




Одним из характерных проявлений жизни является удивительная способность живых организмов кинетически регулировать химические реакции, подавляя стремление к достижению термодинамического равновесия. Ферментативная кинетика занимается исследованием закономерностей влияния химической природы реагирующих веществ (ферментов, субстратов) и условий их взаимодействия (концентрация, рН среды, температуры, присутствие активаторов или ингибиторов) на скорость ферментативной реакции. Главной целью изучения кинетики ферментативных реакций является получение информации, которая может способствовать выяснению молекулярного механизма действия фермента.
Общие принципы кинетики химических реакций применимы и к ферментативным реакциям. Известно, что любая химическая реакция характеризуется константой термодинамического равновесия. Она выражает состояние химического равновесия, достигаемого системой, и обозначается Кр. Так, для реакции:
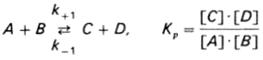
константа равновесия равна произведению концентраций образующихся веществ, деленному на произведение концентрации исходных веществ. Значение константы равновесия обычно находят из соотношения констант скоростей прямой (k +1) и обратной (k – 1) реакций, т.е. Кp = k +1/ k –1. В состоянии равновесия скорость прямой реакции: v +1 = k + 1[ А ] • [ B ] равна скорости обратной реакции: v –1 = k – 1 [ С ] • [ D ], т. е. v +1 = v –1 соответственно k +1[ А ]•[ B ] = k –1[ С ]•[ D ], или
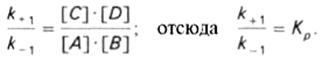
Изучая явление насыщения, Л. Михаэлис и М. Ментен разработали общую теорию ферментативной кинетики. Они исходили из предположения, что ферментативный процесс протекает в виде следующей химической реакции:

т.е. фермент Е вступает во взаимодействие с субстратом S с образованием промежуточного комплекса ES, который далее распадается на свободный фермент и продукт реакции Р. Математическая обработка на основе закона действующих масс дала возможность вывести уравнение, названное в честь авторов уравнением Михаэлиса–Ментен, выражающее количественное соотношение между концентрацией субстрата и скоростью ферментативной реакции:

где v – наблюдаемая скорость реакции при данной концентрации субстрата [ S ]; K S– константа диссоциации фермент-субстратного комплекса, моль/л; V max– максимальная скорость реакции при полном насыщении фермента субстратом.
Из уравнения Михаэлиса–Ментен следует, что при высокой концентрации субстрата и низком значении KSскорость реакции является максимальной, т.е. v = Vmax. При низкой концентрации субстрата, напротив, скорость реакции оказывается пропорциональной концентрации субстрата в каждый данный момент (реакция первого порядка).
Скорость любой ферментативной реакции непосредственно зависит от концентрации фермента (рис. 4.19). Существующая линейная зависимость между этими величинами, когда скорость реакции прямо пропорциональна количеству присутствующего фермента, справедлива только в определенных условиях, например в начальный период ферментативной реакции, так как в этот период практически не происходит обратной реакции, а концентрация продукта оказывается недостаточной для обратимости реакции. Именно в этом случае скорость реакции (точнее, начальная скорость реакции v) будет пропорциональна концентрации фермента.

Рис. 4.19. Зависимость скорости реакции от концентрации фермента в присутствии насыщающих концентраций субстрата.
Как было отмечено, фермент является одной из реагирующих молекул в химической реакции и при взаимодействии с субстратом образует промежуточный фермент-субстратный комплекс, который далее подвергается распаду на продукт и свободный фермент:

Если упростить это уравнение, исключив промежуточный ES-комплекс:

то в уравнениях для скоростей прямой и обратной реакций обязательным компонентом является концентрация фермента:

Однако в уравнениях для константы равновесия (Keqили Кр) концентрация фермента уже не имеет значения:

Как видно, константа равновесия (K р) ферментативной реакции не зависит от концентрации фермента. Определяя скорость и направление химической реакции, фермент тем не менее не оказывает влияния на конечные (равновесные) концентрации реагирующих молекул и продуктов, определяющих величину константы равновесия.
Ферментативные процессы являются регулируемыми.
1.Конкурентное ингибирование – сродство с активным центром.
2.Неконкурентное ингибирование – аллостерическое.
3.Антиконкурентное ингибирование – ингибитор соединяется с (es) комплексом.
4.Смешанное ингибирование – по активному и аллостерическому центру.
5.Ингибирование избытком субстрата.

