




1.6.1. Ядро
У еукаріотичних клітинах ядро має дві мембрани—внутрішню і зовнішню. Між мембранами — перинуклеолярний простір. Ядерні мембрани пронизують пори, через які здійснюється обмін між цитоплазмою і внутрішньою частиною ядра—нуклеоплазмою. Через пори в ядро проникають різні ферменти, що забезпечують процеси реплікації і транскрипції, білки, стероїдні гормони; транспортуються зрілі мРНК, тРНК, рРНК, зібрані субодиниці рибосом і т. д. (рис. 3).
У нуклеоплазмі інтерфазного (неподільного) ядра знаходиться молекула ДНК не у вільному стані, а у виді хроматину—комплексуДНК із п'ятьма основними білками — гістонами (репресорами) і кислими фосфорильованими регуляторними білками.
Структурною одиницею хроматину є нуклеосома (рис. 4). Ядро нук-леосоми створене чотирма типами гістонів, що утворюють октамер. Октамер взаємодіє з подвійною спіраллю ДНК. Лише частина генетич
ної інформації, записаної в ДНК-хроматину, реалізується у вигляді копій ДНК. Молекули ДНК, зв'язані з нуклеосомами, далі згортаються у над-спіраль. Перед поділом клітини хроматин знаходиться в найбільш конденсованому стані й утворює хромосоми.


А
500 нм
Б
500 нм
Рис. 3. Ядро клітини:
А. Тонкий зріз клітини дріжджів ШіоскЛогиІа ^игіпв; фіксація за допомогою КМп04. Видно рельєфні ядра (Я) с декількома порами (П) і мітохондрії (М). Б. Препарати клітин 8асс1іаготусе8 сегеувіае, отримані методом замороження — травлення. Видно ядра (Я) із численними великими порами і декілька мітохондрій (М)
Генетична функція ДНК була вперше доведена в експериментах Евері. Усі соматичні диплоїдні клітини організму даного виду містять ту
саму кількість ДНК, незалежно від живлення й умов зовнішнього середовища. Нуклеотидний склад ДНК різний у різних видів, але специфічний для організмів даного виду. В усіх ДНК кількість залишків аденіну дорівнює кількості залишків тиміну, а кількість залишків гуаніну дорівнює кількості залишків цитозину.
На підставі даних рентгенострук-турного аналізу волокон ДНК, а також закону «еквімолярності» основ (Чаргафф, 1949—1953) Уотсон і Лемент (1953) висловили припущення, що молекула ДНК складається з двох антипаралельних ланцюгів, що утворюють подвійну спіраль, у якій за рахунок водневих зв'язків утворюють-Рис. 4. Будова нуклеосоми ся пари основ Аденін — Тимін,

Гуанін — Цитозін. Пари основ вкладені щільною «стопкою» вздовж осі молекули ДНК на відстані 3,4 А° одна від одної.
Подвійна спіраль побудована зі строго комплементарних ланцюгів, і ця її здатність дозволяє зрозуміти механізм, що забезпечує точну реплікацію молекул ДНК.
Гіпотеза Уотсона — Кріка дозволила сформулювати загальний принцип, названий Кріком «центральним постулатом молекулярної генетики»: потік генетичної інформації спрямований від ДНК через РНК до білка (ДНК —► РНК —► білок). Відповідно до центрального постулату, є три основні процеси, що беруть участь у збереженні і передачі генетичної інформації: реплікація—копіювання ДНК з утворенням ідентичних дочірніх молекул; транскрипція — процес, у результаті якого генетична інформація, поміщена в ДНК, переписується на РНК із наступним переносом РНК до рибосом; трансляція—процес, у результаті якого генетична інформація переводиться на «мову» білкової структури, у якому використовується двадцятибуквенний алфавіт амінокислот.
Склад нуклеоплазми. Нуклеоплазма — це рідка фаза ядра. У нуклео-плазмі локалізовані ферменти гліколізу, є всі амінокислоти, вільні нук-леотиди, ферменти реплікації ДНК.
Функції ядра:
1. Ядро є хранителем генетичної інформації у вигляді нуклеотидної послідовності молекул ДНК.
2. У ядрі відбуваються такі етапи синтезу білка:
— реплікація (точне відтворення) ДНК напівконсервативним способом, так як лише один з ланцюгів дочірньої ДНК синтезується заново;
— транскрипція — копіювання інформації з ділянок ДНК на матричну РНК, синтез транспортних і рибосомальних РНК на матриці
ДНК;
— дозрівання мРНК, тРНК та іРНК;
— складання субодиниць рибосоми;
— репарація — видалення за допомогою комплексу ферментів помилок ланцюга ДНК.
3. Участь у процесах поділу клітини. Поділ клітини починається з поділу ядра, що містить ДНК, кількість якого постійна для всіх клітин даного виду, і в яких міститься однакова кількість хромосом.
4. Транспортна функція.
1.6.2. Ядерце
У клітинах, де відбувається активний синтез білка, у ядрі спостерігається декілька ядерець. У ядерці розрізняють фібрілярні структури —
нуклеолонема (нитки гетерохроматину — більш спіралізована молекула ДНК) і гранулярні структури—ґранули РНП фибонуклеопротещів). Аморфна речовина, що заповнює простір між гранулами і фібрилярни-ми структурами, містить: 3—5 \% РНК; білки-фосфопротеїди; ферменти: кислу фосфотазу, нуклеозидфосфорилазу, що бере участь у синтезі нуклеотидів і коферментів; ферменти, які каталізують утворення мРНК. Функції ядерця:
1. Синтез компонентів, що йдуть на складання рибосоми.
2. Синтез рибосомної і матричної РНК.
3. Збирання субодиниць рибосоми.
1.6.3. Ендоплазматичний ретикулум
У світловому мікроскопі ендоплазматичний ретикулум («ендо» — усередині, «ретикулум» — мережа) виглядає як мережа (рис. 5).
Мембрани ендоплазматичного ретикулуму складаються в основному з ліпопротеїдів.
У клітині енцогшазматичний ретику-лум складається із систем, обмежених мембраною, каналів, що являють собою сплощені «цистерни», трубочки, які з'єднуються одна з одною. При гомогенізації тканини ендоплазматичний ретику-лум руйнується, мембрани замикаються, утворюючи дрібні пухирці, — макросоми.
До поверхні ендоплазматичного ре-тикулуму можуть бути прикріплені рибосоми. У цьому випадку ендоплазматичний ретикулум називають шорсткуватим.
На поверхні мембран шорсткуватого ендоплазматичного ретикулуму розташовані білки-рецептори для рибосом, фун-
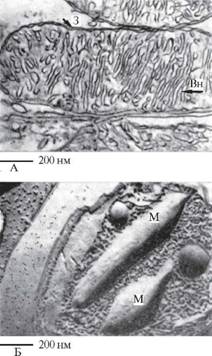
Рис. 5. Ендоплазматичний ретикулум:
А. Ультратонкий зріз через канальці і цистерни шорсткуватого ендоплазматичного ретукулуму, усіяного рибосомами; ацинарні клітини підшлункової залози летючої миші. Б. Препарати клітин ХУіскегіктіа піюгезсеш, отримані методом заморожування — травлення. Видна зовнішня поверхня (З) ендоплазматичного ретукулуму, покрита частинками розміром 5 нм, і внутрішня поверхня (Вн) з невеликою кількістю частинок розміром 10 нм.

-;- 100 нм
А

50 нм
кція яких полягає у зв'язуванні рибосом з ендоплазматичним ретику-лумом.
Мембрани ендоплазматичного ретикулуму, що не мають рибосом, називають гладкими.
Шорсткуватий ендоплазматичний ретикулум складається з великої кількості цистерн, розташованих паралельно одна одній і зв'язаних між собою так, що в клітині утворюється система комунікацій, по якій синтезовані на мембранах ендоплазматичного ретикулуму білки можуть транспортуватися або в інші частини клітини, або назовні.
1. Після завершення білкового синтезу поліпептидний ланцюг виходить з рибосоми через пори, утворені рецепторним білком, попадає в цистерни ендоплазматичного ретикулуму. Там самовільно формуються II і III структури білка. Рішення про те, залишиться заново синтезований білок у цитоплазмі чи потрапить у систему ЕР — Гольджі, приймається в процесі синтезу білка (трансляції) у залежності від присутності чи відсутності в складі цього білка «сигнальної» послідовності амінокислот.
2. Мембрани гладкого ендоплазматичного ретикулуму також беруть участь у синтезі різних речовин: у них протікає синтез гормонів-сте-роїдів, тригліциридів; метаболізм глікогену; цикл глюкуронової кислоти, що пов'язаний із розщепленням токсинів.
3. У мембрані гладкого ендоплазматичного ретикулуму, наприклад, у клітинах печінки існує система транспорту електронів, необхідна для окиснювання чужорідних речовин. При перебігу процесів детоксикації площа мембрани гладкого ендоплазматичного ретикулуму збільшується в десятки разів. У процесі детоксикації чужорідні речовини, як правило, погано розчинні у воді, окисляються до водорозчинних продуктів, що легко виводяться з організму. Ця система містить ферменти-цито-хроми В5, Р45 і флавопротеїни.
4. Однією з функцій гладкого ендоплазматичного ретикулуму м'язової клітини саркоплазматичного ретикулуму, є поглинання іонів кальцію, що викидаються в цитоплазму (саркоплазму) при скороченні м'язового волокна у відповідь на подразнення.
Слід зазначити, что гладкий ендоплазматичний ретикулум розвинутий в тих клітинах, що транспортують іони (наприклад, у залозистих клітинах слизової оболонки шлунка).
1.6.4. Комплекс Гольджі
«Купка» Гольджі складається зі сплощених, обмежених мембраною, структур-цистерн, з якими зв'язані різного роду пухирці («купка млинців») (рис. 6).
Кількість апаратів Гольджі на клітину залежить від ступеня диференціювання. Пухирці формуються на цистернах, відбруньковуючись від них. Через те, що цистерни мають вигнуту форму (чаша), то розрізняють дві поверхні апарату Гольджі: 1) увігнуту (внутрішньо-секретую-чий полюс), повернену до цитоплазми; 2) випуклу поверхню (формуючий полюс), зверненудо ендоплазматичного ретикулуму і ядра.
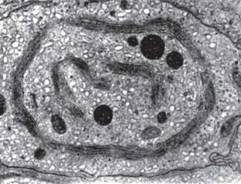
Рис. 6. Комплекс Гольджі в клітині епідерміса кролика
Функції комплексу Гольджі:
1. На формуючому полюсі відбувається поглинання білків і ліпідів, синтезованих у шорсткуватому і гладкому ендоплазматичному ретику-лумі і функціонально зв'язаних із пухирцями апарату Гольджі.
2. Численні дослідження різних клітин травної системи показали, що апарат Гольджі бере активну участь у включенні вуглеводних компонентів у білки з утворенням різних важливих глікопротеїдів (тирео-глобулін, імуноглобулін, муциген), полісахарид-білкових комплексів, необхідних для створення нових клітинних структур. Включення термінальних вуглеводів у білкову молекулу є пусковим моментом для процесу екзоцитозу продуктів секреції.
3. В апараті Гольджі накопичуються вуглеводи у виді розчинних олі-гомерів і уронідів, полімери яких здатні сильно набухати. Вони входять до складу слизу, що виділяється апаратом Гольджі.
4. Апарат Гольджі бере участь у завершенні формування ліпопро-теїнів шляхом обволікання ліпіду мембраною і доставкою їх у певні місця плазматичної мембрани.
5. Є дані про те, що ферменти апарату Гольджі беруть активну участь у сульфатируванні продуктів секреції (наприклад, утворення мукополі-сахаридів епітеліальних клітин ШКТ).
6. Апарат Гольджі бере участь у процесах секреції молока (включення галактози в глікопротеїди, накопичення лактози).
7. На думкудеяких дослідників, апарату Гольджі належить важлива роль у синтезі і секреції вуглеводмістких компонентів міжклітинного матриксу, а також у процесах перетворення в упорядковані структури таких структурних білків, як колаген.
1.6.5. Мітохондрії
Мітохондрії — субклітинні органели завдовжки 2 мкм і діаметром 0,5 мкм (рис. 7).
Мітохондрії часто знаходяться в клітині поблизу від структур, що потребують АТФ, чи від джерел клітинного «палива». За розмірами вони подібні з бактеріями, однак, форма їх варіює. Мітохондрії мають дві мембрани, що можуть бути відділені одна від одної. Зовнішня мембрана містить моноамінооксидазу і ферменти, що активують жирні кислоти; вона вільно проникна для більшості розчинних низькомолекулярних сполук. Внутрішня мембрана має значно більшу поверхню, створювану за рахунок наростів, називаних кристами. Вона непроникна для іонів №+, їуі£2+, СІ-, більшості амінокислот. На поверхні внутрішньої мембрани розташовуються окремі групи ферментів ланцюга перенесення електронів, відомі за назвою дихальних ансамблів. Крім того, із внутрішньою поверхнею внутрішньої мембрани пов'язані структурні одиниці, які містять молекули ферментів, що беруть участь у синтезі АТФ (мітохондріальна АТФаза). Внутрішня мембрана містить декілька ферментів, що
Рис. 7. Мітохондрії:
А. Ультратонкий зріз мітохондрії із ниркової клітини миші; З — зовнішня міто-хондріальна мембрана; Вн — внутрішня мітохондріальна мембрана. Б. Препарат клітин 8асс1іаготусос1е8 ludwigii, одержаний методом заморожування — травлення. Видно дві мітохондрії (М) із оголеними поверхнями. У лівій частині рисунка видно частини типової плазматичної мембрани з жолобками
здійснюють перенесення специфічних метаболітів через мембрану. Внутрішній матрикс мітохондрій містить велику кількість білка, частину якого складають різні ферменти циклу Кребса, р-окислення жирних кислот, циклу сечовини, а також пул (депо) АДФ, АТФ, НАД, НАДН і КоА, відділений від їх цитоплазматичного пулу внутрішньою мембраною, непроникною для цих сполук. У матриксі знаходяться великі гранули, що характеризуються високою електронною щільністю. Удеяких мітохондріях були виявлені рибосоми. Мітохондрії містять молекули ДНК. У мітохондріальній ДНК кільцева структура (як у бактерій), що несе інформацію для синтезу певних поліпептидних ланцюгів цито-хромів і АТФази. Таким чином, у мітохондріях синтезуються незвичайно гідрофобні водонерозчинні білки, і вдається уникнути необхідності їх пересування на будь-яку відстань через цитоплазму. Інформація із синтезу багатьох інших поліпептидних ланцюгів закладена, як правило, у ДНК ядра.
Функції мітохондрій:
1. Мітохондрії — енергетичні станції клітини, у яких синтезується АТФ — основне джерело енергії в живому організмі. Мітохондрії існують у всіх еукаріотичних клітинах, крім зрілих еритроцитів.
2. У мітохондріях здійснюється процес клітинного дихання — споживається кисень і виділяється СО2. У цих органелах мембранні білки містять як коферменти (небілкова частина ферменту) вітаміни, неорганічні сполуки: залізо, сірку, мідь.
3. У мітохондріях відбувається перетворення енергії, запасеної в їжі, в інші види енергії.
Клітини одержують необхідну енергію за рахунок окиснення білків, жирів, вуглеводів, які гідролізуються за межами мітохондрії до амінокислот, моносахаридів, гліцерину і жирних кислот. Ці низькомолекулярні продукти в результаті послідовних реакцій перетворюються в похідні оцтової кислоти—ацетил-КоА. У такому вигляді ацетильні групи окисляються до СО2 у циклі Кребса. Ферменти цього процесу знаходяться в матриксі мітохондрії. У результаті циклу Кребса відбувається утворення атомів водню, акцептором якого є НАД-кофермент. Далі електрони з цієї сполуки надходять у ланцюг переносу на внутрішній мембрані мітохондрій, де рухаються від переносника до переносника на молекулярний О2, що відновлюється до Н2О. Цей процес називається диханням, а ланцюг перенесення електронів — дихальним ланцюгом. Дихання сполучене із фосфорилуванням. У процесі перенесення електронів уздовж дихального ланцюга створюється градієнт концентрації
Н+ у напрямку, перпендикулярному поверхні мембрани (Мітчел, 1966), що використовується для синтезу АТФ. Таким чином, у мітохондріях відбуваються важливі процеси окисного фосфорилування.
Крім дихання, пов'язаного із фосфорилуванням, у мітохондріях існує дихання не пов'язане із накопиченням макроергів. Це так зване нефосфориловане окиснення. Енергія, що вивільнюється при цьому, розсіюється у вигляді тепла. Існують деякі сполуки, здатні розривати зв'язок між диханням і фосфорилуванням. Зокрема, до них належать динітрофенол, дикумарин, тироксин, олігоміцин та ін.
Мітохондрії можуть накопичувати іони Са2+ за рахунок енергії, що вивільняється в процесі перенесення електронів, але процес акумуляції Са2+ у мітохондріях альтернативний процесу окисного фосфорилування.
1.6.6. Лізосоми
Лізосоми — субклітинні структури діаметром від 0,5 до 2—3 мкм. Обмежені елементарною мембраною завтовшки 7 нм і містять гідролітичні ферменти (протеази, нуклеази, глікозідази, ліпази, фосфоліпази, фосфатази тощо). Лізосоми присутні, за окремим винятком (еритроцити ссавців), у всіх тваринних клітинах.
Лізосоми беруть участь у «перетравлюванні» матеріалів, що надходять ззовні, і структурних елементів клітини. У випадку загибелі клітини відбувається розрив лізосомальної мембрани і починається автоліз клітини: гідролази розщеплюють білки до амінокислот, нуклеїнові кислоти до нуклеотидів, полісахариди до моносахаридів.
У тканинах із високою швидкістю відновлення клітин вміст лізо-сом у клітинах підвищений (у клітинах епітелію ниркових канальців, остеокластів кістки, молочних залоз).
Обмежений елементарною мембраною пухирець із гідролітичними ферментами називається первинною лізосомою.
Коли первинна лізосома зливається з вакуолями, фагосомами, облямованими пухирцями, вона стає вторинною лізосомою. Вторинна лізосома здатна до фагоцитозу: захоплювати, наприклад, мітохондрію, яка дегенерує, і «перетравлювати» її (аутофагія), утворюючи аутолізо-сому. Лізосоми, що перетравлюють чужорідні речовини, називаються гетеролізосомами.
Лізосоми беруть участь у процесах екзоцитозу гідролаз.
У шорсткуватому ендоплазматичному ретикулумі синтезуються гідролітичні ферменти. Вони надходять в апарат Гольджі, де відбувається упакування ферменту в секреторних гранулах та злиття їх з первинною лізосомою. Утворюються зимогенні ґранули (щільні агрегати
молекули зимогену—неактивного ферменту, оточені мембраною). Зи-могенні гранули мігрують до поверхні клітини (мембрани) і секретиру-ються назовні.
Функції лізосом:
1) структура клітинного «перетравлювання», яке супроводжують піноцитоз і фагоцитоз;
2) участь у катаболізмі структурних компонентів самої клітини і речовин, що надходять ззовні;
3) участь у процесі фагоцитозу (лізосоми макрофагів відіграють роль у захисних механізмах);
4) екзоцитоз;
5) посмертний автоліз.

