




Тагам көмірсулары корытылған кезде түзілген жэне ішектің қабырғасына сіңірілген Моносахаридтер қан агынымен бауырга жеткізіледі.Бауырга v. portae арқылы глюкозаныц исеткізілуі мен v. hepatica арқылы шығарылуы кезеңінде бауыр маңызды реттеуші кызмет атқарады да, қанда глюкозаныц қалыпты, физиологияльщ деңгейін сактауды қамтамасыз етеді, яғни глюкостатикалық қызмет атқарады.
Бауырдың глюкостатикалық қызметі бірқатар күрделі метаболитикалық процестермен, агап айтқанда, моносахаридтердің глюкозага өзара айналу реакциялары, метаболиттерден глюконеогенез процестері, бауырда глюкозаныц гликоген түрінде жинақталуы жэне гликоген мобилизациясы процестерімен қамтамасыз етіледі.
Жоғары сатылы жануарлардың бауырында жеңіл экстрагирленген (85%) жэне аль-буминмен байланыскан (15%) гликоген бар. Адамның бауырындағы гликогеннің мөлшері т^уліктік жэне маусымдық өзгерістерге байланысты эртүрлі болады. Бір тэулік ішінде бауырдағы гликогеннің жалпы мөлшерінің 70% -га жуыгы ыдырайды жэне қайта синтезделin отырады.
Ағзаның физиологияльщ қызметтерін қамтамасыз ету үшін гликогеннің синтезделу процесі (қорға жиналуы) жэне оныц ыдырауы (мобилизациясы) үлкен орын алады. Бауырда пшкоген негізінен глюкозадан синтезделеді. Қанда глюкозаныц деңгейі артқан кезде оныц бауырга түсуі артады жэне «Көмірсулар алмасуы мен қызметтері» деген тарауда көрсетілген механизмдер арқылы гликогеннің синтезі мен оныц бауырда қорға жиналуы күшейеді.
Гликоген синтезі АТФ-тың қатысуын қажет ететін энергияга тэуелді процесс. АТФ энергиясы гексокиназаның (фосфоглюкокиназаның) эсерінен глюкозо-6-монофосфаттың түзілуі үіпін пайдаланылады. Ары қарай фосфоглюкомутазаның қатысуымен глюкозо-1- монофосфат түзіледі. Одан УДФ - глюкозамен қатар гликоген синтезделеді.
Қанда глюкоза деңгейі төмендеген кезде, гликогеннің мобилизация (ыдырау) процесі б^лсендіріледі, ол фосфорилазаны белсендірудің аденилатциклазды каскадты механизмі арқылы іске қосылады. Фосфорилаза «а» эсерінен гликогеннің 1,4 - байланысының фосфоролизі жүреді де, канда глюкозаның деңгейі артады. Гликогеннің 1,6 - байланысының ыдырауы амило - 1,6 - гликозидаза эсерінен болады.
Бауыр арқылы реттелетін қандағы глюкозаның деңгейінің реттелуі ағзаның барлық тканьдерінің, эсіресе жүйке тканінің энергиямен қамтамасыз етілуіне эсер етеді. Гликогеннің мобилизациясы кезінде қанға келіп түсетін глюкозаның шамамен 70%- ын жүйке тканьдері пайдаланады. Бауырдз глюкоза катаболизмы негізінен гексозомонофосфатты жолмен (пен-тозды немесе апотомиялық жол) жүзеге асырылады. Соның салдарынан эртүрлі синтездер үшін пайдаланылатын бос энергия бөлінеді, детоксикация процестерін қамтамасыз ету үшін, холестерин синтезі үшін, май қышқылдары синтезі жэне т.б. үшін пайдаланылатын НАДФН2 -нің көп мөлшері түзіледі. Бүл жолда, сонымен қатар, нуклеотидтер мен нуклеин қышқылдарының синтезі үшін бауыр жэне басқа да тканьдер пайдаланатын пентозалар (рибоза) түзіледі. Бауырда глюкурон қышкылы мен мукополисахаридтер (гепарин) синтезі қарқынды жүреді.
Ішек қабырғасында үшацилглицеридтердің қорытылу өнімдері көбінесе (70-90 %) бейтарап майларға дейін ресинтезделеді, олар бауырды айналып өтіп, лимфатикалық жүйе бойымен тасымалданады. Қысқа кемірсутекті тізбегі бар май қышқылдарының бір бөлігі ішек қабырғасынан қанға түседі де, бауырға жеткізіледі. Сонымен қатар ішектен бауырға қан ағынымен липоидтар корытылуының өнімдері (холин, коламин жэне т.б.), сол сияқты ішек қабырғасында ресинтезделген фосфолипидтер, гликолипидтер, холестеридтер жеткізіледі.
Химиялық қүрамьіна байланысты фосфолипидтер глицеридтер (лецитиндер, кефалин-дер, серинфосфатидтер, плазмогендер,инозинфосфатидтер) жэне фосфолипидтер глицеридтер емес (сфингомиелиндер) болып бөлінеді. Бүл қосылыстардың барлығы да бауырда синтезделеді жэне белоктармен комплекс түрінде (липопротеидті комплекстер) қан ағынымен барлық тканьдерге жеткізіледі де, жаңа түзіліп келе жатқан клеткалардың клеткалық мембра-наларын құру үшін паЙдаланылады.Адамның бауырында шамамен 2,9 % фосфолипидтер бар. Холиннің, бетаинның жэне метиониннің қатысуымен олардың синтезі күшейе түседі. Сондықтан бұл қосылыстар липо-тропты факторлар деп аталады. Холин жетіспеушілігі кезінде лецитин синтезі төмендейді, бұл бауырда бейтарап майлар синтезінің күшеюіне жэне осы мүшенің майлы инфильтрация-сына экеледі. ХолиннЫН деңгейі, яғни коламиннен оның синтезі метильді топтың донаторы ретінде метионинның катысуына байланысты болады. Холинның мөлшері арткан кезде оның бір бөлігі бетаинге айналады.
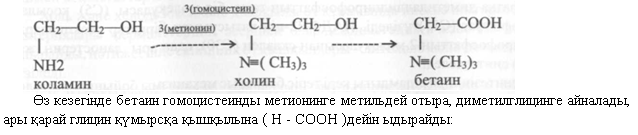

Қоршаған ортаның температурасы артқан сайын ағзаның холинге деген қажеттілігі артады. Бүл холинның бетаинге, одан ары глицинге айналуының күшеюіне байланысты. Бауырда холинның жетіспеушілігі туады, бұл фосфолипидтер синтезі жылдамдығының тежелуін жэне бауырдың майлы инфильтрациясын тудырады. Бауырда фосфолипидтер синтезінің жылдамдығы қанда холестерин деңгейі артқан кезде баяулайды, ал кальций ион-дары фосфолипидтердің синтезін күшейтеді.
Бауыр - холестерин синтезі қарқынды жүретін негізгі мүше болып табылады (80 %). Бүдан аз мөлшерде холестерин бүйрек үсті бездерінде, аталық бездерде, аналық бездерде, теріде, тері қабаттарында синтезделеді. Холестерин синтезі үшін қажетті ферменттер жетілген эритроциттерден басқа барлық клеткаларда болады.
Бір тэулік ішінде бауырда 1,5-4,0 г холестерин синтезделеді (орташа 2,0 гр). Тағаммен бірге бір тэулік ішінде 0,4-0,5 грамм холестерин түседі. 70 кг салмағы бар адам ағзасында холестериннің жалпы мөлшері 105-175 г немесе дене салмағының 0,2 %. Бүл жағдайдаоның ең көп мөлшері бүйрек үсті безінде (4,5-10%) жэне мида (2%) болады. Бауырда холестерин шамамен 0,3%, оның ішінде, бос холестериннің (этерифицирленбеген) үлесі 80% жэне эте-рифицирленген холестериннің үлесі 20%.
Бауырда холестерин синтезінің жеделдеуіне тағаммен түсетін экзогенді холестерин мөлшері эсер етеді. Тағамда холестерин деңгейі төмендеген кезде, бауырда холестерин синтезі артады жэне керісінше. Бауырда холестерин синтезінің үдеуі майларды көп мөлшерде пайдаланған кезде байқалады, бүл жерде майлар ацетил КоА-ның қайнар көзі болып табылады, олар кетон денелері мен май қышқылдарының ғана емес, сонымен қатар холестериннің де синтезінің қайнар көзі. Алгашқы кезеңде ацетил - КоА-дан бета-гидрок-си - бета-метилглутарил - КоА (ГМТ-КоА) синтезделеді. Ары қарай редуктазаның эсерінен тотықсызданған НАДФ (НАДФН2) қатысуымен ГМГ-КоА мевалон қышқылына айнала-ды, одан бірқатар айналулардан кейін диметилаллилпирофосфат молекуласы түзіледі. Одан ары диметилаллилпирофофаттың 2 молекуласынан геранилпирофосфат (С10) синтезделеді. Геранилпирофосфатқа диметилаллилпирофосфаттың тағы бір молекуласы (С5) қосылады да, фарнезилпирофосфат (С15)түзіледі. НАДФН2-нің қатысуымен бірқатар реакциялардан кейін фарнезилпирофосфаттың 2 молекуласынан сквален (С30), одан ары ланостерин жэне холестерин (С 27) синтезделеді.Холестерин синтезінің жылдамдығы кері теріс байланыс механизмы бойынша реттеледі. Реттеудің негізгі тармагы - мевалон қышқылын синтездейтін ГМГ-КоА-редуктаза ферменті болып табылады. Холестерин өзінің синтезінің осы ферментін ингибиторлейді.
Адамның тэуліктік тағамында холестерин мөлшері 2-3 гр болғанда хоелстериннің өз синтезі тоқтайды. Егер холестерин тағамда болмаса, онда ткнаьдерде оның синтезі өте жоғары жылдамдықпен жүреді. Холестерин тағаммен көбірек түскен сайын, тканьдерде холестерин аз синтезделеді жэне агзадан шығаралатын холестеринді тагам холестерині курайды.
Өт қышқылдары ішекте майлардың эмульгациялануына жэне олардың қорытылу өнімдерінің сіңірілуіне қатысады. Соның салдарынан, от қышқылдарының негізгі бөлігі ішектен клеткаларға сіңіріледі де, қақпалы вена арқылы бауырға келіп түседі және қайтадан өт түзілуі үшін пайдаланылады. Өт қышқылдарының біраз бөлігі (тэулігіне 0,5г) агзадан нэжіспен шыгарылады. Аралас мицеллалардың күрамында өт қышқылдарымен бірге холестерин де шыгарылады. Өт қабынын жэне бауырдың өтінде холестерин этерифицирленген түрде болады, липидті комплекстын макромицелласының курамында. Ішекте бул комплекс ыдыраған кезде холестерин босап шығады жэне оның жартылай реабсорбциясы жүреді. Өт қышқылдары, холэстеразаның белсенділігі, тағамның белоктарының, лактозаның жэне басқа факторлардың болуы холестериннің ішек қабырғасына реабсорбциялануына эсер етеді.
Ішек қабырғасында сіңірілмеген холестерин ішек микрофлорасының эсерінен ко-простеринге дейін гидролизденеді де, агзадан нэжіс курамында шыгарылады. Агзадан өттің курамында нэжіспен шыгарылатын жэне өт қышқылдары түрінде шығарылатын холестериннің жалпы мөлшері ересек адамда тэулігіне шамамен 1,3 гр болады. Тағаммен түсетін жэне жіңішке ішектің төменгі бөлімдерінде өтпен шыгарылатын холестерин ішектің клеткаларына сіңіріледі де, қайтадан ТӨТЛП-дердің қурамына кіреді. Қалыпты стационарлы жагдайда холестеринның ішекке тағаммен түсетін мөлшері мен тканьдерде синтезделетін холестериннің қосынды молшері экскрецияланатын холестерин мен от қышқылдарына айналған жэне от қышқылдары түрінде экскрецияланатын холестериннің қосында мөлшеріне тең: ХОЛ (таг) + ХОЛ(синт) = ХОЛ(экскр) + Өт қышқ.(экскр).
Ағзадағы холестериннің қалыпты, турақты деңгейі осы процестерге байланысты болады. Осы баланстың бузылу салдарының бірі- гиперхолестеринемия болып табылады, оның зардаптары өтке тас байлану ауруы, жүректің ишемия ауруы жэне атеросклероз.
Ішекте сіңірілген аминқышқылдары қақпалы вена жүйесі арқылы бауырға келіп түседі де, эртүрлі анаболитикалық жэне катаболитикалық айналымдарға қосылады. Гепатоциттар-да жэне ретикулоэндотелиальды жүйенің клеткаларында аминқышкылдары эртүрлі белоктар синтезі үшін пайдаланылады. Бір тәулік ішінде бауырда шамамен 12 г альбуминдер синтезделеді. Бауырда қан плазамасының а жэне р - глобулиндерінің коп бөлігі, фибриноген, протромбин жэне гемокоагуляция жүйесі мен қан үюға қарсы жүйесінің басқа да фак-торлары синтезделеді. Бауыр ишемиясы, цирроз, токсикалық гепатит, өттің шығарылуының бұзылуы кезінде бауырда альбуминдер синтезі төмендейді. Ретикуло—эндотелиальды жүйе клеткаларының қызметі зақымдалған кезде глобулиндер синтезі төмендейді. Паренхиматоз-ды гепатиттерде бауырдың полигональды клеткаларында РНҚ синтезі төмендейді, ал купфер клеткаларында жэне перипортальды мезенхимальды клеткаларда оның мөлшері артады. Мүның барлығы қан плазмасында альбуминдер мөлшерінің төмендеуіне жэне а, р -глобулиндер мөлшерінің артуына экеледі. Қан плазмасының белокты фракцияларының бүл өзгерістерін альбуминдердің жетіспеушілігінен туындаған қанның онкотикалық қысымын түрақтандыруға бағытталған компенсаторлы реакция деп қарастыруға болады. Бүдан көретініміздей, патологиялық процеске аз ілінген клеткаларда белоктар биосинтезін күшейту арқылы қанның осмостық қысымын түрақтандыруда бауыр реттеуші қызмет атқарады. Патология кезінде бауырда синтезделетін белоктардың сапалық өзгерістері болуы мүмкін. Мысалы, бауыр циррозында қан плазмасы белоктарының сульфгидрильді топтарының мөлшері төмендейді, бүл олардың электрофорездік қозғалғыштыгы мен иммунологиялық қасиеттерінің өзгеруіне экеледі. Аномальды белоктар - парапротеиндер қалыптасады, олар өздерінің физико-химиялық қасиеттері бойынша гамма-глобулиндерге үдсас болады.
Бауырда темірдің тасымалдануын (ферритин, трансферритин), мыстың (церулоплаз-мин), гормондардың (транккортин), витаминдер мен липидтердің (ӨТТЛП, ТТЛП, ЖТЛП) тасымалдануын қамтамасыз ететін белоктар синтезделеді. Соматомединдердің 7 түрі (А1,А2,В1,В2,ВЗ,В4,С) синтезделеді, олар гипофиздың соматотропты гормонының эсерін іске асыруда байланыстырушы қызмет атқарады. Адам ағзасында қайта синтезделетін белоктардың шамамен 50% -ы бауырда түзіледі. Белоктық ашығу кезінде басқа тканьдерді аминқышқылдарымен камтамасыз ету үшін бауыр өзінің резервті белоктарын іске қосады. Гепатоциттерде темірпорфириндердің, цитохром жүйесі ферменттерінің қүрылымдық компоненттерінің, миоглобин мен гемоглобиннің синтез процестері жүреді, бірақ гемоглобин биосинтезінде сүйек миы негізгі роль атқарады, мұнда эритроциттердің жетілуі жүреді. Бүл синтездердің бастапқы субстраттары - глицин жэне сукцинил - КоА. Гемоглобин гемінің синтез механизмы 13 тарауда 13.2 бөлімде көрсетліген.
Бауырда аминқышқылдардың өзара айналу процестері, ауыстырылатын аминқышқылдары мен азотты белокты емес қосылыстардың (холин, креатин, глутатион, пуринды жэне пиримидинді негіздер порфириндер) синтезі қарқынды жүріп жатады. Сонымен қатар, бауырда глюкогенді аминқышқылдардың азотсыз қалдықтарынан глюконеогенез процестері де қарқынды жүреді.
Бауырда мочевинаның түзілуімен өтетін аммиакты залалсыздандыру процесі жүреді (мочевина түзілуінің орнитинды циклы). Аммиак пен көмірқышқыл газының фиксациясы кезінде түзілген карбомоилфосфат бауырда пуринды нуклеотидтердің синтезі үшін пайдаланылады.
Бауырдың купфер клеткаларында гемоглобин гемінің билирубинге дейін ыдырауы, от пигменттерінің глюкурон қышқылымен коньюгаттар түзуі жүреді жэне олар өт қүрамында шығарылады.

