




Найбільш примітивні з сучасних ссавців — качконіс, єхидна, ха-рактеризуються овоцитами, які містять велику кількість жовтка, нале-жать до телолецитального типу, зигота дробиться за меробластичним методом — дискоїдально. Іх ембріогенез подібний до розвитку птахів.
Розділ 2
Ембріологія
Зигота качконоса розвивається поза материнським організмом. У єхидни зигота розвивається в складці шкіри на черевній поверхні тіла, але живлення від матері зародок не одержує.
У зиготі сумчастих жовтка мало, в процесі дроблення він навіть відторгується (сумчаста куниця, опосум).
У деяких сумчастих розвивається примітивна плацента, яка функ-ціонує нетривалий час. Зародок цих представників ссавців народжу-ється недорозвиненим, занадто маленьким (1-2 см) і доношується ма-тір'ю в сумці, де встановлюється зв'язок стравоходу зародка з соском матері, її молоко час від часу впорскується в стравохід скороченням м'язів молочної залози.
Ссавці різняться між собою не лише будовою яйцеклітин та умова-ми ембріонального розвитку, а й іншими ознаками: дроблення зиготи, початком імплантації зародка, рівнем зрілості при народженні, трива-лістю внутрішньоутробного розвитку тощо.
Вторинна поява в процесі еволюції оліголецитальних яйцеклітин у вищих ссавців зумовлена переходом їх зародка на живлення за рахунок організму матері. Зародкові оболонки і маткові тканини вступають в тісну взаємодію, утворюється плацента, яка безпосередньо постачає все необхідне для розвитку і виводить продукти обміну 3 розвитком пла-центи зникає необхідність у жовтку, однак у вищих ссавців жовтковий мішок утворюється, хоча і не виконує функції, пов'язаної з живленням.
Дроблення (рис. 34). В результаті запліднення яйцеклітина перехо-дить із інгібірувального стану в стан надзвичайно інтенсивного метабо-лізму, що характеризується різким підвищенням дихальної і синтетичної активності. Дроблення оліголецитальних зигот ссавців повне, асинхрон-не, нерівномірне, відбувається відносно повільно. В той час, коли зигота рухається у матковій трубі, утворюється зародок, який складається із не-парної кількості бластомерів, що мають вигляд купки клітин. Цю стадію називають морулою (від лат. тошіа - тута). В кінці дроблення зародок складається із купки темних великих клітин — зародкового вузлика (емб-ріобласта) і поверхневого шару світлих дрібних клітин — трофобласта. За рахунок ембріобласта розвивається тіло зародка, а трофобласт вико-нує трофічну функцію, за його участю забезпечується зв'язок із стінкою матки. У трофобласті розвиваються численні вирости — первинні вор-синки, утворені тільки трофобластичними розростаннями, в які потім вростає позазародкова мезодерма (мезенхіма) і утворює там кровоносні
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
судини. Ворсинки трофобласта з кровоносними судинами є вторинними, вони мають мезодермальну основу, а трофобласт з вторинними ворсин-ками називають хоріоном. Третинні ворсинки містять кровоносні суди-ни. Якщо тварина багатоплідна (свині, собаки, кролі тощо), то у маткові труби надходять одночасно кілька яйцеклітин. Під час дроблення заро-док переміщується у матковій трубі у напрямку матки (рис. 35).
Рис. 34. Електронна мікрофотографія чотириденного зародка хом'яка х 700; на поверхні клітини трофобласта
У різних представників вищих ссавців період дроблення зиготи буває різної тривалості. Так, у кролематок через три доби після пару-вання зародок знаходиться на стадії бластули.
Дроблення зигот у свині подібне з процесом у кролематок. В овець дроблення зиготи закінчується за 3-4 доби, у корів — за 6 діб, у прима-тів дроблення триває з 1 по 6 добу життя зародка. Спочатку трофобласт всмоктує секрет залоз матки «маточне молоко», зародок збільшується в об'ємі і перетворюється у бластодермічний міхурець, який називаєть-ся бластоцистою. Під час дроблення і утворення бластоцисти прозора зона зиготи ще зберігається. В подальшому за допомогою ворсинок, що утворилися на поверхні трофобласта, бластоциста імплантується (від лат. іт — проникнення в, ріапїаїіз — посадка), тобто прикріплюється до слизової оболонки матки. Встановлення тісного взаємозв'язку із слизовою оболонкою матки — перший критичний період у розвитку за-родка. У ссавців різних видів стінка трофобласта неодночасно вступає в тісний контакт з слизовою оболонкою матки. Так, у великої рогатої
іділ2
Ембріологія
худоби імплантація настає на 17 добу, у кобили — на 63-70 добу, у ма-каки — на 9 день після запліднення, у людини — на 7 добу В цей час під впливом ферментних систем трофобласта руйнується оболонка за-пліднення, яка до цього часу оточувала зародок і бластоциста вступає в контакт із слизовою оболонкою матки (рис. 36).
В Г
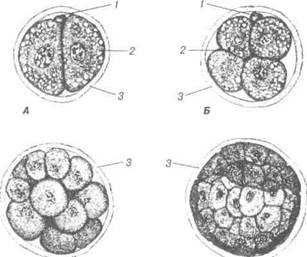
Рис. 35. Дроблення зиготи свині х 400:
А — стадія двох бластомерів; Б — стадія чотирьох бластомерів; В — морула; Г — бластула, світліша центральна ділянка, в якій формується бластоцель шляхом перерозподілу клітин: 1- полярне тільце; 2 — жирові вакуолі; 3 — прозора оболонка
Клітини зародкового вузлика формують зародковий диск, анало-гічний такому у птахів. У центрі зародкового диска утворюється зарод-ковий щиток, із його матеріалу розвивається тіло зародка, а із остан-ньої частини — провізорні органи. Процес гаструляції у сумчастих і плацентарних ссавців принципово такий, як у плазунів і птахів.
Шляхом делямінації зародкового диска утворюється ектодерма і ентодерма. Зародкові листки, що утворилися з матеріалу зародкового щитка, називаються зародковими, а ті, що виникли із позазародкової зони бластодиска — позазародковими.
| В.П. Новак, Ю.П. Бичков, М.Ю. Пилипежо Цитологія, гістологія, ембріологія |
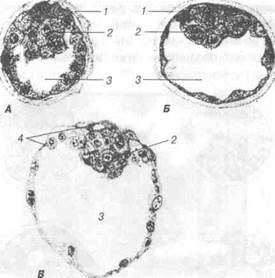
Рис. 36. Три стадії бластодермного міхурця свині х 375:
А — через 114 год. після парування; Б — через 145,75 год.; В — через 164 год. після парування; 1 — прозора зона; 2 — внутрішня клітинна маса; 3 — бласто-цель; 4 — трофобласт.
Розвиток зародка ссавців пов'язаний з утворенням первинної смужки, яка подібна до первинної смужки птахів. Від переднього кінця первинної смужки, клітини переміщуються і утворюють головний від-росток. Останній разом із первинною смужкою дає початок мезодер-мі, головний відросток і його мезодерма відповідають хордомезодермі нижчих тварин. Диференціювання зародкових листків відбувається так, як у інших тварин. Ці ознаки є найдавнішими і відображають ево-люцію, якій піддавалися у своєму розвитку ссавці.
Необхідно зазначити, що однією з особливостей розвитку ссавців при ізолецитальному типі овоцитів і голобластичному дробленні зиго-ти вважають утворення тимчасових органів. Друга особливість розви-тку ссавців — це раннє відокремлення зародкової частини від позаза-родкової. На початку дроблення формується трофобласт, за допомогою якого зародок одержує поживні речовини з материнського організму.
Розділ 2
Ембріологія
Після утворення зародкових листків трофобласт, що знаходиться над зародком, тимчасово розвивається і до утворення амніона знахо-диться на поверхні. Внаслідок розростання ектодерми зародкового листка останній починає вигинатися, утворює амніотичні складки, які потім зростаються в загальний амніон. Зовнішні листки складок, які прямо переходять у трофобласт, також з'єднуються над зародком в одну оболонку, яка нагадує серозу птахів. До цієї оболонки зсередини підростає мезодерма паріетального листка спланхнотома, в якій роз-виваються кровоносні судини. Так виникає хоріон (від лат. спогіоп — шкіра). Слід зазначити, що у ссавців (як і у птахів) розвитку зарод-кових оболонок передує утворення тулубової та амніотичної складок. Тулубова складка піднімає зародок над жовтковим мішком і відокрем-лює його зародкову частину від незародкової, а зародкова ентодерма замикається в первинну кишку, проте остання залишається з'єднаною з жовтковим мішком вузьким жовтковим протоком.
Жовтковий мішок, як і у птахів, утворюється із позазародкової енто-дерми та вісцерального листка мезодерми. На відміну від птахів, він міс-тить не жовток, а білкову рідину. В стінці жовткового мішка утворюють-ся кровоносні судини, він виконує кровотворну та трофічну функції.
Амніон (водна оболонка) розвивається з внутрішньої частини ам-ніотичної складки, а хоріон — із зовнішньої. Амніон у ссавців виконує функцію, подібну до такої у птахів. Хоріон гомологічний серозній обо-лонці птахів. На його поверхні утворюються ворсинки, які вростають в стінку матки. Ця зона дуже потовщена, у ній багато кровоносних судин і її називають плацентпою. Надходження поживних речовин у кров за-родка та вихід із неї відбувається дифузним шляхом або за допомогою активного перенесення, тобто з витратами на цей процес енергії. Кров матері не змішується з кров'ю зародка.
Ентодерма задньої частини кишечника утворює вип'ячування — алантоїс. Так, як у ссавців сечовина із тіла зародка виділяється через плаценту в кров матері, але, на відміну від птахів, алантоїс у них не є сечовим мішком, однак його проксимальна частина пізніше бере участь в утворенні сечового міхура. Його мезодерма дає початок кровоносним судинам, які підростають до хоріона.
Поясна плацента характерна для хижаків. Ворсинки хоріона зна-ходяться в середній частині плодового міхура і утворюють на його по-верхні ніби поясок (рис. 38).
7 - 8-305
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
Й'.№.
 * ■ Ч' 'і» • •'' -'''
* ■ Ч' 'і» • •'' -'''
■?Л:
Рис. 37. Котиледонна плацента жуйних:

|
а — хоріон, б — амніон, в — котиледон

 Рис. 38. Поясковидна плацента хижаків
Рис. 38. Поясковидна плацента хижаків
Оскільки зародок починає живитися за рахунок материнського ор-ганізму, у багатьох ссавців жовтковий мішок не відіграє значної ролі, як у птахів, хоча жовтковий мішок утворюється, що характерно для ембріонів свиней, кролів, коней. В жовтковому мішку утворюються кровоносні судини, які забезпечують зародок поживними речовина-ми матки. Пізніше, коли алантоїс досягає значного розвитку і набуває функції жовткового мішка, останній поступово редукується.
Амніон виникає у вигляді шару соматоплеври, яка утворює навколо зародка складки таким чином, як це було описано для курячого зародка, проте у ссавців частина трофобласта, що знаходиться над зародковим диском, тимчасово розривається і зародок до утворення амніона залиша-ється відкритим. Коли амніон замикається, він заповнюється рідиною,
Розділ 2
Ембріологія
яку продукує. В амніотичній рідині зародок знаходиться у зваженому стані. При цьому зародок зазнає однакового тиску зовні і зсередини, що запобігає механічним пошкодженням. Крім того, амніотична оболонка забезпечує розвиток плода в оптимальному за складом електролітів та інших речовин водному середовищі. Амніон прикріплюється до тіла за-родка у тому місці, де на черевній поверхні в ділянці жовткового сте-бельця стінка тіла відкрита, в подальшому тут утворюється пуповина.
Хоріоном є зовнішній шар бластоцисти ссавців. На різних стадіях розвитку його називають по-різному: на стадії внутрішньої клітинної маси — трофобластом, а після утворення ентодерми та мезодерми зо-внішній шар бластоцисти продовжується в ектодерму зародка, й тому має трофоектодерму, оскільки частина цього шару нагромаджує по-живні речовини матки. Після того, як мезодерма розшарується і її па-ріетальний шар з'єднується з ектодермою, цей шар, який став тепер позазародковою соматоплеврою, називають хоріоном. Про утворення алантоїса та його значення було описано вище.
Плаценти
Плацента (від гр. ріасез — коржик) цей позазародковий орган отримав таку назву тому що у приматів, в тому числі у людей він має форму коржика або диску.
Плацента розвивається пізніше вище вказаних провізорних орга-нів і функціонує до кінця вагітності. Вона забезпечує зародок пожив-ними речовинами, киснем та виводить продукти обміну В плаценті синтезуються такі гормони: прогестрон, естрогени, плацентарний лак-тоген, хоріонічний гонадотропін та інші біологічно активні речовини необхідні для нормального перебігу вагітності.
Плацента утворена з двох частин: зародкової (плодової) і материн-ської. Зародкову частину формує алантохоріон (у жуйних) і амніо-хоріон (у хижаків), а материнська частина представлена ендометрієм (слизовою оболонкою матки).
Залежно від розміщення ворсинок хоріону на ендометрії розрізня-ють чотири типи плацент: дифузну, котиледонну, кільцеподібну та дис-коподібну.
Після короткочасного живлення зародка запасом жовтка наявного в зиготі, встановлюється живлення крізь трофобласт секретом залоз
7*
В.П. Новак, Ю.П. Бичков, М.Ю. Пилипенко Цитологія, гістологія, ембріологія
матки - «маточним молоком». Потім розвивається гемотрофний спо-сіб живлення крізь кровоносні судини. Останній спосіб має місце тіль-ки у плацентарних ссавців.
Існує декілька типів класифікації плацент. Один з них оснований на принципі відходження або не відходження материнської плаценти від матки при народженні плода. Другий тип класифікації грунтується головним чином на формі плацент. Третій тип класифікації грунтуєть-ся на складі тканинного бар'єру між кровоносними судинами матері і плода. На підставі різниць в ступені стикання судин материнської і плодної частин плацент останні поділяються на дві групи: напівпла-центи (три типи) та справжні плаценти (два типи).
Перший тип напівплацент - дифузний і епітеліохоріальний (коби-ла, свиня, ослиця, верблюдиця). Кров зародка відокремлена від крові матері шарами епітелію і сполучної тканини. Ворсинки хоріону зану-рюються в маткові залози і контактують з епітелієм.
Другий тип напівплацент - котиледонний і десмохоріальний (жуй-ні). Ворсинки хоріону розміщуюються групами - котиледонами, які руйнують епітелій ендометрію і контактують з сполучною тканиною ен-дометрія, котрий утворює відповідні випинання карункули (рис. 37)
Третій тип напівплацент - кільцеподібний і ендотеліохоріальний. Трофобласт хоріона руйнує сполучну тканину і контактує з ендотелі-єм кровоносних капілярів. Ця плацента властива хижакам.
Четвертий тип справжніх плацент - дископодібний та гемохорі-альний (гризуни, примати). Судини ендометрія частково утворюють лакуни, ворсинки хоріона омиваються материнською кров'ю (рис. 38)
П'ятий тип справжніх плацент - гемоендотеліохоріальний (деякі гризуни). Поміж кров'ю плода і кров'ю матері єдиною перепоною за-лишається ендотелій капілярів плода. Ці капіляри занурюються в си-нуси, де циркулює кров матері.

