




Черепные нервы (nn. craniales) так же, как и спинномозговые нервы, относятся к периферическому отделу нервной системы. Различие заключается в том, что спинномозговые нервы отходят от спинного мозга, а черепные — от головного мозга, причем 10 пар черепных нервов начинаются от ствола мозга; это — глазодвигательный (III), блоковый (IV), тройничный (V), отводящий (VI), лицевой (VII), преддверно-улитковый (VIII), языкоглоточный (IX), блуждающий (X), добавочный (XI), подъязычный (XII) нервы; все они имеют различное функциональное значение (рис. 67). Еще две пары нервов — обонятельный (I) и зрительный (II) — не являются типичными нервами: они формируются как выросты стенки переднего мозгового пузыря, имеют необычное по сравнению с другими нервами строение и связаны с проведением специализированных видов чувствительности.
По общему плану строения черепные нервы сходны со спинномозговыми нервами, однако имеют и определенные отличия. Как и спинномозговые нервы, они могут состоять из волокон разного типа: чувствительных, двигательных и вегетативных. Однако некоторые черепные нервы включают только афферентные или только эфферентные волокна. Для части черепных нервов, связанных с жаберным аппаратом, присущи некоторые внешние признаки метамерии (рис. 68). Общий состав волокон черепного нерва практически соответствует составу его ядер в стволе мозга. Сенсорные афферентные волокна, как правило, берут начало от нейронов, расположенных в чувствительных ганглиях. Центральный отросток каждого из этих нейронов проникает в ствол в составе черепного нерва и заканчивается в соответствующем чувствительном ядре. Двигательные и вегетативные эфферентные волокна отходят от групп нейронов, расположенных в соответствующих черепному нерву двигательных и вегетативных ядрах (см. рис. 55, 63).
В формировании черепных нервов прослеживаются те же закономерности, что и при образовании спинномозговых нервов:
• двигательные ядра и двигательные волокна являются производными
базальной пластинки нервной трубки;
• чувствительные ядра и чувствительные нервы образуются из нервно
го гребня (ганглионарной пластинки);
• вставочные нейроны (интернейроны), обеспечивающие связи между
разными группами ядер черепных нервов (чувствительными, двига
тельными и вегетативными), формируются из крыльной пластинки
нервной трубки;


Рис. 67. Места выхода из головного мозга 12 пар черепных нервов и их функции.


Рис. 68. Закладка черепных нервов у эмбриона 5 недель.
• вегетативные ядра и вегетативные (преганглионарные) волокна закладываются в межуточной зоне между крыльной и базальной пластинками.
В расположении ядер черепных нервов наблюдаются и специфические, свойственные только им особенности, обусловленные характером формирования ствола мозга. В процессе его развития происходит увеличение и видоизменение крыши нервной трубки на уровне всех отделов ствола мозга, а также смещение материала крыльных пластинок в вентролатеральном направлении. Эти процессы приводят к тому, что ядра черепных нервов смещаются в покрышку ствола мозга. При этом двигательные ядра III—XII пар черепных нервов занимают наиболее медиальное положение, чувствительные — наиболее латеральное, а вегетативные — промежуточное. Это хорошо видно на их проекции на дно ромбовидной ямки (см. рис. 63).
Все черепные нервы, за исключением блуждающего (X пара), иннервиру-ют только органы головы и шеи. Блуждающий нерв, в состав которого включены парасимпатические преганглионарные волокна, участвует также в иннервации почти всех органов грудной и брюшной полостей. С учетом функциональных особенностей, а также специфики развития все черепные нервы можно разделить на следующие основные группы: сенсорные (связанные с органами чувств), соматомоторные, соматосенсорные и бранхиогенные (табл. 4).
Сенсорные, или нервы органов чувств (I, II и VIII пары), обеспечивают проведение в центральную нервную систему импульсов специфической чув-
Таблица 4. Черепные нервы и области их иннервации
| Функциональные группы нервов | Черепные нервы | Состав нервных волокон | Область иннервации |
| Сенсорные | I— обонятельный (n. olfactorius) | Отростки вставочных нейронов | Орган обоняния |
| II — зрительный (n. opticus) | Орган зрения | ||
| VIII — преддверно-улитковый (n. vestibulocochlearis) | Чувствительные | Орган слуха, орган равновесия | |
| Соматомоторные | III — глазодвигательный (n. oculomotorius) | Двигательные Вегетативные | Глазные мышцы, внутренние мышцы глаза |
| IV — блоковый (n. trochJearis) | Двигательные | Глазные мышцы | |
| VI — отводящий (n. abducens) | Двигательные | ||
| XII — подъязычный (n. hypoglossus) | Двигательные | Мышцы языка | |
| Соматосенсорные | V1— тройничный (n. trigeminus) | Чувствительные | Кожа головы (верхняя часть лица), роговица; проприоцептивная иннервация глазных мышц |
| Бранхиогенные | V2,3 — тройничный (n. trigeminus) | Чувствительные Двигательные | Кожа головы (средняя и нижняя части лица); слизистые оболочки полости носа и рта; зубы; язык; проприоцептивная иннервация мышц языка; жевательные мышцы |
| VII — лицевой (n. facialis) | Чувствительные Двигательные Вегетативные | Слизистая оболочка полости рта; язык (вкусовая); мимические мышцы лица; слезная и слюнные железы; железы слизистых оболочек полостей рта и носа | |
| IX— языкоглоточный (n. glossopharyngeus) | Чувствительные Двигательные Вегетативные | Слизистые оболочки зева и глотки, глоточные миндалины; язык (вкусовая); мышцы глотки; околоушная слюнная железа; миндалина; железы слизистой оболочки глотки | |
| X — блуждающий (n. vagus) | Чувствительные Двигательные Вегетативные | Сердце, легкие, гортань, органы желудочно-кишечного тракта (от корня языка до нисходящей ободочной кишки); мышцы гортани и глотки, мышцы, опускающие мягкое нёбо | |
| XI — добавочный (n. accessories) | Двигательные | Трапециевидная и грудино-ключично-сосце-видная мышцы |
 ствительности от органов чувств (обоняния, зрения и слуха). Они содержат только чувствительные волокна, как, например, VIII пара черепных нервов, которые берут начало от нейронов, расположенных в сенсорном ганглии (спиральный узел). Нервы I и IIпары представляют собой фрагменты проводящего пути обонятельного и зрительного анализаторов.
ствительности от органов чувств (обоняния, зрения и слуха). Они содержат только чувствительные волокна, как, например, VIII пара черепных нервов, которые берут начало от нейронов, расположенных в сенсорном ганглии (спиральный узел). Нервы I и IIпары представляют собой фрагменты проводящего пути обонятельного и зрительного анализаторов.
С обонятельным нервом связаны два маленьких терминальных нерва (п. terminalis), обозначенных как 0 (нулевая) пара черепных нервов. Терминальный, или концевой, нерв был открыт у низших позвоночных, но обнаруживается также у человека. Он содержит в основном немиелинизиро-ванные нервные волокна, отходящие от биполярных или мультиполярных нейронов, собранных в небольшие группы, локализация которых у человека неизвестна. Связи нейронов, образующих ядро терминального нерва, также неизвестны. Каждый нерв располагается медиальнее обонятельного тракта, а его ветви, как и обонятельные нервы, проходят через решетчатую пластинку в основании черепа и оканчиваются в слизистой оболочке носовой полости.
В функциональном плане терминальный нерв является сенсорным и есть основание думать, что он служит для обнаружения и восприятия феромонов — пахучих веществ, выделяемых для привлечения существ противоположного пола (подробнее о сенсорных нервах см. гл. 6).
К соматосенсорным нервам относится верхняя (или первая) ветвь тройничного нерва (V1), поскольку она содержит только чувствительные волокна нейронов сенсорного узла тройничного нерва, проводящие импульсы, вызванные тактильными, болевыми и температурными раздражениями кожи верхней трети лица, а также проприоцептивными раздражениями глазодвигательных мышц.
Соматомоторные, или двигательные, черепные нервы (III, IV, VI, XII пары) иннервируют мышцы головы. Все они образованы длинными отростками мотонейронов, расположенных в двигательных ядрах ствола.
Глазодвигательный нерв (п. oculomotorius) — III пара; оба нерва (правый и левый) имеют 5 ядер: двигательное ядро глазодвигательного нерва (парное), добавочное ядро (парное) и срединное ядро (непарное). Срединное и добавочные ядра являются вегетативными (парасимпатическими). Эти ядра расположены в покрышке среднего мозга под водопроводом мозга на уровне верхних холмиков четверохолмия.
Двигательные волокна глазодвигательного нерва после выхода из ядер частично перекрещиваются в покрышке среднего мозга. Затем глазодвигательный нерв, включающий двигательные и парасимпатические волокна, покидает ствол мозга с медиальной стороны ножек мозга и через верхнюю глазничную щель проникает в глазницу. Он иннервирует глазодвигательные мышцы (верхнюю, нижнюю, медиальную прямые и нижнюю косую мышцы глаза), а также мышцу, поднимающую верхнее веко (рис. 69).
Парасимпатические волокна глазодвигательного нерва прерываются в ресничном узле, лежащем в глазнице. От него постганглионарные волокна направляются к глазному яблоку и иннервируют ресничную мышцу, сокращения которой изменяют кривизну хрусталика глаза, и сфинктер зрачка.

Рис. 69. Глазодвигательный, блоковый и отводящий нервы (III, IV и VI пары), иннервирующие мышцы глаза. А. Ствол головного мозга. Б. Глазное яблоко и глазодвигательные мышцы.
Ядра глазодвигательного нерва получают афферентные волокна в основном от медиального продольного пучка (обеспечивающего согласованную работу ядер черепных нервов, которые управляют движением глаз, а также их связь с вестибулярными ядрами), от ядер верхних холмиков пластинки крыши среднего мозга и ряд других волокон.
Благодаря связям ядер глазодвигательного нерва с корой больших полушарий возможны не только непроизвольные (автоматизированные, машинальные), но и произвольные (сознательные, целенаправленные) движения глазного яблока.
Блоковый нерв (n. trochlearis) — IV пара — относится к группе глазодвигательных нервов. Он берет начало от нейронов парного двигательного ядра блокового нерва, расположенного в покрышке среднего мозга под дном водопровода мозга на уровне нижних холмиков четверохолмия.
Волокна блокового нерва выходят из ядер в дорсальном направлении, огибают сверху водопровод мозга, входят в верхний мозговой парус, где образуют перекрест и выходят из ствола мозга на дорсальной его поверхности. Далее нерв огибает ножку мозга с латеральной стороны и направляется вниз и вперед. В глазницу он проникает вместе с глазодвигательным нервом через глазничную щель. Здесь блоковый нерв иннервирует верхнюю косую мышцу глаза, поворачивающую глазное яблоко вниз и латерально (см. рис. 69).
 Отводящий нерв (n. abducens) — VI пара — также относится к глазодвигательной группе нервов. Он берет начало от нейронов парного двигательного ядра отводящего нерва, расположенного в покрышке моста. Двигательные волокна отводящего нерва выходят из ствола мозга между мостом и пирамидой продолговатого мозга. Направляясь вперед, нерв проникает в глазницу через верхнюю глазничную щель. Иннервирует наружную прямую мышцу глаза, поворачивающую глазное яблоко наружу (см. рис. 69).
Отводящий нерв (n. abducens) — VI пара — также относится к глазодвигательной группе нервов. Он берет начало от нейронов парного двигательного ядра отводящего нерва, расположенного в покрышке моста. Двигательные волокна отводящего нерва выходят из ствола мозга между мостом и пирамидой продолговатого мозга. Направляясь вперед, нерв проникает в глазницу через верхнюю глазничную щель. Иннервирует наружную прямую мышцу глаза, поворачивающую глазное яблоко наружу (см. рис. 69).
Подъязычный нерв (n. hypoglossus) — XII пара — берет начало в парном двигательном ядре подъязычного нерва, расположенном в покрышке продолговатого мозга. Ядро проецируется на дно ромбовидной ямки в области ее нижнего угла в треугольнике подъязычного нерва. Ядро продолжается в спинной мозг до шейных сегментов (Q_n).
Волокна подъязычного нерва в виде нескольких корешков покидают продолговатый мозг между пирамидой и оливой. Корешки сливаются в общий ствол, который выходит из полости черепа через канал подъязычного нерва. Этот нерв иннервирует мышцы языка.
Бранхиогенные, или жаберные, нервы (V2,3, VII, IX, X, XI пары) представляют собой группу наиболее сложно устроенных черепных нервов. Исторически они развивались в связи с процессом закладки жаберных дуг. Именно эта группа нервов имеет признаки метамерии: V2,3 пара — нерв I висцеральной (челюстной) дуги; VII пара — нерв II висцеральной (подъязычной) дуги; IX пара — нерв III висцеральной (I жаберной) дуги; X пара — нерв II и последующих жаберных дуг. XI пара в процессе своего развития отделилась от X пары черепных нервов.
Тройничный нерв (n. trigeminus) — V пара. Это один из наиболее сложно устроенных нервов, так как фактически он объединяет в себе два нерва: V1 — соматосенсорный нерв головы и V2,3 — нерв I висцеральной (челюстной) дуги. На основании мозга тройничный нерв появляется из толщи средних ножек мозжечка в виде толстого и короткого стволика, состоящего из двух корешков: чувствительного и двигательного. Двигательный корешок нерва более тонкий. Он передает двигательные импульсы к жевательным и некоторым другим мышцам. Чувствительный корешок в области вершины пирамиды височной кости образует утолщение полулунной формы — тройничный узел. Он, как и все сенсорные ганглии, состоит из псевдоуниполярных нейронов, центральные отростки которых направляются к сенсорным ядрам тройничного нерва, а периферические идут в составе трех основных ветвей тройничного нерва к иннервируемым органам.
Тройничный нерв имеет одно двигательное ядро и три чувствительных ядра. Двигательное ядро тройничного нерва лежит в покрышке моста. Среди чувствительных ядер выделяют:
• среднемозговое, или мезэнцефалическое, ядро тройничного нерва, расположенное в покрышке ствола мозга от моста до среднего мозга; оно обеспечивает преимущественно проприоцептивную чувствительность глазодвигательных мышц;


Рис. 70. Тройничный нерв (V пара): его ядра, ветви и области иннервации.
• главное чувствительное, или мостовое, ядро тройничного нерва, лежа
щее в покрышке моста; обеспечивает тактильную и проприоцептив-
ную чувствительность;
• спинномозговое ядро тройничного нерва, расположенное в покрышке
моста и продолговатого мозга, а также частично в задних рогах шей
ных сегментов Си спинного мозга; обеспечивает болевую и тактиль
ную чувствительность.
Тройничный нерв дает три основные ветви: первую — глазной нерв, вторую — верхнечелюстной нерв и третью — нижнечелюстной нерв (рис. 70).
Глазной нерв проходит в глазницу через верхнюю глазничную щель. Он иннервирует кожу лба, темени и слизистую оболочку верхнего отдела полости носа. В составе этого нерва имеются чувствительные проприоцептивные волокна, идущие от мышц глазного яблока.
 Верхнечелюстной нерв проходит через круглое отверстие в основании черепа. Он дает ряд ветвей, иннервирующих десны и зубы верхней челюсти, кожу носа и щек, а также слизистую оболочку носа, нёба, пазух клиновидной кости основания черепа и верхней челюсти.
Верхнечелюстной нерв проходит через круглое отверстие в основании черепа. Он дает ряд ветвей, иннервирующих десны и зубы верхней челюсти, кожу носа и щек, а также слизистую оболочку носа, нёба, пазух клиновидной кости основания черепа и верхней челюсти.
Нижнечелюстной нерв проходит через овальное отверстие в основании черепа. Он делится на ряд ветвей: сенсорные ветви иннервируют десны и зубы нижней челюсти (нижний альвеолярный нерв, проходящий в толще нижней челюсти), слизистую оболочку языка (язычный нерв) и щек, а также кожу щек и подбородка; двигательные ветви иннервируют жевательные и некоторые другие мышцы.
Нейроны сенсорных ядер тройничного нерва (вторые нейроны сенсорного пути) дают начало нервным волокнам, которые после перекреста в покрышке ствола мозга, формируют тройничную петлю — восходящий путь общей чувствительности от органов головы и шеи. Он присоединяется к медиальной и спиналъной петлям и далее вместе с ними направляется к группе вентролатеральных ядер таламуса. Ответвления аксонов нейронов тройничного узла и сенсорных ядер направляются к ядрам других черепных нервов, ретикулярной формации, мозжечку, пластинке крыши среднего мозга, суб-таламическому ядру, гипоталамусу и многим другим образованиям головного мозга.
Лицевой нерв (n. facialis) — VII пара. Этот нерв имеет три ядра: ядро лицевого нерва — двигательное, расположенное в покрышке моста ближе к срединной плоскости под ядром отводящего нерва; ядро одиночного пути — сенсорное, общее с IX и X парами, расположенное в покрышке продолговатого мозга; верхнее слюноотделительное ядро — парасимпатическое, расположенное в покрышке моста.
На основании мозга лицевой нерв появляется из ямки между мостом, нижней оливой продолговатого мозга и нижней ножкой мозжечка. Вместе с преддверно-улитковым нервом он проходит через внутреннее слуховое отверстие в толщу пирамиды височной кости, где идет в лицевом канале и выходит через шило-сосцевидное отверстие в основании мозгового черепа. В зачелюстной ямке лицевой нерв разветвляется на моторную и сенсорную ветви (рис. 71).
Двигательные ветви лицевого нерва иннервируют мимические мышцы лица и мышцы свода черепа, а также мышцы шеи жаберного происхождения — подкожную мышцу шеи, шилоподъязычную и заднее брюшко двубрюшной мышцы.
Сенсорная порция лицевого нерва лежит обособленно; ее иногда недостаточно обоснованно называют промежуточным нервом. Сенсорный узел лицевого нерва (узел коленца) расположен в лицевом канале в толще пирамиды височной кости. Лицевой нерв содержит вкусовые волокна, идущие от вкусовых луковиц передних 2/3 языка, от мягкого нёба к нейронам узла коленца и далее по их центральному отростку к ядру одиночного пути.
В составе лицевого нерва проходят также парасимпатические (секреторные) волокна. Они берут начало в верхнем слюноотделительном ядре и по специальной ветви (барабанной струне) достигают поднижнечелюстного узла, где переключаются на нейроны, отростки которых в виде постганглио-


Рис. 71. Лицевой нерв (VII пара): его ядра, ветви и области иннервации.


Рис. 72. Языкоглоточный нерв (IX пара): его ядра, ветви и области иннервации.
нарных волокон следуют к подъязычной и подчелюстной слюнным железам, а также к железам слизистой оболочки полости рта.
Языкоглоточный нерв (n. glossopharyngeus) — IX пара. Этот нерв имеет три ядра, расположенных в покрышке продолговатого мозга: двойное ядро (двигательное, общее с X и XI парами), ядро одиночного пути (сенсорное, общее с VII и X парами) и нижнее слюноотделительное ядро (парасимпатическое).
Языкоглоточный нерв выходит из продолговатого мозга через латеральную заднюю борозду продолговатого мозга позади оливы и покидает полость черепа вместе с X и XI парами черепных нервов через яремное отверстие, в котором лежит сенсорный верхний узел языкоглоточного нерва. Чуть ниже за пределами полости черепа располагается сенсорный нижний узел нерва. Далее языкоглоточный нерв спускается по боковой поверхности шеи, разделяясь на несколько ветвей (рис. 72).
Языкоглоточный нерв и его ветви состоят из сенсорных, двигательных и парасимпатических волокон.

Рис. 73. Блуждающий нерв (X пара): его ядра, ветви и области иннервации.
Сенсорные волокна общей чувствительности в составе языкоглоточного нерва начинаются от нейронов обоих сенсорных узлов, сенсорные волокна вкусовой чувствительности — в нижнем узле. Их периферические отростки иннервируют слизистую оболочку нёбной миндалины и нёбных дужек, глотки, задней трети языка, барабанную полость. Центральные отростки
 направляются к ядру одиночного пути. От языкоглоточного нерва отходит ветвь каротидного синуса, которая направляется к месту разветвления общей сонной артерии на внутреннюю и наружную сонные артерии. Здесь расположены хемо- и барорецепторы, сигнализирующие о состоянии внутренней среды организма.
направляются к ядру одиночного пути. От языкоглоточного нерва отходит ветвь каротидного синуса, которая направляется к месту разветвления общей сонной артерии на внутреннюю и наружную сонные артерии. Здесь расположены хемо- и барорецепторы, сигнализирующие о состоянии внутренней среды организма.
Двигательные волокна являются аксонами нейронов двойного ядра. В составе нерва они иннервируют шилоглоточную мышцу, которая при глотании приподнимает глотку и гортань, констрикторы (мышцы-сжиматели) глотки, а также ряд мышц мягкого нёба.
Вегетативные волокна начинаются от нейронов нижнего слюноотделительного ядра, лежащего в покрышке продолговатого мозга. Продолжаясь в составе языкоглоточного нерва, они по его ветвям достигают ушного узла, где переключаются на его нейроны. Идущие от него постганглионарные парасимпатические волокна обеспечивают секреторную иннервацию околоушной слюнной железы.
Блуждающий нерв (n. vagus) — X пара. Этот нерв имеет три ядра, расположенных в покрышке продолговатого мозга: двойное ядро (двигательное, общее с IX и XI парами), ядро одиночного пути (сенсорное, общее с VII и IX парами) и заднее ядро блуждающего нерва (парасимпатическое).
Блуждающий нерв является самым крупным парасимпатическим нервом. Он принимает участие в афферентной и эфферентной иннервации органов дыхания, сердца, желез внутренней секреции и пищеварительного тракта (рис. 73). Блуждающий нерв выходит из вещества продолговатого мозга несколько ниже языкоглоточного нерва и вместе с ним и добавочным нервом покидает полость черепа через яремное отверстие. В шейном отделе от блуждающего нерва отходят глоточные ветви, верхний гортанный нерв и ряд других мелких ветвей. Он отдает верхнюю и нижнюю шейные сердечные ветви, а в грудной части — грудные сердечные ветви. Вместе с сердечными нервами, отходящими от симпатического ствола, они образуют сердечное сплетение. В грудную полость блуждающий нерв проникает через верхнее отверстие грудной клетки, где дает ветви к пищеводу, легким, бронхам и околосердечной сумке, образуя одноименные нервные сплетения на этих органах. Вместе с пищеводом блуждающий нерв проникает через диафрагму в брюшную полость, где иннервирует желудок, печень, селезенку, всю тонкую и часть толстой кишки до ее левого изгиба, почки, а также дает ветви к чревному сплетению (подробнее см. гл. 3).
Многочисленные ветви блуждающего нерва, идущие к различным органам, включают сенсорные, двигательные и вегетативные волокна.
Сенсорные волокна общей чувствительности в составе блуждающего нерва начинаются от псевдоуниполярных нейронов верхнего и нижнего сенсорных узлов, лежащих вблизи яремного отверстия. Периферические отростки части нейронов иннервируют наружный слуховой ход, барабанную перепонку и задний отдел твердой оболочки мозга, а их центральные отростки направляются к спинномозговому ядру тройничного нерва. Другая часть сенсорных нейронов проводит висцеросенсорную информацию от задней трети языка, глотки, гортани и других внутренних органов, иннервируемых блуждающим нервом, к ядру одиночного пути.
 Двигательные волокна в составе ветвей блуждающего нерва начинаются от двойного ядра и иннервируют почти все мышцы мягкого нёба, глотки, гортани.
Двигательные волокна в составе ветвей блуждающего нерва начинаются от двойного ядра и иннервируют почти все мышцы мягкого нёба, глотки, гортани.
Вегетативные волокна берут начало от парасимпатических нейронов заднего ядра блуждающего нерва. Преганглионарные волокна в составе блуждающего нерва направляются к парасимпатическим терминальным ганглиям, расположенным поблизости от внутренних органов или непосредственно в них; ряд мелких парасимпатических ганглиев рассеяны по ходу ствола блуждающего нерва.
Ядра блуждающего нерва связаны с ядрами тройничного, лицевого, языкоглоточного нервов, вестибулярными и ретикулярными ядрами ствола, а также со спинным мозгом. Комплекс этих связей облегчает регуляцию жевания и глотания, реализацию защитных дыхательных, пищеварительных, сердечно-сосудистых рефлексов (глубина и частота дыхания, кашель, рвотный рефлекс, изменение артериального давления, частоты сокращений сердца) и др.
Добавочный нерв (n. accessorius) — XI пара. Этот нерв, являющийся двигательным нервом, в процессе развития отделяется от блуждающего нерва. Он берет начало от двух двигательных ядер. Одно из них, двойное ядро, общее с IX и X парами черепных нервов, лежит в покрышке продолговатого мозга, а другое, спинномозговое ядро добавочного нерва, расположено в передних рогах спинного мозга на уровне шейных CI-VI сегментов (см. рис. 63).
Бульбарная часть добавочного нерва присоединяется к блуждающему нерву и в дальнейшем в виде нижнего гортанного нерва иннервирует мышцы гортани. Волокна спинномозговой части иннервируют грудино-ключично-сосцевидную и трапециевидную мышцы (мышцы шеи и спины).
Мозжечок и его связи
Мозжечок (cerebellum) представляет собой отдел головного мозга, развивающийся из крыши заднего мозга. Его еще нередко называют «малым мозгом». Он располагается в задней черепной ямке под затылочными долями больших полушарий. Под мозжечком находится IV желудочек мозга.
Основные функции мозжечка состоят в поддержании равновесия тела, регуляции мышечного тонуса, осуществлении позно-тонических рефлексов, а также в управлении процессами сенсомоторной координации. Мозжечок программирует плавное, точное и автоматическое выполнение движений, что становится возможным благодаря его связям со спинным мозгом и стволовыми центрами управления движениями, а также с корой больших полушарий.
В мозжечке различают филогенетически более старую среднюю часть — червь, играющий важную роль в регуляции автоматических движений туловища и конечностей, например в процессе ходьбы, и более новую — полушария мозжечка, принимающие участие преимущественно в управлении координированными автоматизированными движениями конечностей.
Сверху мозжечок покрыт слоем серого вещества — корой, образующей многочисленные поперечные извилины — листки мозжечка. Группы лист-
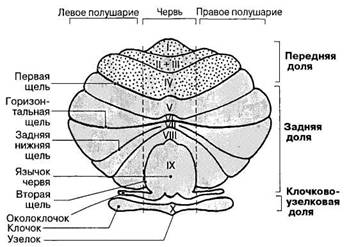
Рис. 74. Мозжечок человека: части, доли и дольки (римскими цифрами обозначены дольки червя).
ков, отделенные глубокими бороздами, образуют дольки мозжечка (рис. 74). Посредством более глубоких щелей мозжечок делится на переднюю, заднюю и клочково-узелковую доли. В переднюю долю мозжечка приходят импульсы преимущественно из спинного мозга, в заднюю — от коры больших полушарий после их переключения в собственных ядрах моста. Наименьшая по размерам клочково-узелковая доля состоит из небольшой парной обособленной дольки — клочка — и расположенного рядом участка червя — узелка, связанных между собой с помощью ножек клочка. Клочково-узелковая доля представляет собой самую древнюю часть мозжечка, участвующую в управлении вестибулярными рефлексами. С помощью двусторонних связей она наиболее тесно взаимодействует с вестибулярными ядрами моста.

|
| Рис. 75. Слои в коре мозжечка новорожденного. Окрашивание по Нисслю. х90. |
В коре мозжечка выделяют три слоя (рис. 75). Наружный молекулярный слой беден нервными клетками и состоит преимущественно из белого веще-

Рис. 76. Нейроархитектоника коры мозжечка.
ства. Во внутреннем зернистом слое находится большое число плотно расположенных зерновидных нейронов малого размера. Между молекулярным и зернистым слоями находится ганглионарный слой, состоящий из расположенных в один ряд грушевидных нейронов, или клеток Пуркинье, — самых крупных нейронов коры мозжечка. Каждая клетка Пуркинье образует густо ветвящееся дендритное дерево, лежащее в молекулярном слое, куда продолжаются аксоны зерновидных клеток. Аксоны клеток Пуркинье обеспечивают эфферентный выход из коры мозжечка на его ядра.
Афферентные волокна от спинного мозга и от различных ядер ствола заканчиваются в зернистом слое коры мозжечка, где образуют многочисленные контакты с дендритами зерновидных нейронов. Это — моховидные волокна (рис. 76). Возбуждение от этих волокон через аксоны зерновидных нейронов передаются на дендриты клеток Пуркинье. Здесь же (на дендритах клеток Пуркинье) заканчиваются лиановидные волокна, идущие в мозжечок от нижних оливных ядер продолговатого мозга. Лиановидные волокна оказывают быстрое возбуждающее действие на клетки Пуркинье, позволяющее мозжечку контролировать быстрые и неосознанные двигательные реакции на внезапные внешние раздражители. Зерновидные клетки оказывают на грушевидные нейроны медленное возбуждающее действие, сопровождающее регуляцию позно-тонических рефлексов и обучение новым типам движений. В молекулярном и зернистом слоях коры мозжечка имеется также несколько разновидностей мультиполярных нейронов. Все они представляют собой интернейроны тормозного типа, избирательно подавляющие активность зерновидных нейронов и клеток Пуркинье, что важно при выработке координации в процессе обучения новым движениям.
Внутри мозжечка различают белое вещество и находящиеся в нем парные ядра серого вещества, тесно связанные с корой червя {ядра шатра), корой

Рис. 77. Ядра мозжечка.
околочервячной зоны {шаровидные и пробковидные ядра) и корой, покрывающей полушария {зубчатые ядра) (рис. 77). Белое вещество состоит из волокон, связывающих участки коры в различных дольках и листках мозжечка, ядра ствола мозга с корой мозжечка, а также кору с ядрами мозжечка.
Волокна от ядер мозжечка, связанных с червем, направляются в основном к вестибулярным и ретикулярным ядрам, и от них — в спинной мозг. Волокна от зубчатого ядра идут к таламусу и далее к коре больших полушарий, а также к красному ядру, от которого начинается красноядерно-спинномозговой путь к мотонейронам спинного мозга.
Связи мозжечка со стволом мозга и спинным мозгом осуществляются с помощью трех пар ножек, состоящих из белого вещества. Посредством верхних ножек мозжечок соединяется со средним мозгом, средних — с мостом и нижних — с продолговатым мозгом и спинным мозгом.
По нижним ножкам к мозжечку направляются волокна заднего спинно-мозжечкового пути, идущие в составе боковых канатиков спинного мозга. Через нижние ножки мозжечок получает также волокна от нижнего оливно-го ядра, от ядер преддверно-улиткового нерва (VIII пара) и от расположенных в продолговатом мозге тонкого и клиновидного ядер (наружные дугообразные волокна).
По средним ножкам к мозжечку идут волокна корково-мосто-мозжечко-вого пути, обеспечивающие связи коры мозжечка с различными отделами коры больших полушарий (подробнее см. гл. 5).
По верхним ножкам к мозжечку проходят волокна от переднего спинно-мозжечкового пути, несущие проприоцептивную информацию о работе спинномозговых центров рефлекторной регуляции движений. В противопо-
 ложном направлении от зубчатого ядра мозжечка к покрышке среднего мозга идут волокна, которые заканчиваются в красном ядре среднего мозга (зубчато-красноядерный путь). Это основной путь влияния мозжечка на систему подсознательной регуляции движений — экстрапирамидную систему.
ложном направлении от зубчатого ядра мозжечка к покрышке среднего мозга идут волокна, которые заканчиваются в красном ядре среднего мозга (зубчато-красноядерный путь). Это основной путь влияния мозжечка на систему подсознательной регуляции движений — экстрапирамидную систему.
При регуляции сложнокоординированных движений тонкие влияния мозжечка на спинной мозг транслируются через красное ядро среднего мозга. При этом мозжечок обеспечивает преодоление организмом силы тяжести в процессе движения, а также целесообразное перемещение общего центра тяжести тела (ОЦТ) относительно площади опоры. Он определяет, где находятся в каждый данный момент части тела и сравнивает их действительное положение с тем, которое должно быть. Например, при ходьбе или беге он контролирует установку и движение туловища и рук в гармонии с движениями ног, а также с перемещением ОЦТ. При письме мозжечок регулирует поддержание оптимальной позы и координацию движений головы, глаз и рук. Мозжечок играет также важную роль при выполнении быстрых последовательных и одновременных движений, таких, например, как движения рук пианиста или машинистки.
Благодаря связям с корой больших полушарий мозжечок координирует активность моторной коры и спинного мозга, способствуя более гладкому выполнению контролируемых ими тонких движений. Он обеспечивает хранение и своевременное использование уже выработанных алгоритмов и программ сложнокоординированных движений и активно участвует совместно с корой больших полушарий и подкорковыми центрами в формировании новых двигательных навыков. Фактически мозжечок является самообучающейся системой, которая в свою очередь существенно облегчает и ускоряет процессы обучения в больших полушариях и стволовых двигательных центрах.
Промежуточный мозг
Промежуточный мозг (diencephalon) располагается между конечным и средним мозгом. На основании мозга его граница спереди проходит по передней поверхности перекреста зрительных нервов, переднему краю заднего продырявленного вещества и зрительным трактам, а сзади — по краю ножек мозга. На дорсальной поверхности передней границей является терминальная полоска, отделяющая промежуточный мозг от конечного мозга, а задней границей — борозда, отделяющая промежуточный мозг от верхних холмиков среднего мозга. На сагиттальном срезе промежуточный мозг виден под мозолистым телом и сводом (рис. 78).
Дорсальный отдел промежуточного мозга представляет собой филогенетически более молодой таламический мозг, являющийся высшим подкорковым сенсорным центром, в котором переключаются практически все афферентные пути, несущие сенсорную информацию от органов тела и органов чувств. Вентральный отдел, гипоталамус, более старое в филогенетическом отношении образование, играет роль высшего центра регуляции вегетативных функций организма.


Рис. 78. Промежуточный мозг на сагиттальном срезе головного мозга.
Таламический мозг в свою очередь подразделяется на парные образования — талaмус (зрительный бугор), метаталамус (заталамическая область), эпиталамус (надталамическая область) и субталамус (подталамическая область).
Полостью промежуточного мозга является III желудочек, который посредством правого и левого межжелудочковых отверстий сообщается с боковыми желудочками, расположенными внутри больших полушарий, и посредством водопровода мозга — с полостью IV желудочка мозга. В верхней стенке III желудочка располагается сосудистое сплетение, участвующее наряду со сплетениями в других желудочках мозга в образовании спинномозговой жидкости.
Таламус
Таламус (thalamus), или зрительный бугор, представляет собой образование яйцевидной формы, состоящее в основном из скопления многочисленных ядер. Таламусы образуются за счет утолщения боковых стенок промежуточного мозга (рис. 79). Спереди заостренная часть таламуса образует передний бугорок, в котором располагаются промежуточные центры сенсорных (афферентных) путей, идущих от ствола мозга в кору больших полушарий. Задняя, расширенная часть таламуса — подушка — содержит подкорковый зрительный центр.
Медиальные поверхности таламусов обращены друг к другу и образуют боковые стенки III желудочка; посередине они соединены между собой межталамическим сращением. Латеральная поверхность таламуса граничит с


Рис. 79. Дорсальная поверхность промежуточного мозга и части ствола мозга.
внутренней капсулой — слоем белого вещества полушарий головного мозга, состоящего из проекционных волокон, соединяющих кору больших полушарий с нижележащими мозговыми структурами.
Толща серого вещества таламуса разделена вертикальной Y-образной прослойкой (пластинкой) белого вещества на три части — переднюю, медиальную и латеральную (рис. 80). Передняя часть таламуса лежит внутри раздвоенной части пластинки, медиальная и латеральная — соответственно внутри и снаружи от этой пластинки. В каждой из этих частей таламуса находится несколько групп таламических ядер. Всего в таламусе по разным данным содержится от 40 до 150 специализированных ядер, объединенных в 8 основных групп. Ниже рассматриваются только основные группы ядер таламуса, необходимые для понимания его связей с другими отделами мозга, а также его функциональной роли в передаче сенсорной (чувствительной) информации в кору больших полушарий.
В передней части таламуса располагается передняя группа таламических ядер. Наиболее крупные из них — передневентральное ядро и переднемеди-альное ядро. Они получают афферентные волокна в составе сосцевидно-та-ламического пути от сосцевидных тел — обонятельного центра промежуточного мозга. От передних ядер эфферентные волокна направляются к поясной извилине коры больших полушарий. Передняя группа таламических ядер и связанные с нею структуры являются важным компонентом лимбической системы мозга, управляющей психоэмоциональным поведением, о чем подробнее будет сказано ниже.
В медиальной части таламуса различают медиодорсальное ядро и группу ядер средней линии. Медиодорсальное ядро имеет двусторонние связи с обонятельной корой лобной доли и поясной извилиной больших полушарий, миндалевидным телом и переднемедиальным ядром таламуса. Функцио-


Рис. 80. Топография ядер таламуса.
нально оно тесно связано также с лимбической системой и имеет двусторонние связи с корой теменной, височной и островковой долей мозга. Благодаря этому медиодорсальное ядро непосредственно участвует в реализации высших психических процессов. Его разрушение приводит к снижению беспокойства, тревожности, напряженности, агрессивности, устранению навязчивых мыслей.
Ядра средней линии занимают наиболее медиальное положение в таламусе и довольно многочисленны. Они получают афферентные волокна от гипоталамуса, от ядер шва и голубого пятна ретикулярной формации ствола мозга. Сюда проецируется часть волокон спинно-таламических путей в составе медиальной петли. Эфферентные волокна от нейронов ядер средней линии направляются к гиппокампу, миндалевидному телу и поясной извилине больших полушарий, входящих в состав лимбической системы. Связи с корой больших полушарий двусторонние. Это позволяет ядрам средней
линии играть важную роль в процессах пробуждения и активации коры больших полушарий, а также в обеспечении процессов памяти.
В латеральной части таламуса располагаются дорсолатеральная, вентролaтералъная, вентральная заднемедиалъная и задняя группы ядер.
Ядра дорсолатералъной группы относительно мало изучены в функциональном плане. Есть данные о причастности дорсолатеральных ядер к системе восприятия боли.
Ядра вентролатералъной группы анатомически и функционально существенно различаются между собой. Задние ядра вентролатералъной группы часто рассматриваются как одно вентролатеральное ядро таламуса. Эта группа ядер получает основную массу волокон восходящих путей общей чувствительности в составе медиальной петли. Сюда приходят также волокна вкусовой чувствительности и волокна от вестибулярных ядер. Эфферентные волокна, начинающиеся от ядер вентролатеральной группы, направляются в кору теменной доли больших полушарий, куда проводят соматосенсорную информацию от всего тела.
Задняя группа ядер таламуса состоит из клеточных скоплений подушки таламуса. К ядрам идут афферентные волокна от верхних холмиков четверохолмия и волокна в составе зрительных трактов. Эфферентные волокна, начинающиеся от ядер задней группы, широко распространены в коре лобной, теменной, затылочной, височной и лимбической долей больших полушарий. Ядерные центры подушки таламуса имеют отношение к зрению и причастны к комплексному анализу различных сенсорных раздражителей. По-видимому, они играют значительную роль в перцептивной (связанной с восприятием) и когнитивной (познавательной, мыслительной) деятельности мозга, а также в хранении и воспроизведении информации — памяти.
Интраламинарная группа ядер, таламуса лежит в толще вертикальной Y-образной прослойки белого вещества. Интраламинарные ядра взаимосвязаны с базальными ядрами, зубчатым ядром мозжечка и корой больших полушарий. В функциональном отношении эти ядра играют важную роль в активационной системе мозга. Повреждение интраламинарных ядер в обоих таламусах приводит к резкому снижению двигательной активности (кинетический мутизм), а также апатии и разрушению мотивационной структуры личности.
Следует отметить, что кора больших полушарий благодаря двусторонним связям практически со всеми ядерными центрами таламуса способна оказывать регулирующее воздействие на их функциональную активность.
Эпиталамус
Эпиталамус (epithalamus) включает эпифиз (шишковидное тело) и ряд ядерных скоплений нейронов.
Эпифиз — это железа внутренней секреции, функция которой заключается в тормозном влиянии на работу большей части других эндокринных желез (гипофиза, щитовидной и паращитовидных желез, половых желез, надпочечников и др.). Эпифиз вырабатывает нейрогормон мелатонин, имеющий
большое значение для поддержания иммунного статуса организма. Гормоны эпифиза также играют определенную роль в регуляции сезонных ритмов жизнедеятельности организма.
Метаталамус
Метаталамус (metathalamus) располагается в заднебоковом отделе промежуточного мозга, где под подушкой таламуса лежат два парных овальных образования — более крупное медиальное и меньшее по размеру латеральное коленчатые тела. С помощью ручек верхнего и нижнего холмиков, состоящих из белого вещества, медиальные и латеральные коленчатые тела соединяются соответственно с нижними и верхними холмиками крыши среднего мозга. Сверху коленчатые тела покрыты белым веществом, внутри содержатся скопления серого вещества — ядра.
Ядра медиального коленчатого тела (как и ядра нижнего холмика четверохолмия), являются подкорковым центром слуха, поскольку в них оканчиваются афферентные волокна в составе латеральной петли, берущие начало в области моста (слуховой путь) от ядер преддверно-улиткового (VIIIпара) нерва. Ядра латерального коленчатого тела (вместе с ядрами верхнего холмика четверохолмия) являются подкорковыми центрами зрения: в них оканчивается латеральная часть волокон, идущих в составе зрительного тракта (IIпара). Ядра коленчатых тел формируют также восходящие пути к центрам зрительного и слухового анализаторов в коре больших полушарий.
Субталамус
Субталамус (subthalamus) является подталамической областью, где заканчиваются продолжающиеся из ножек мозга красные ядра и черное вещество. Впереди и латерально от черного вещества располагается субталамическое ядро, или люисово тело. Это ядро относится к образованиям экстрапирамидной системы мозга, обеспечивающей управление автоматизированными сложнокоординированными движениями тела. При локальных повреждениях субталамического ядра возникают сильные «бросковые» движения конечностей (баллизм).
Гипоталамус
Гипоталамус (hypothalamus) представляет собой вентральный отдел промежуточного мозга. В его состав входит комплекс образований, расположенных под III желудочком. Гипоталамус спереди ограничивается зрительным перекрестом (хиазмой), латерально — передней частью субталамуса, внутренней капсулой и зрительными трактами, отходящими от хиазмы. Сзади гипоталамус продолжается в покрышку среднего мозга (рис. 81).
В гипоталамусе различают большую по размерам переднебоковую часть и меньшую заднюю часть.
Переднебоковая часть образует дно IIIжелудочка мозга и включает серый бугор, воронку с гипофизом, зрительные тракты, перекрест зрительных нервов


Рис. 81. Вентральная поверхность промежуточного мозга.
(хиазму), пограничную (терминальную) мозговую пластинку. Сюда же относится собственно подбугорная область — скопление ядер серого вещества гипоталамуса.
Серый бугор — это тонкая, выпуклая снизу часть нижней стенки III желудочка, расположенная между перекрестом зрительных нервов спереди и сосцевидными телами сзади. В стенке серого бугра расположены вегетативные ядра, которые причисляют к эмоциогенным зонам мозга. Истонченное продолжение передней стенки серого бугра кверху и назад представляет собой пограничную пластинку, образующую переднюю стенку III желудочка. Вентрально и чуть кпереди серый бугор образует воронку, которая служит местом присоединения гипофиза.
Подбугорная область расположена под таламусами чуть выше серого бугра и частично внутри между его стенками. Сверху она отделена от таламусов гипоталамической бороздой (рис. 82). Подбугорная область содержит до 40 ядер, функциональная роль которых очень важна.
В передне-боковой части гипоталамуса различают переднюю и среднюю группы гипоталамических ядер (см. рис. 79).
К передней группе относятся паравентрикулярное ядро, преоптическое ядро, супраоптические и супрахиазматические ядра. Паравентрикулярное ядро располагается под эпендимой вдоль стенки III желудочка, преоптическое ядро — спереди ближе к латеральной стенке гипоталамуса над серым бугром, группа супраоптических и супрахиазматических ядер — чуть ниже над хиаз-мой и зрительным трактом своей стороны.
В нейронах паравентрикулярного и супраоптических ядер образуется нейросекрет, который по их аксонам перемещается в задний отдел гипофиза, или нейрогипофиз, где высвобождается в виде нейрогормонов. ва-зопрессина и окситоцина, поступающих в кровь. Вазопрессин, или антидиуретический гормон (диурез — выделение мочи), стимулирует обратное всасывание (ре'зорбцию) воды в почечных канальцах. Повреждение передних ядер гипоталамуса приводит к прекращению выделения вазопресси-на, вследствие чего развивается несахарный диабет. Окситоцин оказывает


Рис. 82. Топография ядер гипоталамуса (сагиттальный срез).
стимулирующее действие на гладкую мускулатуру внутренних органов, например матки. В целом от этих гормонов зависит водно-солевой баланс организма.
В преоптическом ядре образуется один из рилизинг-гормонов — люлибе-рин, стимулирующий выработку в аденогипофизе лютеинизирующего гормона, контролирующего активность половых желез.
Супрахиазматические ядра принимают активное участие в регуляции циклических изменений активности организма — циркадианных, или суточных, биоритмов (например, в чередовании сна и бодрствования).
Известно, что стимуляция переднего отдела гипоталамуса приводит к реакциям парасимпатического типа: сужению зрачка, снижению частоты сокращений сердца, расширению просвета сосудов и падению артериального давления, усилению перистальтики кишечника. Разрушение этого отдела сопровождается также необратимым повышением температуры тела.
К средней группе гипоталамических ядер относят дорсомедиальное и вентромедиальное ядра, ядро серого бугра и ядро воронки. Средняя группа ядер контролирует водный, жировой и углеводный обмен, влияет на уровень сахара в крови, ионный баланс организма, проницаемость сосудов и клеточных мембран. В ядрах средней группы локализуются центры голода и насыщения.
Например, разрушение вентромедиального ядра гипоталамуса приводит к избыточному потреблению пищи (гиперфагии) и ожирению, а повреждение ядра серого бугра — к снижению аппетита и резкому исхуданию (кахексии). Нейроны ядер данной группы вырабатывают ряд рилизинг-гормонов (сома-тостатин, соматолиберин, люлиберин, фоллиберин, пролактолиберин, тире-олиберин и др.). Через гипоталамо-гипофизарную систему они оказывают влияние на ростовые процессы, скорость физического развития и полового созревания, формирование вторичных половых признаков, функции половой системы, а также на обмен веществ. В целом можно сказать, что ядра средней группы имеют отношение к регуляции пищевого и полового поведения, а также к контролю функций эндокринных желез. Есть данные о том, что в этой зоне гипоталамуса находятся специфические центры (центры удовольствия), играющие важную роль в процессах формирования мотиваций и психоэмоциональных форм поведения.
Задняя часть гипоталамуса расположена между серым бугром и задним продырявленным веществом и состоит из правого и левого сосцевидных тел. Внутри каждого из них под тонким слоем белого вещества находятся два серых ядра, относящихся к подкорковым обонятельным центрам.
К задней группе относится также заднее гипоталамическое ядро. Вместе с ядрами передней группы заднее гипоталамическое ядро участвует в терморегуляции, а также содержит центры, координирующие активность симпатической части автономной нервной системы. Стимуляция этого ядра приводит к реакциям симпатического типа: расширению зрачка, повышению частоты сокращений сердца и артериального давления, учащению дыхания и уменьшению тонических сокращений кишечника. Разрушение заднего отдела гипоталамуса вызывает вялость, сонливость и снижение температуры тела.
Гипоталамус имеет широкую систему связей со многими образованиями головного мозга, что объясняет его участие во многих поведенческих реакциях. Система афферентных и эфферентных волокон свода связывает передний гипоталамус и сосцевидные тела с гиппокампом больших полушарий, а сосцевидно-таламический (пучок Вик д'Азира) и сосцевидно-покрышечный пути связывают сосцевидные тела с таламусом и покрышкой среднего мозга.
Гипофиз (hypophysis) — железа внутренней секреции, включающая две доли. Задняя доля, или нейрогипофиз, состоит из нейроглиальньгх клеток и является продолжением воронки гипоталамуса. Спереди находится более крупная доля — аденогипофиз, построенная из железистых клеток. Благодаря тесному взаимодействию гипоталамуса с гипофизом в промежуточном мозге функционирует единая гипоталамо-гипофизарная система, управляющая работой всех эндокринных желез, а с их помощью — вегетативными функциями организма (рис. 83). Взаимодействие с гипофизом осуществляется посредством выделяемых ядрами гипоталамуса нейрогормонов — рилизинг-гормонов. По системе воротных кровеносных сосудов они попадают в переднюю долю гипофиза (аденогипофиз), где способствуют высвобождению тротшых гормонов, стимулирующих синтез специфических гормонов в других эндокринных железах.

Рис. 83. Гипофиз и его влияние на другие эндокринные железы.
Гипоталамо-гипофизарная система осуществляет контроль над гуморальной регуляцией водно-солевого баланса, обменом веществ и энергии, работой иммунной системы, терморегуляцией, репродуктивной функцией и т. д. Выполняя в этой системе регулирующую роль, гипоталамус является высшим центром, управляющим автономной (вегетативной) нервной системой.
Конечный мозг
Конечный мозг (telencephalon), или большой мозг, представляет собой самую развитую и в филогенетическом отношении новую часть головного мозга, непосредственно связанную с наиболее сложными проявлениями психической и интеллектуальной деятельности человека.
Конечный мозг состоит из двух полушарий большого мозга, соединенных между собой мозолистым телом, передней и задней спайками и спайкой свода. Полости конечного мозга образуют правый и левый боковые желудочки, каждый из которых находится в соответствующем полушарии; медиальную стенку бокового желудочка в ростральном отделе образует прозрачная перегородка.
Полушария большого мозга сверху покрыты корой мозга — слоем серого вещества, образованного нейронами более пятидесяти разновидностей. Под корой мозга в больших полушариях находится белое вещество, состоящее из миелинизированных волокон, большая часть которых соединяет кору больших полушарий с другими отделами и центрами головного мозга. В толще белого вещества полушарий находятся скопления серого вещества — базалъ-ные ганглии (подкорковые ядерные центры). Слой белого вещества, называемый внутренней капсулой, отграничивает полушария от таламусов промежуточного мозга.

