




Ферменты рестрикции стали эффективным инструментом исследования. Они позволяют превращать молекулы ДНК очень большого размера в набор фрагментов длиной от нескольких сотен до нескольких тысяч оснований. С помощью метода электрофореза в агарозном геле (см. раздел 1) фрагменты ДНК, различающиеся по размеру, можно легко разделить, а затем исследовать каждый фрагмент отдельно.
Короткие фрагменты мигрируют намного быстрее, чем длинные. При сравнительно высокой концентрации агарозы большие фрагменты вообще не могут проникнуть в гель. В процессе миграции рестрикционные фрагменты не деградируют, их можно элюировать (вымывать) в виде биологически активных двуцепочечных молекул. При окрашивании гелей красителями, связывающимися с ДНК, выявляется набор полос, каждая из которых отвечает рестрикционному фрагменту, молекулярную массу которого можно определить, проведя калибровку с помощью ДНК с известными молекулярными массами.
Сравнение размеров фрагментов ДНК, полученных после обработки определенного участка генома набором рестрицирующих нуклеаз позволяет построить рестрикционную карту, на которой указано положение каждого сайта рестрикции относительно других участков.
Молекулу ДНК длиной 5000 пар нуклеотидов (п. н.). обрабатывают отдельно рестриктазами А и В. Фрагменты разделяют электрофорезом. Фермент А разрезал ДНК на 4 фрагмента размером 2100, 1400, 1000 и 500 п. н. Обработка рестриктазой В дала 3 фрагмента: 2500, 1300 и 1200 п. н. (рис. 37). Для определения расположения сайтов рестрикции этих ферментов на следующем этапе применяют процедуру двойного расщепления – обрабатывают ДНК двумя эндонуклеазами. Обработка изучаемого фрагмента одновременно двумя рестриктазами дала 6 фрагментов: 1900, 1000, 800, 600, 500, 200 п. н.
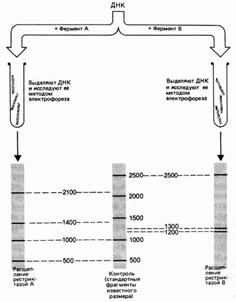
Рис. 37. Результаты электрофореза после обработки фрагмента ДНК разными рестриктазами
Наиболее полный вариант – элюировать каждый фрагмент, образующийся в результате расщепления одной рестриктазой, а затем обработать его второй. Смесь фрагментов, полученных после такой обработки, также анализируют с помощью электрофореза. В нашем примере были получены следующие результаты:
Обработка каждого из 4-х А-фрагментов рестриктазой В
2100 - 1900 и 200,
1400 - 800 и 600,
1000 - 1000 (изменений нет)
500 - 500 (изменений нет)
Обработка каждого из 3-х В-фрагментов рестриктазой А
2500 - 1900 и 600
1300 - 800 и 500
1200 - 1000 и 200
Анализ полученных результатов показывает, что каждый из ферментов, полученный при расщеплении А-фрагментов рестриктазой В можно обнаружить в образцах, полученных при расщеплении В-фрагментов рестриктазой А. Ключом к рестрикционному картированию являются перекрывающиеся фрагменты. Такими в рассматриваемом примере являются В-фрагмент 2100 и А-фрагмент 2500. При обработке другой рестриктазой они дают фрагмент 1900.
Из данных о расщеплении этих фрагментов мы предполагаем, что с одной стороны на расстоянии 200 п. н. от фрагмента 1900 находится следующий А-сайт, а с другого конца, на расстоянии 600 п. н. – следующий В-сайт (рис. 38). При обработке двумя эндонуклеазами фрагмент 200 п. н. образуется 1 раз, при обработке рестриктазой А из В-фрагмента 1200, т. е. фрагмент 1200 лежит слева. Остается определить, как продолжается карта вправо. Очевидно, это А-фрагмент 1400, так как он рассечен рестриктазой В на фрагменты 600 и 800. Вправо от фрагмента 2500 следует отложить, очевидно, фрагмент 1300. Тогда логично наличие А-фрагмента 500 и деления В-фрагмента 1300 рестриктазой А на 800 и 500.
При построении рестрикционных карт обычно используют несколько рестриктаз, поэтому приходится анализировать сложные соотношения между фрагментами, полученными при действии разных ферментов. Для упрощения процедуры картирования можно применять неполное расщепление. В определенных условиях рестриктаза узнает и расщепляет не все сайты в молекуле ДНК. Например, при частичном расщеплении ДНК ферментом А могут образовываться фрагменты 3100 п. н., 1400 п. н. и 500 п. н. Сопоставив их с данными полного расщепления (2100, 1400, 1000 и 500), можно сразу поставить рядом 2100 и 1000 (фрагмент 3100). А получив фрагмент 3500 – расположить рядом 2100 п. н. и 1400 п. н.
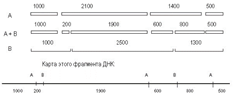
Рис. 38. Анализ фрагментов рестрикции и карта фрагмента ДНК
Другой прием – введение радиоактивной концевой метки. Концевые фрагменты определяются в этом случае по включению метки. Можно также сопоставить фрагменты путем гибридизации нуклеиновых кислот. Перекрывающиеся фрагменты (в данном случае 2100 и 2500) будут гибридизоваться.
Первая карта была получена для вируса SV40 (обезьяний вирус, вызывающий злокачественную трансформацию), содержащего 5423 пары оснований. Использовали рестриктазу Hind-II, расщепляющую кольцевую ДНК вируса на 11 фрагментов. Порядок их расположения в ДНК был установлен путем исследования наборов фрагментов, образующихся по мере того, как расщепление доходит до конца. Первый разрыв превращал кольцевую молекулу в линейную, которая затем расщеплялась на все меньшие и меньшие фрагменты. Исследовали вначале наборы перекрывающихся фрагментов, а затем продукты полного расщепления. Таким образом была получена рестрикционная карта кольцевой вирусной ДНК, на которую были нанесены сайты расщепления рестриктазой. Повторив подобные эксперименты с другой рестриктазой можно получить более подробную карту, где отмечено много сайтов рестрикции.
Располагая такой информацией, можно идентифицировать на ДНК биологически важные участки. Поскольку рестрикционная карта отражает расположение определенной последовательности нуклеотидов в данном участке, сравнение таких карт для двух или более родственных генов позволяет оценить гомологию между ними. Анализируя рестрикционные карты, можно сравнивать определенные участки ДНК разных видов животных без определения их нуклеотидной последовательности. Таким образом, например, было установлено, что хромосомные участки, кодирующие цепи гемоглобина у человека, орангутанга и шимпанзе сохранились в практически неизменном виде в течение последних 5 - 10 млн. лет (с тех пор как виды дивергировали).
Метод рестрикционного картирования позволяет увидеть крупные генетические изменения, такие как делеции или инсерции. При этом происходит уменьшение или увеличение рестрикционных фрагментов, а также исчезновение или возникновение сайтов рестрикции.
Один из приемов картирования – фингерпринт («метод отпечатков пальцев» или DNA-fingerprint). Он подразумевает использование неупорядоченных и неполных наборов фрагментов, которые являются характеристикой генома, хотя описывает его не полностью.
10. Блоттинг -методы молекулярной биологии по переносу определённых белков или нуклеиновых кислот из раствора, содержащего множество других молекул, на какой-либо носитель (мембрану из нитроцеллюлозы,PVDF или нейлона) в целях последующего анализа.
Метод Саузерн-блот для определения позиции гена в геноме; метод Нозерн-блот для определения экспрессии генов (анализ мРНК);
Метод Саузерн-блот.
Для анализа генов необходимо определить локализацию сайтов рестрикции. Эта информация используется для сравнения гомологичных генов разных видов, для анализа организации интронов, для выбора фрагмента, подвергающегося клонированию. Расположение сайтов рестрикции в изучаемом гене можно определить без клонирования гена, используя молекулярные зонды (меченные радиоактивно или флуоресцентным красителем), комплементарные изучаемой последовательности. Анализ состоит из последовательности следующих этапов:
(1) выделение высокомолекулярной геномной ДНК из клетки;
(2) ферментативное расщепление ДНК разными ре- стриктазами с образованием фрагментов разного размера;
(3) разделение рестрикционных фрагментов в агарозном геле;
(4) денатурация фрагментов ДНК щелочным раствором;
(5) нейтрализация геля буферным раствором;
(6) капиллярный перенос фрагментов на нейлоновый или нитроцеллюлезный фильтр
(7) гибридизация с одноцепочечными радиоактивными зондами;
(8) авторадиография для выявления гибридов: исследуемая ДНК / зонд.
В результате анализа полученного материала можно определить наличие или отсутствие некоторых сайтов рестрикции, характерных для изучаемого гена, которые ассоциируются с определенными мутациями. Различные вари- анты расположения сайтов рестрикции в ДНК у двух разных людей называют Полиморфизмом Длины Рестрик- ционных Фрагментов (ПДРФ). Этот полиморфизм ис- пользуется как генетический маркер в изучении генотипа.
Метод Норзерн-блот.
Метод состоит в переносе разделенных молекул РНК на нейлоновые или нитроцеллюлезные фильтры с последующей гибридизацией с мечеными зондами. Метод схож с методом Саузерн-блот, за исключением того, что выделенные и очищенные мРНК не подвергают рестрикции, а электрофорез происходит в денатурирующих условиях. Метод Норзерн-блот позволяет определить транскрипты анализируемых генов, кДНК, размеры генов.
11. Методы ДНК - зондов основаны на реакции гибридизации нуклеиновых кислот — способности односпиральных комплементарных цепей ДНК или РНК формировать двуспиральные структуры. Реакция может протекать между комплементарными молекулами вида: ДНК + ДНК, ДНК + РНК, РНК + РНК.
У большинства ДНК-содержащих вирусов (адено-, герпес-, покс-, папилломавирусы и др.) нуклеиновая кислота находится в виде двух цепей, которые связаны (водородная связь) комплементарно через азотистые основания (аденин—тимин, гуанин—цитозин).
Так, у аденовирусов таких нуклеотидных пар 35—36,5 тыс., у герпесвирусов — 125—240 тыс., у поксвируеов — 130—375тыс. и т. д.
Любой вирус включает одну специфическую для него молекулу ДНК (или РНК) со строго определенной последовательностью нуклеотидов. Чтобы в исследуемом материале обнаружить вирусную нуклеиновую кислоту, можно воспользоваться ее способностью после разделения цепей (если она двуспиральная) образовывать снова двойную цепь с комплементарной ей молекулой нуклеиновой кислоты, предварительно как-либо помеченной. Такая меченая односпиральная молекула нуклеиновой кислоты, комплементарная молекуле нуклеиновой кислоты определенного вируса, называется ДНК-зондом. Для каждого вида вируса зонд готовят заранее и вводят в него метку в виде атома радиоактивного фосфора (32Р) или в виде биотина, или другим веществом.
Методы ДНК-зондов сводятся к следующему:
1) получение односпирального фрагмента ДНК определенного вируса и его метки — это ДНК-зонд;
2) выделение из исследуемого материала нуклеиновых кислот и их денатурация;
3) контакт образовавшихся односпиральных молекул ДНК (или РНК) с ДНК-зондом при 55—65 °С, приводящих к образованию двуспиральных молекул (гибридизация) в случаях их взаимной комплементарности;
4) удаление всех негибридизированных односпиральных молекул нуклеиновых кислот (отмывают или добавляют нуклеазы);
5) обнаружение (по метке) образовавшихся двуспиральных молекул нуклеиновых кислот, которые и будут указывать на наличие в исследуемом материале того вируса, на который был получен ДНК-зонд.
Если использовали радиоактивный зонд, то результаты учитывают методом авторадиографии или путем подсчета импульсов в счетчике.
Существует большое количество модификаций метода гибридизации с мечеными нуклеиновыми кислотами. Их можно разделить на две основные группы: способы гибридизации в растворе и способы гибридизации на твердых носителях.
Наиболее популярен метод гибридизации с использованием носителей — нитроцеллюлозных или нейлоновых фильтров и твердых полимеров. Он заключается в том, что одну нуклеиновую кислоту (чаще исследуемую), предварительно денатурированную, иммобилизуют на носителе (фильтре), проводят гибридизацию с ДНК-зондом и непрореагировавшие молекулы зонда отмывают, а иммобилизованные меченые гибриды остаются на фильтре. Сигнал, подаваемый связанным зондом, обнаруживают посредством авторадиографии, если зонд был радиоактивным, или путем образования цветных пятен, если использовали меченный ферментом зонд.
Достоинства метода: высокая чувствительность и специфичность; быстрота анализа; универсальность; отсутствие необходимости в стерильной работе, математической обработке результатов. Недостатки: относительная технологическая сложность и трудность получения зонда; если метка радиоактивная, то нельзя зонд нарабатывать впрок, так как период полураспада 32Р — 14 сут, а1311 — 8 сут; если метка нерадиоактивная, то чувствительность метода снижается в 10 раз и более.
Молекулярные маяки — олигонуклеотиды, содержащие на 5'-конце тушитель флюоресценции (нефлюоресцирующий хромофор), а на 3'-конце флюорохром; нуклеотидные последовательности, прилежащие к красителям, самокомплементарны, в результате чего в М.м. формируется структура типа «стебель-петля», и флюоресценция флюорохрома подавлена. Область петли М.м. комплементарна анализируемому продукту полимеразной цепной. После образования гибрида между М.м. и продуктом ПЦР флюорохром начинает флюоресцировать («маяк» зажигается). М.м. используются в качестве зондов в одном из вариантов ПЦР в реальном времени.
Недостаток радиоактивных зондов — их нестабильность из-за непродолжительного времени жизни изотопов (32Р — 14,5 сут) и быстрого радиолиза зонда. Использование изотопов с более продолжительным периодом полураспада приводит к снижению чувствительности и удлиняет время, необходимое для радиоавтографии. Кроме того, для работы с изотопами необходимы специально оснащенные лаборатории.
12. Гибридизаци я - соединение in vitro комплементарных одноцепочечных нуклеиновых кислот в одну молекулу. При полной комплементарности объединение происходит легко и быстро, а в случае частичной некомплементарности слияние цепочек замедляется, что позволяет оценить степень комплементарности. Возможна гибридизация ДНК-ДНК и ДНК-РНК.
Есть доводы в пользу гибридизации:
-с помощью зондов (зонд — определенный фрагмент ДНК, содержащего известный ген — генетический маркер) можно обнаружить микроорганизмы, которые трудно культивируются in vitro;
-зонды позволяют отличить вирулентные штаммы от авирулентных in vitro;
-быстрая идентификация микроорганизма непосредственно в биологическом материале от больных животных, пробах из окружающей среды и в фиксированных спиртом или формалином образцах после хранения;
-нет необходимости направлять образцы для идентификации в специализированные лаборатории и институты;
-высокая достоверность выявления единичных микробных клеток.
Принцип метода. Метод основан на специфическом взаимодействии (гибридизации) двух образцов ДНК: меченого ДНК-зонда и участка плазмидной или хромосомной ДНК. Образовавшиеся двухцепочечные ДНК отделяют от одноцепочечных и определяют присутствие метки. В качестве контроля используют ДНК известного штамма микроорганизма. Степень реассоциации таких гомологичных одноцепочечных ДНК принимают условно за 100 %. При 60…100 % гомологии ДНК можно говорить о принадлежности сравниваемых бактерий к одному виду.
Проведение реакции. Разработаны два способа гибридизации: в растворе или с использованием нерастворимого носителя. Гибридизация на носителях применяется значительно шире: анализируемый образец связывается с нитроцеллюлозным или нейлоновым фильтром.
Существует несколько разных методов мечения комплекса между зондом и ДНК-мишенью. Традиционным способом является радиоактивное мечение зонда с помощью введения в ДНК радиоактивных изотопов 33Р, 32Р, 35S. После гибридизации образцы-мишени выявляют с помощью радиоавтографии. Этот метод обладает высокой чувствительностью и позволяет обнаруживать около 1 пг ДНК. Недостаток радиоактивных зондов — их нестабильность из-за непродолжительного времени жизни изотопов (32Р — 14,5 сут) и быстрого радиолиза зонда. Использование изотопов с более продолжительным периодом полураспада приводит к снижению чувствительности и удлиняет время, необходимое для радиоавтографии.
Наиболее широко используют нерадиоактивное мечение. Для нерадиохимической детекции применяют методы, основанные на присоединении к зондам остатков биотина. Метод дот-гибридизации. В случаях, когда необходимо установить наличие или отсутствие в ДНК выявляемой последовательности, применяют метод дот-гибридизации. В этом случае очищенную денатурированную ДНК наносят непосредственно на фильтр, иммобилизуют, гибридизируют с зондом и регистрируют на автографах положительные сигналы.
13. Электрофорез — это метод разделения веществ в электрическом поле на основании разницы в заряде молекул. Методом электрофореза в лабораторной практике разделяют белки и нуклеиновые кислоты.
Электрофорез ДНК в агарозном геле — это аналитический метод, применяемый для разделения фрагментов ДНК по размеру (длине). Плазмидная ДНК образует вторичные структуры, например, скручивается, в суперспираль. Таким образом, чтобы определить размер плазмиды, необходимо разрушить элементы вторичной структуры. Для этого перед нанесением на гель плазмиду «линеризуют», то есть обрабатывают одной из рестриктаз (эндонуклеаз). Рестриктазу выбирают так, чтобы она разрезала плазмиду только в одном месте.
Для разделения фрагментов ДНК разной длины используют гель с различной концентрацией агарозы. Чем меньше длина разделяемых фрагментов ДНК, тем больше должна быть концентрация агарозы:
| Длина разделяемых фрагментов ДНК | Концентрация агарозы, % |
| 50-1500 п. р. | |
| 300-3000 п. о. | 1,5 |
| 400-6000 п. о. | 1,2 |
| 500-10000 п. о. | |
| 800-10000 п. о. | 0,7 |
| 1000-20000 п. о. | 0,5 |
При проведении электрофореза фрагменты ДНК мигрируют в геле под воздействием сил электрического поля. Дело в том, что сахарофосфатный остов молекул ДНК заряжен отрицательно и поэтому цепи ДНК двигаются от катода, заряженного отрицательно, к положительному аноду. Более длинные молекулы мигрируют медленнее, так как задерживаются в геле, более короткие молекулы двигаются быстрее.
Перед началом электрофореза к образцам принято добавлять два разных красителя с кислым значением pH (часто для этих целей используют, ксиленовый голубой и бромфеноловый синий), чтобы визуализировать ход электрофореза и утяжелять образцы глицерином (до 20 % глицерина в образце). Краситель также необходим для того, чтобы определить, когда стоит остановить процесс. Электрофорез проводится в камере, заполненной буферным раствором. Чаще всего используются буферы, содержащие ЭДТА, трис, а также борную или уксусную кислоты. Соответственно, буфер, содержащий борную кислоту называется TBE (tris-borate-EDTA), а буфер, содержащий уксусную кислоту TAE (tris-acetate-EDTA). Концентрации маточных растворов TBE и TAE составляют 6x и 50x, соответственно, если сравнивать с их рабочими концентрациями Буфер необходим для повышения ионной силы раствора, в котором будет происходить разделение молекул ДНК под действием приложенного электрического поля.
После разделения (иногда краситель вносят в расплавленную агарозу) фрагменты ДНК разной длины визуализируют при помощи флюоресцентных красителей, специфично взаимодействующих с ДНК, например, агарозные гели обычно красят бромистым этидием, который интеркалирует между азотистыми основаниями дуплекса и флюоресцирует в УФ-лучах.
Определение размеров производят путём сравнения наборов коммерческих фрагментов ДНК известной длины («DNA ladder», «линейка», «маркеры ДНК»), и ДНК в проверяемых образцах Обычно в коммерческих маркерах также указывают содержание отдельных фрагментов, чтобы можно было, сопоставив интенсивность полос, быстро оценить концентрацию ДНК в проверяемых образцах. При необходимости ДНК маркеры для геля можно изготовить самостоятельно, если обработать плазмиду рестриктазой, которая режет её в нескольких местах. Для этого выбирается эндонуклеаза, при действии которой образуются 5-6 фрагментов, различной длины.
ГОРИЗОНТАЛЬНЫЙ ЭФ
Проводится в пластине агароз. геля, кот. предст. собой заст. в эф буфере агарозу 0,5 – 2,5 %.Застывшая агароза образует простр. решетку, для формирования лунок исп. гребенку. Подвижность зависит т концентрации агарозы, напряженности эл.поля, состава буфера, температуры.
Вертикальный ЭФ
исп. акриламид. ЭФ проходит в вертикальной плоскости. Преимущество: большая разрешающая способность. Позволяет различать молекулы ДНК, отлич. на 1 основание.
Недостатки: 1) больше времени на проведение. 2) акриламид токсичный. 3) нельзя автоматизировать.

