




Гемоцитопоез (від гр. паіта — кров, куіпоз — клітина, роезіз — тво-рення) — складний послідовний процес утворення клітин, у результаті якого досягається постійність кількісного та якісного складу форме-них елементів крові. Розрізняють кровотворення після народження протягом усього життя тварини та кровотворення у період ембріональ-ного розвитку.
Після народження тварин кровотворення відбувається у мієлоїд-ній (гр. тіеіоз — мозок) тканині червоного кісткового мозку плоских та епіфізів трубчастих кісток, де відбувається розвиток стовбурових клітин і усіх формених елементів крові — еритроцитів, лейкоцитів, кров'яних пластинок. Утворення еритроцитів, лейкоцитів і кров'яних пластинок називають терміном мієлопоез (від гр. туеіоз — мозок).
Кровотворна тканина тимуса, селезінки, лімфатичних вузлів, сли-зових оболонок внутрішніх порожнинних органів називається лімфоїд-ною. У ній утворюються лімфоцити і плазмоцити, а також здійсню-ється диференціація і розмноження Т- тпа В-лімфоцитів. Цей процес називають лімфоцитопоезом.
У птахів лімфоїдна тканина знаходиться у фабрицієвій бурсі, сли-зових оболонках органів травлення, печінці, легенях, лімфоїдному утво-ренні ока тощо. Між органами мієлоїдної та лімфоїдної тканин існує тісна взаємодія, що відбувається шляхом міграції і рециркуляції клітин крові, нейрогуморальної регуляції кровотворення і розвитку крові.
Відповідно до сучасної унітарної теорії кровотворення, усі зрілі клітини крові походять з однієї загальної родоначальної клітини, яку називають стовбуровою кровотворною клітиною (СКК). її диференці-ювання у різні види клітин крові зумовлюється мікрооточенням рети-кулярної тканини органів мієлоїдного та лімфоїдного кровотворення і дією специфічних гемопоетинів. Основи унітарної теорії кровотворен-ня були закладені відомим російським гістологом О.О.Максимовим у 1927р. Такою клітиною є малий лімфоцит, який зберігає здатність перетворюватися у клітини крові. Ця теорія значно поглиблена по-дальшими різноманітними новими методами дослідження — клональ-ними (групи клітин, що утворюються в селезінці з однієї клітини), трансплантації, радіобіологічними, генетичними, імунологічними, електронномікроскопічними. Було встановлено, що стовбурові клі-тини крові (СКК) виникають в ембріональному періоді у жовтковому
9*
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
мішку 7-8 денних ембріонів мишей і потім розселяються у всій крово-творній системі.
У дорослому організмі найбільша кількість СКК міститься у чер-воному кістковому мозку (на 100000 клітин кісткового мозку припа-дає 50 СКК). За допомогою світлового та електронного мікроскопів встановлено, що за своєю будовою СКК подібні до малого лімфоцита.
За даними І.Л.Черткова і А.І.Воробйова (1981), у сучасній схемі кровотворення розрізняють шість класів клітин:
I. — Поліпотентні клітини — попередники СКК;
II. — Частково детерміновані клітини — попередники, потенції цих
клітин частково обмежені, вони здатні до утворення мієлопоезу і лім-
фопоезу;
III. — Уніпотентні клітини — попередники, здатні розвиватися в одному напрямі, під впливом гемопоетинів;
IV. — Бласти, морфологічно розпізнавані проліферуючі клітини — попередники, здатні до мітозу;
V. — Дозріваючі клітини;
VI. — Зрілі клітини, здатні до виходу в кров.
Еритроцитопоез — це розвиток еритроцитів, який відбувається у
червоному кістковому мозку із стовбурової кровотворної клітини. Ця клітина ділиться, диференціюється у клітину — попередника мієло-поезу. її ще позначають як колонієутворюючу одиницю гранулоцитів, еритроцитів, моноцитів, мегакаріоцитів (КУО-ГЕММ), або напівстов-бурову клітину (НСК). 3 цієї клітини утворюються більш детерміно-вані попередники двох видів: КУО-НГЕ (колонієутворююча одиниця нейтрофільних гранулоцитів та еритроцитів) і КУО-МГЦЕ (колоні-єутворююча одиниця мегакаріоцитів та еритроцитів). Таким чином, наступна стадія розвитку еритроцитів — їх уніпотентний попередник КУО-Е розвиваються лише в напрямі еритроцитів. Може утворитися двома шляхами — з КУО-ГНЕ або КУО-МГЦЕ. Уніпотентну клітину еритроцитопоезу називають ще еритроцитопоетинчутливою (ЕЧК), оскільки її подальша диференціація індукується гормоном еритропое-тином, який виробляється у нирках і посилює проліферацію ЕЧК та їх перетворення в проеритробласти. Цей гормон також стимулює розви-ток і розмноження еритроїдних клітин подальших стадій.
Проеритробласти — перші морфологічно розпізнавані клітини еритроїдного ряду, круглої форми, діаметром 15-25 мкм. Ядро вели-
Роздія 3
Загальна гістологія
ке кругле, розміщене центрально, дрібносітчасто-зернистої структури, містить одне-два ядерця; цитоплазма забарвлюється базофільно, на-вколо ядра знаходиться світла перинуклеарна зона. Клітини містять багато рибосом, центросому з двома центріолями, характерною є на-явність зерен феритину і виростів цитоплазми. Проеритробласти поді-ляються і перетворюються на базофільні еритробласти.
Базофільні еритробласти мають трохи менші розміри, порівняно з проеритробластами (10-18 мкм), хроматин у них розміщений гру-дочками променеподібно, як спиці в колесі, цитоплазма базофільна за рахунок великої кількості РНК. Базофільні еритробласти діляться мітозом, починається синтез гемоглобіну і перетворення їх у поліхро-матофільні еритробласти.
Поліхроматофільні еритробласти величиною 10-14 мкм, ядро міс-тить щільний хроматин, чіткої колесоподібної структури, ядерця не ви-значаються. Цитоплазма поліхромна, тобто забарвлюється кислими та основними барвниками. Оксифілія зумовлена наявністю гемоглобіну, а базофілія — РНК. Кількість рибосом у цих клітинах зменшується, фери-тин розміщується агрегатами. Поліхроматофільні еритробласти діляться мітозом, їхні генерації називають поліхроматофільними нормобластами.
Поліхроматофільні нормобласти розміром до 10 мкм, ядро ущіль-нюється, втрачає колесоподібне розміщення хроматину, стає пікнотич-ним. Клітини не здатні до поділу, більшість їх втрачає ядро і перетво-рюється спочатку в ретикулоцити, а потім в еритроцити. У ретикуло-цитах, що проникають у кровоносні судини, ще деякий час зберігаєть-ся РНК у вигляді сіточки.
Гранулоцитопоез. Джерелом для розвитку зернистих лейкоцитів є також стовбурові та напівстовбурові кров'яні клітини, які одночас-но починають диференціюватися через ряд проміжних стадій у трьох різних напрямках, вони продукують клітини трьох видів: нейтрофіли, еозинофіли та базофіли. При цьому утворюються більш детерміновані клітини — попередники і моноцити.
У міру дозрівання гранулоцити зменшуються у розмірах, змінюєть-ся форма їх ядер від круглої до сегментованої, у цитоплазмі нагрома-джується специфічна зернистість. Диференціація та розмноження клі-тин цього ряду відбувається під впливом гормона гранулоцитопоетину.
Першою морфологічно розпізнаваною клітиною в напрямі грану-лоцитопоезу є мієлобласт. Експериментально доведено, що серед
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
мієлобластів є нейтрофільні, еозинофільні та базофільні мієлобласти. Клітини досягають розміру до 20 мкм, ядро у них кругле або овальне, ніжно-сітчастої будови, містить 2-5 ядерець. У цитоплазмі багато міто-хондрій, рибосом, комплекс Гольджі, лізосоми, що характеризуються позитивною реакцією на кислу фосфатазу.
Мієлобласти діляться мітозом і перетворюються у проміелоцити. Останні мають загрубілу будову ядра, їх цитоплазма характеризується появою специфічної зернистості, залежно від якої розрізняють ней-трофільні, еозинофільні та базофільні клітини. Промієлоцити діляться і перетворюються у мієлоцити.
Нейтрофільні мієлоцити розміром 12-16 мкм інтенсивно діляться мітозом. їх цитоплазма дифузно ацидофільна, містить первинні та спе-цифічні гранули, у них є усі органели. В міру дозрівання овальне ядро стає бобоподібним, кількість гетерохроматину збільшується, ядерця не виявляють. Ці клітини належать до юних форм. Вони перетворюються в метамієлоцити. Цитоплазма останніх містить збільшену кількість специфічних гранул. При подальшому дозріванні їх ядро набуває ви-гляду зігнутої палички. Подібні форми називаються паличкоядерними лейкоцитами. Потім ядро сегментується, і клітина перетворюється у сегментоядерний нейтрофіл. Повний період розвитку нейтрофільного гранулоцита становить приблизно 14 діб.
Еозинофільні мієлоцити. За характером будови ядра мало чим від-різняються від нейтрофільних мієлоцитів. Цитоплазма їх містить окси-фільну зернистість. В міру дозрівання еозинофільні мієлоцити діляться, а їх ядро набуває підковоподібної форми. Такі клітини називаються ео-зинофільними метамієлоцитами. Поступово їх ядро в середній частині набуває ниткоподібної форми, потім стає двочастковим, а у цитоплазмі збільшується кількість специфічних гранул. Клітина не здатна до поді-лу. Базофільні мієлоцити зустрічаються у меншій кількості, ніж нейтро-фільні або еозинофільні. Цитоплазма базофільних мієлоцитів містить у широко варіюючій кількості специфічні базофільні гранули різних роз-мірів, які проявляють метахромазію при фарбуванні азуром і легко роз-чиняються у воді. Найменше сегментується ядро у базофілів.
Моноцитопоез. Утворення моноцитів відбувається із стовбурових клітин червоного кісткового мозку за схемою: стовбурова кровотворна клітина, напівстовбурова кровотворна клітина, із неї утворюється уні-потентний попередник моноцитів. Наступною клітиною, яка морфо-
Розділ 3
Загальна гістологія
логічно розпізнавана є монобласт. Це значного розміру клітина, у якої базофільна цитоплазма має вигляд вузенької смужки, що оточує вели-ке ядро з двома ядерцями. монобласт ділиться мітозом, диференцію-ється у промоноцит. У останнього збільшується кількість цитоплазми, ядро набуває бобоподібної форми, втрачає здатність до мітотичного поділу, перетворюється в моноцит. Моноцити надходять у кровотік, потім проникають у тканини, де й перетворюються у незрілі, а потім зрілі макрофаги.
Лімфоцитопоез. Розвиток лімфоцитів починається із стовбурової кров'яної клітини червоного кісткового мозку, з якої утворюється клі-тина-попередник лімфоцитопоезу.
Важлива особливість цього процесу полягає у тому, що розвивають-ся морфологічно подібні, але різнорідні за функціональним значенням клітини, тобто розвиток лімфоцитів відбувається у двох напрямах - Т і В-лімфоцитів. В обох рядах виникають уніпотентні попередники, які через лімфобласти (Т і В) перетворюються у лімфоцити (Т і В). Розви-ток Т-лімфоцитів продовжується у тимусі, тут вони перетворюються в бласти, які діляться і диференціюються, перетворюючись у ефекторні клітини. У ссавців у червоному кістковому мозку утворюються В-лім-фоцити (у птахів в бурсі Фабриціуса). Тут вони диференціюються, у них з'являються імуноглобулінові рецептори. В подальшому диферен-ційовані В-лімфоцити заселяють селезінку, лімфатичні вузли, лімфа-тичні вузлики шлунково-кишкового тракту. У цих органах під впли-вом антигенів відбувається проліферація і подальша спеціалізація В-лімфоцитів з утворенням ефекторних клітин.
Розвиток кров 'яних пластинок (тромбоцитів) відбувається в чер-воному кістковому мозку і пов'язаний з розвитком у ньому гігантських клітин — мегакаріоцитів. У міру дозрівання, у результаті своєрідної ба-гаторазової ендометатичної репродукції, утворюються великі клітини (40-50 мкм), які містять у багатолопастевому ядрі 32-64 хромосомних набори. У цитоплазмі мегакаріоцитів утворюється система мікрофі-ламентів і мікротрубочок, а також специфічні гранули. У подальшому відбувається фрагментація цитоплазми мегакаріоцитів на відоремлен-ні ділянки — кров'яні пластинки, які через стінки синусоїдів червоно-го кісткового мозку потрапляють у кровоток.
Ембріональне кровотворення (розвиток крові як тканини) відбува-ється у стінці жовткового міхура, потім у печінці, червоному кістковому
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
мозку і лгмфоїдних органах (тимусі, селезінці, лімфатичних вузлах; у птахів — у фабрицієвій сумцї). Порівняно із ссавцями, у птахів добре розвинений жовтковий міхур. У мезенхімі стінки жовткового міхура відмежовуються і ущільнюються скупчення — кров'яні острівці. На їх периферії мезенхімні клітини стають плоскими, з'єднуються між собою і утворюють ендотеліальну стінку судини. Центральні клітини острів-ця округлюються, втрачають відростки: перетворюються у стовбурові клітини крові (СКК). Частина СКК диференціюється у первинні клі-тини крові — бласти. Це великі клітини з базофільною цитоплазмою і ядром з добре помітними ядерцями. Частина первинних кров'яних клітин після мітотичного ділення перетворюється у первинні еритро-бласти, що мають великий розмір і називаються мегалобластами. У їх базофільній цитоплазмі нагромаджується гемоглобін і вони перетво-рюються в поліхроматофільні, а потім в оксифільні еритробласти з ве-ликою кількістю гемоглобіну.
Деякі первинні еритробласти втрачають ядро, в інших воно збері-гається. Внаслідок цього утворюються без'ядерні та ядерні первинні еритроцити, які відрізняються від нормоцитів великими розмірами і тому їх називають мегалоцитами. Такий тип кровотворення назива-ється мегалобластичним. Він характерний для ембріонального періо-ду, але може з'являтися і після народження при деяких захворюваннях (злоякісне недокрів'я). Поряд з мегалобластичним у стінці жовтково-го міхура починається нормобластичне кровотворення, у результаті якого із бластів утворюються вторинні еритробласти; спочатку вони перетворюються у поліхроматофільні еритробласти, потім в нормо-бласти, із яких утворюються вторинні еритроцити (нормоцити). Роз-міри яких відповідають еритроцитам (нормоцитам) дорослої тварини. Розвиток еритроцитів у стінці жовткового міхура відбувається інтра-васкулярно; крім того, в стінці жовткового міхура екстраваскулярно з частини первинних клітин-бластів утворюється невелика кількість нейтрофілів та еозинофілів.
Частина СКК розноситься з кров'ю до різних органів зародка, у яких починається процес кровотворення. На п'ятому тижні ембріоге-незу великої рогатої худоби кровотворення починається у печінці, вона стає універсальним органом, де екстраваскулярно утворюються ери-троцити, зернисті лейкоцити, мегакаріоцити. В кінці ембріонального періоду розвитку кровотворення в печінці закінчується. На 7-8-му
Розділ 3
Загальна гістологія
тижні у великої рогатої худоби диференціюються лімфоцити тимуса і, мігруючи із нього, Т-лімфоцити заселяють Т-зони селезінки, лімфа-тичних вузлів. Селезінка також орган, у якому утворюються усі різно-види формених елементів крові.
На четвертому місяці ембріонального розвитку починається кро-вотворення у червоному кістковому мозку. У період ембріонального еритропоезу замінюється генерація утворення еритроцитів, які відріз-няються морфологією і типом утворення гемоглобіну. Ембріональний (фетальний) тип гемоглобіну утворюється первинними еритроцита-ми; фетальний тип гемоглобіну — еритроцитами в печінці та селезінці. Дефінітивний тип гемоглобіну утворюється еритроцитами червоного кісткового мозку.
Власне сполучні тканини
Власне сполучну тканину поділяють на: волокнисту і тканину зі спеціальними властивостями. Волокниста сполучна тканина поді-бно до інших різновидів утворена з клітин та міжклітинної речовини, обсяг якої перевищує об'єм клітин. Міжклітинна речовина утворена основною речовиною та волокнами. Залежно від цього співвідношення цю тканину поділяють на пухку і щільну.
Для сполучної тканини зі спеціальними властивостями притаман-ний переважний розвиток окремих клітинних елементів та деякі осо-бливості міжклітинної речовини. В залежності від цього її поділяють на: жирову, пігментну, ретикулярну і слизову.
Ці тканини становлять велику групу сполучних тканин зі спеці-альними властивостями. У тілі вищих хребетних вони значно поши-рені і виконують різноманітні механічні та формоутворюючі функції. їм належить важливе місце у забезпеченні реакції імунного захис-ту — фагоцитозі, утворенні імуноглобулінів та інших біологічно ак-тивних речовин. Сполучні тканини виконують трофічну і пластичну функції тощо.
Пухка сполучна тканина
Цій тканині належить важливе місце в життєдіяльності організму. Вона є у складі майже всіх органів — заповнює проміжки між ними, утворює їх оболонки та прошарки. Крім опорно-механічної фукції,
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
пухка сполучна тканина виконує захисну і трофічну функції, а також бере участь у пластичних процесах при загоюванні ран, утворенні кап-сули навколо стороннього тіла тощо.
До складу пухкої сполучної тканини входять численні клітини та
міжклітинна речовина. Усі її клітини розмежовані міжклітинною речо-
виною. Більшість з них здатні до розмноження і міграції, знаходяться в
єдиному функціональному зв'язку з клітинами судинної крові талім-
фоїднбГсигтемТн^ртанізтйу:—~~------- "~ ~~~ --------------- "~
Адвентиційні або періцити (від лат. аауептісш - прийдешній) клі-тини. Вони мають видовжену або зірчасту форму, овальне ядро, базо-фільну цитоплазму і незначну кількість органел. Ці клітини розташо-вуються на зовнішніх стінках гемокапілярів, здатні до мітозу і дифе-ренціації в інші клітини (фібробласти, міофібробласти і адіпоцити). Деякі автори заперечують існування цих клітин.
Фібробласти (від лат. пБга — волокно і гр. Біазїоз — зародок), виникають з мезенхімних клітин. У пухкій сполучній тканині вони зустрічаються частіше від інших клітин, продукують різноманітні компоненти міжклітинної речовини, синтезують і виділяють висо-комолекулярні речовини для утворення волокон основної речовини (рис. 50, 51).
В постембріональному періоді і при регенерації попередниками фібробластів є адвентиційні клітини. Залежно від їх зрілості, будови та функціонального значення фібробласти поділяють на три різно-види. Малодиферещійовані фібробласти (20-25 мкм) веретеноподіб-ної форми з незначною кількістю коротких відростків. Цитоплазма базофільна, під електронним мікроскопом в ній виявляють невелику кількість вільних рибосом, вузькі канальці гранулярної ендоплазма-тичної сітки, комплекс Гольджі, мітохондрії. Ядро овальне, має ядер-це. Малодиференційовані фібробласти здатні до активного мітозу. Вважається, що ці клітини продукують глікозаміноглікани. Зрілі фі-бробласти пластинчастої форми, з відростками досягають 50 мкм; на поперечному розрізі веретеноподібні за рахунок потовщеного ядра. В цитоплазмі фібробласта розрізняють дві зони — внутрішню ендо-плазму (більш базофільну), зовнішню ектоплазму (її забарвлення слабше). Ядро овальне, світле, багате на еухроматин і містить 1-2 ядерця. Фібробласти здатні рухатися за рахунок скоротливих біл-ків актину і міозину, які розміщені в мікрофіламентах ектоплазми.
 Розділ 3 Загальна гістологія
Розділ 3 Загальна гістологія
Цитоплазма зрілих фібробластів містить добре розвинену грануляр-ну ендоплазматичну сітку, комплекс Гольджі у вигляді цистерн і мі-хурців, тут синтезуються глікозаміноглікани, різної форми і розміру мітохондрії, лізосоми. Фібробласти, крім глікозаміногліканів синте-зують білки, проколаген, проеластин, ферментні білки, а також фі-брилярний глікопротеїд фібронектин, який впливає на рух клітин і зв'язування їх з мікрооточенням.

Рис. 50. Пухка сполучна тканина:
1-фібробласт; 2-макрофаг; 3-тканинний базофіл; 4-колагенові волокна; 5-еластичні волокна.
Фіброцити — веретеноподібні клітини з видовженим ядром, зна-ходяться між волокнами сполучної тканини. Фіброцити містять не-значну кількість органел, нездатні до розмноження, характеризуються зниженими синтетичними процесами.
Міофібробласти. Функціональну дію проявляють у грануляційній тканині; містять розвинену ендоплазматичну сітку та скоротливі фі-брили. Ці клітини є похідними фібробластів.
| В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія |
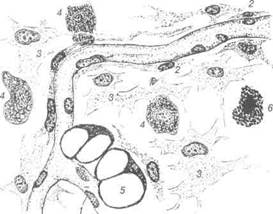
Рис. 51. Пухка сполучна тканина із підшкірної клітковини:
1-ендотелій капіляра; 2-камбіальна клітина; 3-фібробласт; 4-макрофаг; 5-жи-рова клітина; 6-тканинний базофіл.
Гістіоцити - макрофаги пухкої сполучної тканини, котрі дифе-ренціюються з моноцитів крові. Вони розміщуюються поодиноко або групами. Розмір їх 12-18 мкм, здатні до міграції, форма кругла, витяг-нута. Цитоплазма базофільно-плямиста, містить помірні мітохондрії і комплекс Гольджі, гранулярна ендоплазматична сітка майже не роз-винена. Багато в них лізосом та фагосом, за їх допомогою перетравлю-ються фагоцитовані речовини. Ядро невелике, овальне, містить багато гетерохроматину, інтенсивно забарвлюється. Під електронним мікро-скопом в гістіоцитах виявлені псевдоподії, пальцеподібні вирости, мі-кроворсинки, що відходять від їх поверхні. За їх допомогою макрофаги захоплюють макромолекулярні частинки, мікроорганізми, віруси, про-дукти клітинного розпаду. На поверхні плазмолеми-истюцита містять-ся рецептори для пухлинних клітин, еритроцитів, Т- ї В-лімфоцитів, імуноглобулінів тощо. Наявність рецепторів макрофагів забезпечує їх участь в імунних реакціях. Попередниками гістіоцитів є моноцити периферійної крові. Моноцити, які мігрують із судин, у відповідних середовищах адаптуються і перетворюються у органо- і тканинноспе-цифічні макрофаги, утворюють макрофагічну систему.
Розділ 3
Загальна гістологія
Макрофаги — багатофункціональні клітини. їм належить важли-ве місце у формуванні природного і набутого імунітету. У природному імунітеті участь макрофагів проявляється в здатності до фагоцитозу і піноцитозу, Фагоцитозний матеріал всередині макрофага піддаєть-ся ферментативному розщепленню за допомогою лізосом. Так лікві-дуються шкідливі для організму речовини. Макрофаги синтезують активні речовини — лізоцим, інтерферон, пірогени тощо. В набутому імунітеті їх значення проявляється в пізнанні антигена, його перетво-ренні з корпускулярної форми в молекулярну, передачі імунокомпе-тентним клітинам і виконання ефекторної функцп' при взаємодії з Т-і В-лімфоцитами.
Окрім гістіоцитів до макрофагів належать зірчасті клітини Куп-фера (печінки), вільні та фіксовані макрофаги лімфовузлів, селезінки, червоного кісткового мозку, альвеолярні макрофаги легень, плевральні та перитонєальні макрофаги, остеокласти, клітини Лангерганса (шкі-ра), клітини мікроглії (нервова тканина), макрофаги синовіальних обо-лонок суглобів та інших органів.
Тканинні базофіли (мастоцити, лаброцити, тучні клітини) похо-дять від стовбурової кровотворної клітини (СКК).
Термін «тучні клітини» вперше запропонував Ерліх в 1877 р. для позначення клітин, які були насичені гранулами. Кількість тканинних базофілів і базофілів крові деяких тварин має зворотню залежність. Тканинні базофіли є у дрібних кровоносних судинах сполучної ткани-ни органів травлення, молочної залози, шкіри, тимуса, матки, яйцепро-воду птахів. За формою вони округлі, овальні, веретеноподібні тощо. Розмір тканинних базофілів 10-25 мкм і більше. У птахів вони значно менших розмірів, ніж у ссавців. Ядро цих клітин у більшості випадків оточене гранулами, кругле, займає центральне положення, має звичай-ну будову. Серед органел тканинні базофіли містять добре розвинений комплекс Гольджі, велику кількість мітохондрій, небагато гранулярної та агранулярної ендоплазматичної сітки.
Головною складовою частиною тканинних базофілів є велика кількість базофільних гранул (0,3-1 мкм), рівномірно розміщених в цитоплазмі. Вони характеризуються метахромазією, зумовленою сульфатованим глікозаміногліканом. Гранули тканинних базофілів містять ряд біологічно активних речовин, серед яких гепарин стано-вить 30% вмісту гранул, а також гістамін, хондроітинсульфат, дофамін,
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
серотонін. Гепарин знижує проникність міжклітинної речовини, запо-бігає зсіданню крові. Гістамін викликає скорочення міоцитів, сприяє виходу плазми з венул і капілярів за рахунок розширення і підвищен-ня проникності їх стінки. Гістамін стимулює міграцію еозинофілів, ак-тивізує макрофаги. Тканинні базофіли одні із перших реагують на про-никнення антигенів клітин із крові. На плазмолемі знаходиться зна-чна кількість імуноглобулінів, що належать до класу ]$Є. Зв'язування антигенів і утворення комплексу антиген — антитіло супроводжується дегрануляцією і виділенням із тканинних базофілів судинноактивних речовин, які зумовлюють появу місцевих і загальних реакцій.
Плазматичні клітини (плазмоцити). Займають фундаментальне місце в імунних реакціях. Плазматичні клітини забезпечують утворен-ня антитіл, що циркулюють в крові. Ці антитіла називаються гумораль-ними (від лат. пшпог — рідина). Плазматичні клітини високоспеціалі-зовані. Вони синтезують і виділяють основну масу імуноглобулінів. Походять плазматичні клітини з В-лімфоцитів під впливом антигенів. Антиген (від гр. апіі — проти, §епоп — утворювати) означає речовину, проти якої утворюється інша речовина.
Значна частина плазматичних клітин знаходиться під епітелієм органів дихання, кишечника, у лімфатичних вузлах, селезінці, мигда-лику, сполучній тканині слізної залози. За формою вони більш-менш округлі, розміром до 12 мкм.
Цитоплазма, порівняно з об'ємом ядра, займає значно більшу пло-щу. Ядро округле, міститься ексцентрично. Значна частина хроматину знаходиться у конденсованому стані; хроматин часто розміщується подібно ци-фрам на циферблаті годинника. Цитоплазма інтенсивно базофільна, на її фоні біля ядра помітне «світле подвір'я», де знахо-диться комплекс Гольджі. Базофілія цитоплазми зумовлена великою кількістю РНК. Цитоплазма плазмоцитів спеціалізована для біосинте-зу білкового секрету, який знаходиться в добре розвинених цистернах гранулярної ендоплазматичної сітки. Синтезовані молекули імуно-глобулінів надходять у просвіт цистерни, а потім в комплекс Гольджі, звідки після приєднання вуглеводного компонента виводяться на по-верхню клітини. Інші деталі будови плазматичної клітини звичайні, за винятком того, що плазматична мембрана цих клітин часто утворює пальцеподібні відростки. Плазматичні клітини не здатні до поділу, вони старіють, гинуть і поглинаються макрофагами.
Розділ 3
Загальна гістологія
Адіпоцити (від лат. ааерз — жир) - це клітини здатні накопичувати в своїй цитоплазмі резервний жир і мають кулясту форму. їх розміри залежать від кількості жиру і досягають 120 мкм. Жир з'являється в них спочатку у вигляді дрібних капель, котрі зливаються в одну велику кра-плю, яка заповнює всю клітину. Внаслідок цього ядро розміщується під плазмолемою. Такі клітини на поперечному розрізі набувають форму «каблучки».
Значне накопичення жиру інколи призводить до розриву плазмо-леми клітини і жир виходить в міжклітинну речовину. При електрон-ній мікроскопїт в цих клітинах спостерігається майже повна відсутність ендоплазматичної сітки, слабо розвинений комплекс Гольджі. Проте в клітинах бурого жиру ці органели розвинені значно краще.
Пігментоцити - ці клітини в нормальній сполучній тканині ссав-ців трапляються рідко. Вони знаходяться переважно в стінці очного яблука (в склері, судинній та райдужній оболонках).
В окремих ділянках шкіри (вим'я, мошонка) кількість пігменто-цитів значна. Ці клітини мають зірчасту форму і зерна пігменту - ме-ланіну. Одні з цих клітин здатні синтезувати і накопичувати пігмент, інщі^- лише накопичувати.
У нижчих хребетних пігментні клітини (хроматофори) спостері-гаються постійно. Мають відросчасту, неправильну форму. Під регу-люючим впливом нервової системи хроматофори можуть активно ско-рочуватися і внаслідок чого змінюється колір шкіряного покрову, що робить тварину малопомітною.