




Кетоновые тела ‒ ворастворимые соединения, поэтому легко транспортируются через внутреннюю мембрану митохондрий, также как и через гемато-энцефалический барьер и клеточные мембраны. В связи с этим они могут использоваться в качестве источника энергии различными тканями, включая ЦНС (рис.22.10).
• ‒ D-b-гидроксибутират окисляется до ацетоацетата при участии NAD+-зависимой b-гидроксибутиратдегидрогеназы, при этом продуцируется молекула NADH (1 ).
• ‒ Следующая стадия ‒ присоединение HSCoA. Синтез ацетоацетил-СоА происходит путем реакции ацил-тиоэфирного обмена с участием сукцинил-СоА ферментом СоА-трансферазой (2). Сукцинил-СоА ‒ интермедиат цикла лимонной кислоты.
• ‒ При участии тиолазы (3) и еще одной молекулы HSСоА ацетоацетил-СоА расщепляется на 2 молекулы ацетил-СоА.

Рис.22.10. Кетоновые тела ‒ клеточное топливо
Кетоновые тела являются предпочтительными субстратами для аэробных мышц и сердца, чем глюкоза, когда они доступны. В периферических тканях кетоновые тела должны вновь превратиться в ацетил-СоА в митохондриях.
Глиоксилатный цикл
Растения и некоторые бактерии способны использовать ацетил-СоА, образующийся при катаболизме липидов (или из любого другого источника), не только для производства энергии, но и в качестве источника углерода для синтеза большинства других классов соединений. Животным также присуща такая универсальность, однако они существенно менее эффективно могут использовать углерод липидов для синтеза углеводов. Причина этого заключается в том, что растения и бактерии способны синтезировать два дополнительных фермента, изоцитрат-лиазу и малат-синтазу; оба этих фермента не синтезируются в организме животных.
Вместе с некоторыми ферментами цикла лимонной кислоты эти дополнительные ферменты участвуют в глиоксилатном цикле. В семенах растений, запасающих масло, глиоксилатный цикл осуществляется в отдельных органеллах, глиоксисомах.
Пероксисомальная/глиоксисомальная система отличается от митохондриальной двумя аспектами:
1) в первой окислительной реакции электроны переносятся на кислород, образуя воду, и 2) восстановленный NAD, образованный во второй окислительной реакции не реокисляется в пероксисомах или глиоксисомах, поэтому восстановительные эквиваленты переносятся в цитозоль и далее транспортируются в митохондрии (рис.22.11).
| Митохондрия | Пероксисома/глиоксисома |
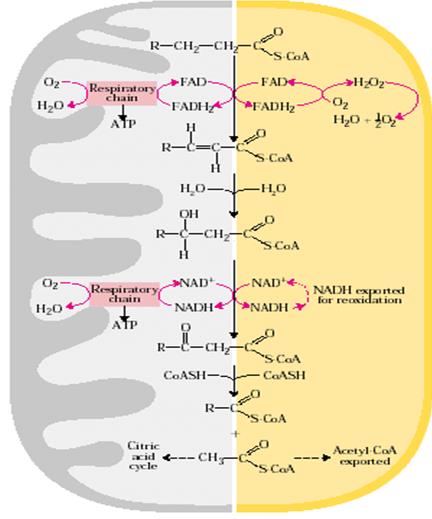
Рис. 22.11. Сравнение процесса β-окисления в митохондрии и пероксисоме/глиоксисоме
Ацетил-СоА, продуцированный в пероксисомах и глиоксисомах также экспортируется: ацетат из глиоксисом используется как биосинтетические предшественники. Ацетил-СоА, образованный в митохондриях, в дальнейшем окисляется в цикле лимонной кислоты.
ЛЕКЦИЯ 23

