




Пирамидная система представляет собой совокупность двигательных центров коры мозга, моторных центров черепных нервов, залегающих в стволе мозга, и моторных центров в передних рогах спинного мозга, а также эфферентных проекционных нервных волокон, связывающих их между собой. Пирамидные пути обеспечивают проведение импульсов в процессе сознательной регуляции движений.
Пирамидные пути формируются от гигантских пирамидных нейронов (клеток Беца), а также крупных пирамидных нейронов, локализованных в V слое коры больших полушарий. Примерно 40% волокон начинается от пирамидных нейронов в предцентральной извилине, где находится корковый центр двигательного анализатора; около 20% волокон — от постцентральной извилины, а остальные 40% волокон — от задних участков верхней и средней лобных извилин, и от надкраевой извилины нижней теменной дольки, в которой расположен центр праксии, контролирующий сложные координированные целенаправленные движения. Число нервных волокон в пирамидных путях на одной стороне тела составляет примерно 1 млн.
Пирамидные пути подразделяют на корково-спинномозговой и корково-ядерный. Их общей особенностью является то, что они, начинаясь в коре правого и левого полушарий, переходят на противоположную сторону мозга (т. е. перекрещиваются) и в конечном итоге осуществляют регуляцию движений контрлатеральной половины тела.

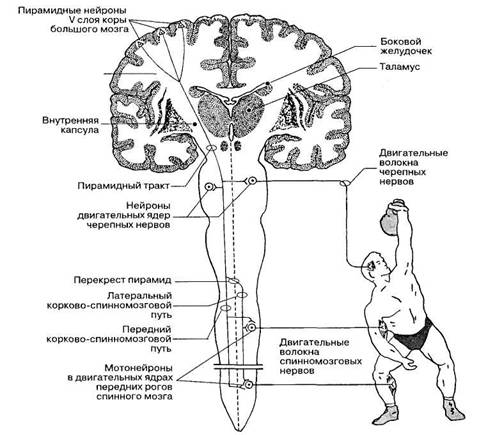
Рис. 102. Проводящий путь пирамидной системы.
Корково-спинномозговой путь, как уже отмечалось, начинается от новой коры больших полушарий (рис. 102). Его волокна проходят в задней ножке внутренней капсулы и продолжаются в основании среднего мозга (средняя часть ножки мозга) и основании моста. В продолговатом мозге на уровне пирамид волокна медиальных частей правого и левого корково-спинномоз-говых путей переходят на противоположную сторону, образуя перекрест пирамид (см. рис. 97). Здесь перекрещивается около 80% волокон пирамидных трактов. Перекрещенные волокна спускаются в спинной мозг и идут в составе боковых канатиков в виде латерального корково-спинномозгового тракта. Волокна пирамидных трактов заканчиваются синапсами на мотонейронах двигательных ядер передних рогов спинного мозга.
Неперекрещенная часть волокон продолжается в спинной мозг в составе его передних канатиков под названием переднего корково-спинномозгового тракта. Волокна этих трактов переходят на противоположную сторону на уровне соответствующих сегментов спинного мозга и заканчиваются синапсами преимущественно на интернейронах (IV—VI пластин Рикседа), которые
 передают центральные нервные влияния на мотонейроны двигательных ядер передних рогов спинного мозга. Только небольшая часть пирамидных волокон (около 8%) непосредственно оканчивается на больших мотонейронах спинного мозга. Бoльшая часть волокон пирамидных путей оканчивается на интернейронах спинного мозга, а уже от них раздражение передается на мотонейроны.
передают центральные нервные влияния на мотонейроны двигательных ядер передних рогов спинного мозга. Только небольшая часть пирамидных волокон (около 8%) непосредственно оканчивается на больших мотонейронах спинного мозга. Бoльшая часть волокон пирамидных путей оканчивается на интернейронах спинного мозга, а уже от них раздражение передается на мотонейроны.
Нервные волокна, образующие пирамидные пути, неоднородны как по своим структурным, так и функциональным параметрам. Б льшую часть составляют тонкие и безмиелиновые нервные волокна, а крупные миелиновые волокна, отличающиеся высокой скоростью проведения нервного импульса, составляют значительно меньшую часть. С этими структурными особенностями связано наличие быстро- и медленнопроводящих частей пирамидной системы.
Б льшая часть волокон пирамидной системы (примерно 55%) заканчивается в шейных сегментах спинного мозга, 20% — в грудных и 25% — в пояс-нично-крестцовых. При этом волокна переднего корково-спинномозгового тракта не опускаются ниже грудных сегментов.
Полагают, что по пирамидным путям происходит активация преимущественно флексорной мускулатуры (сгибателей) и торможение экстензорной (разгибателей). Возможно, что одно из функциональных значений пирамидной системы состоит в тонизирующем воздействии корковых центров на работу мотонейронов спинного мозга.
Корково-ядерный путь начинается там же, где и корково-спинномозговой, а именно на пирамидных нейронах V слоя коры, но только в нижней трети предцентральной извилины. Их волокна проходят в колене внутренней капсулы между ее передней и задней ножками и продолжаются в основании ножки мозга. В стволе мозга большая часть волокон корково-ядерного пути переходит на контрлатеральную сторону и, достигая двигательных ядер черепных нервов, заканчивается в них синапсами на мотонейронах.

