




В транспорте ацил-СоА участвуют карнитин, который связывает молекулу жирной кислоты особым образом, в результате чего положительный (на атоме азота) и отрицательный (на атоме кислорода карбоксильной группы) заряды оказываются сближенными и нейтрализуют друг друга, и три белка ‒ карнитинацилтрансфераза I, локализованная во внешней митохондриальной мембране, карнитинацилкарнитин-транслоказа и карнитинацилтрансфераза II, локализованные во внутренней мембране митохондрий (рис.22.2).
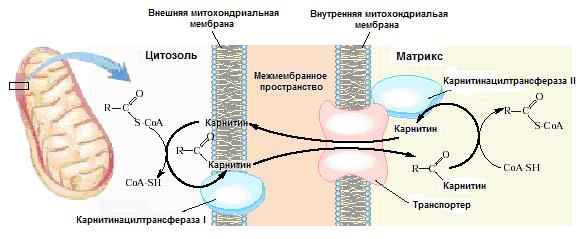
Рис.22.2. Транспорт длинноцепочечных ацил-СоА из цитозоля в матрикс митохондрий
СоА-эфиры жирных кислот взаимодействуют с гидроксигруппой карнитина (γ-триметиламино-β-оксибутират). На внешней стороне наружной мембраны протекает транс -этерификация, катализируемая карнитинацилтрансферазой I, и полученный сложный эфир поступает в матрикс, причем его диффузия усиливается специальным ацилкарнитин/карнитин-транспортером, локализованным во внутренней мембране митохондрий.
Процесс завершается переносом ацильного остатка от карнитина к внутримитохондриальному СоА при участии фермента карнитинацилтрансферазы II, локализованной на внутренней поверхности внутренней митохондриальной мембраны. Транспорт ацил-СоА, опосредованный карнитином, является стадией, лимитирующей скорость окисления жирных кислот.
Окисление жирных кислот
β-окисление ‒ циклический процесс, каждый цикл которого заключается в удалении двууглеродного фрагмента в форме ацетил-СоА, начиная от карбоксильного конца жирной кислоты. Цикл включает четыре реакции: дегидрирование, гидратацию, дегидрирование и тиолитическое расщепление. На рис.22.3 приведена схема β-окисления насыщенной жирной кислоты с четным числом атомов углерода.
В первой реакции происходит окисление ацил-СоА при участии FAD-зависимой ацил-СоА-дегидрогеназы. Три вида ацил-СоА-дегидрогеназ осуществляют окисление длинно-, средне- и короткоцепочечных ацил-СоА эфиров жирных кислот. Образовавшийся в реакции FADH2 в составе ацил-дегидрогеназы окисляется другим флавопротеином, переносящим электроны в митохондриальную электронтранспортную цепь и далее к кислороду. При этом происходит синтез АТР (два моля на одну пару перенесенных электронов). Окисление с участием ацил-СоА-дегидрогеназ, аналогично дегидрированию, катализируемому сукцинатдегидрогеназой в цикле лимонной кислоты.
Продукт окисления ‒ еноил-СоА гидратируется под действием еноил-гидратазы с образованием β-гидроксиацил-СоА. Существуют еноил-СоА-гидратазы, проявляющие специфичность к цис- или транс -формам еноил-СоА-производных жирных кислот. При этом транс- еноил-СоА гидратируется стереоспецифически в L-β-гидроксиацил-СоА, цис -изомеры ‒ в D-β-гидроксиацил-СоА.
В третьей реакции цикла β-окисления происходит дегидрирование L-β-гидроксиацил-СоА специфической только к L-изомерам β-гидроксиацил-СоА NAD+-зависимой дегидрогеназой. Окисление подвергается β-углеродный атом молекулы. Образующийся β-кетоацил-СоА в завершающей реакции цикла легко расщепляется тиолазой с образованием двух продуктов: ацил-СоА, укороченного по сравнению с исходным на два углеродных атома, и ацетил-СоА ‒ двууглеродной молекулы, отщепленной от жирнокислотной цепи. Ацил-СоА-производное подвергается следующему циклу реакций β-окисления, а ацетил-СоА вступает в цикл лимонной кислоты для дальнейшего окисления.
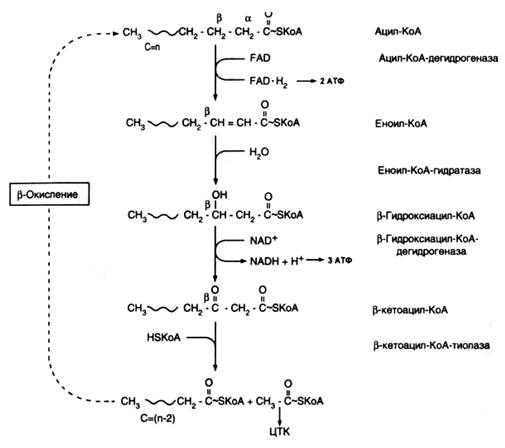
Рис.22.3. β-окисление насыщенных жирных кислот
Рассмотрим энергетический баланс окисления на примере стеариновой кислоты.
Уравнение β-окисления стеароил-СоА:
СН3(СН2)16-СО~SCоА + 8 HSCoA + 8 FAD + 8 NAD+ + 8Н2О →
9 СН3-СО~SCоА + 8 FADH2 + 8 NADH + 8H+ (22-1)
Ацетил-СоА, образованный при окислении жирных кислот, далее подвергается окислению до СО2 и Н2О в цикле лимонной кислоты. Следующее уравнение ‒ результат окисления стеароил-СоА в ЦТК (9 циклов):
9 СН3-СО~SCоА + 9 FAD + 27 NAD+ + 9 GDP + 9 Pi +27 Н2О →
18 СО2 +9 HSCоА + 9 FADН2 + 27 NADН +27Н+ + 9 GTP (22-2)
Уравнения окислительного фосфорилирования:
17 FADН2 + 8,5 О2 + 34 ADP + 34 Pi → 17 FAD + 17 Н2О + 34 ATP
35 NADН + 35Н+ + 17,5 O2 + 105 ADP + 105 Pi →
35 NAD+ + 35 Н2О + 105 ATP (22-3)
Комбинируя уравнения (22-2) и (22-3), получаем окончательное уравнение полного окисления cтеароил-СоА до СО2 и Н2О (исходя из предположения, что GDP = ADP и GTP = ATP):
СН3(СН2)16-СО~SCоА + 26 О2 + 148 Рi + 148 ADP →
18 СО2 + 17 Н2О + 148 АТР + HSCоА (22-4)
Таким образом, катаболизм жирных кислот обеспечивает продукцию энергии. Расчет выделяемой энергии удобно проводить по формуле:
[5(n/2 ‒ 1) + n/2∙12 ‒ 2],
где 5 ‒ число молекул АТР, образуемое при одном акте β-окисления; n ‒ число атомов углерода в ЖК; n/2 ‒ 1 ‒ число актов окисления; n/2 ‒ число молекул ацетил-СоА; 12 ‒ число молекул АТР, образующихся при полном окислении одной молекулы ацетил-СоА в цикле лимонной кислоты; 2 ‒ число молекул АТР, затраченных на активацию жирной кислоты.
При полном β-окислении стеариновой кислоты кислоты образуется 148 молекул АТР. С учетом двух молекул АТР, затраченных на активирование молекулы стеарата (образование стеароил-СоА), общий энергетический выход при окислении одной молекулы стеариновой кислоты в организме животных и человека составит 148 ‒ 2 = 146 молекул АТР. Рассчитано, что около 40% от всей потенциальной энергии стеариновой кислоты при ее окислении в организме используется для ресинтеза АТР, а оставшаяся часть теряется в виде тепла. Наиболее активно β-окисление протекает в митохондриях печени, почек, скелетной и сердечной мышц.
Примечание: β-окисление является источником эндогенной воды.

