельними волокнами (з аксонами клітин — зерен). Нейрити ж цих клітин у зернистому шарі утворюють гальмівні синапси на кінцевих відділах дендритів клітин — зерен вище від синапсів мохоподібних во-локон, блокуючи надходження збуджуючих імпульсів з мохоподібних волокон на клітини — зерна. Клітини Гольджі з довгими відростками характеризуються розгалуженими дендритами й нейритами в зернис-тому шарі, які направляються у білу речовину мозочка.
В кору мозочка входять два види аферентних волокон — мохопо-дібні та повзучі.

