




Під ферментативною кінетикою розуміють залежність швидкості реакції, яка прискорюється ферментом, від хімічної природи субстрату і ферменту й умов їх взаємодії (концентрації, температури, рН середовища, наявності активаторів та інгібіторів).
Доцільно звернути увагу на закономірності, які визначаються природою і концентрацією реагуючих речовин (субстратів і ферментів) і їх зміною.
Швидкість ферментативної реакції в значній мірі залежить від хімічної природи і походження субстрату. Наприклад, різні за природою складові крохмалю -амілоза і амілопектин з різною швидкістю гідролізуються амілазами і до того ж потребують неоднакового набору амілолітичних ферментів. Зокрема, для гідролізу амілози достатньо а-амілази, а для гідролізу амілопектину, крім а-амілази необхідні і ферменти, які каталізують гідроліз a-1,6-глюкозидних зв'язків, тобто глюкоа-мілаза;або декстриназа.
На швидкість і умови ферментативних реакцій впливає і походження ферменту. Ферменти солоду гідролізують крохмаль до декстринів, мальтози і незначної кількості (3-5%) глюкози, а ферменти мікробного походження - до декстринів і глюкози. Крім того, ос-амілаза бактеріального походження (ферментні препарати Амілодіастатін, Термаміл та ін.) мають більш високу оптимальну температуру дії (до 90-115° С), що можна корисно використати для створення нових технологій оцукрювання крохмалю сировини.
Ферменти мікробного походження більш стійкі в порівнянні із ферментами солоду до низьких значень рН середовища. Це особливо важливо для створення у виробничих умовах асептичних умов за рахунок зниження рН живильних середовищ, що запобігає розвитку кислоутворюючих бактерій.
Концентрація ферменту
Швидкість більшості ферментативних реакцій пропорційна концентрації ферменту, особливо це справедливо на найбільш ранніх стадіях. Ця пропорційність є основою методів визначення концентрації ферменту.
Початкову швидкість реакції можна визначити з тангенсу кута нахилу кривої залежності кількості субстрату, яка вступила в реакцію від тривалості реакції на

початку координат (до перетворення субстрату не більше 20-25%). Під час подальшого перетворення субстрату не зберігається пропорційність залежності тривалості реакції від кількості ферменту.
Концентрація субстрату
Вплив концентрації субстрату на початкову швидкість ферментативної реакції має суттєве значення. Криві, які характеризують цю залежність є частиною гіперболи (рис. 4.1).
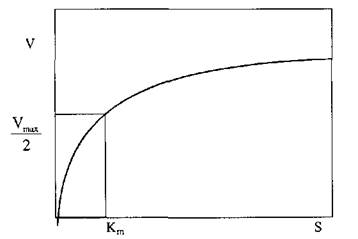
Рис. 4.1 Залежність швидкості ферментативної реакції V від концентрації субстрату  при постійній кількості ферменту
при постійній кількості ферменту
Такі криві характерні для всіх процесів, під час яких проходить проста дисоціація.
У 1913 р. Л.Мехаеліс і М.Ментен розробили теорію дії ферментів, яка передбачає утворення комплексу фермент-субстрат внаслідок оборотної реакції
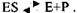
|
Припустивши, що комплекс  утворюється дуже швидко, так що він завжди знаходиться в рівновазі з утворюється дуже швидко, так що він завжди знаходиться в рівновазі з 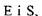 , а розщеплення , а розщеплення 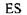 на Е і Р проходить порівняно
ьпиьрпорівняно порівняно на Е і Р проходить порівняно
ьпиьрпорівняно порівняно
|
Константа рівноваги цієї реакції, або її зворотна величина, яку називають константою дисоціації фермент-субстратного комплексу 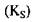 залежить від природи субстрату і ферменту і відображає ступінь їх спорідненості. Чим нижче значення
залежить від природи субстрату і ферменту і відображає ступінь їх спорідненості. Чим нижче значення 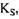 тим вища спорідненість ферменту до субстрату. Наприклад, для
тим вища спорідненість ферменту до субстрату. Наприклад, для 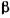 -фруктофура-нозидази (сахарази)
-фруктофура-нозидази (сахарази) 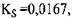 тобто концентрація фермент-субстратного комплексу перевищує концентрацію вільних ферментів і субстрату приблизно в 60 разів. Концентрація фермент-субстратного комплексу змінюється внаслідок перетворення останнього в продукт реакції із регенерацією вільного ферменту
тобто концентрація фермент-субстратного комплексу перевищує концентрацію вільних ферментів і субстрату приблизно в 60 разів. Концентрація фермент-субстратного комплексу змінюється внаслідок перетворення останнього в продукт реакції із регенерацією вільного ферменту
|
повільно і практично не впливає на концентрацію фермент-субстратного комплексу, отримаємо рівняння, яке характеризує зв'язок швидкості ферментативної реакції V із концентрацією субстрату  :
:
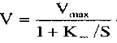
Їм І
де 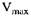 - максимальна швидкість реакції, яка досягається при великих значеннях S,
- максимальна швидкість реакції, яка досягається при великих значеннях S,
і при оптимальній температурі та рН середовища, тобто в тих випадках, коли знаменник правої частини рівняння близький до 1;
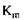 - константа Михаеліса, константа дисоціації в реакції утворення комплексу фермент-субстрат, вона має розмірність концентрації і при
- константа Михаеліса, константа дисоціації в реакції утворення комплексу фермент-субстрат, вона має розмірність концентрації і при 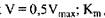 дорівнює концентрації субстрату, при якій швидкість ферментативної реакції дорівнює половині максимальної (рис. 4.1).
дорівнює концентрації субстрату, при якій швидкість ферментативної реакції дорівнює половині максимальної (рис. 4.1).
Величина 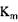 залежить від природи ферменту і субстрату і відображає спорідненість ферменту субстрату. Низька величина
залежить від природи ферменту і субстрату і відображає спорідненість ферменту субстрату. Низька величина 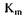 свідчить про високу спорідненість ферменту із субстратом, бо максимальна швидкість реакції досягається уже при низьких концентраціях субстрату.
свідчить про високу спорідненість ферменту із субстратом, бо максимальна швидкість реакції досягається уже при низьких концентраціях субстрату.
Якщо фермент насичений субстратом, то його дія не залежить від концентрації субстрату, а тільки від кількості ферменту І прямо пропорційна їй.
У тому випадку, коли концентрація субстрату менша величини 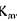 , тоді дія ферменту не залежить від його кількості, а залежить головним чином від концентрації субстрату.
, тоді дія ферменту не залежить від його кількості, а залежить головним чином від концентрації субстрату.
Для прикладу в табл. 4.1 наведені значення 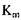 для глюкоамілази в залежності від субстрату.
для глюкоамілази в залежності від субстрату.
Табл. 4.1 Вплив виду лінійних поліцукридів на величину К
| Субстрат | Довжина ланцюга | 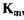 , молярна , молярна
| 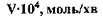
|
| субстрату, | концентрація | ||
| глюкозних | |||
| одиниць | |||
| Мальтоза | 1,16×10-3 | 0,513 | |
| Мальтотріоза | 2, 02×10-4 | 1,150 | |
| Амілодекстрин | 4,90×10-5 | 0,855 | |
| Амілоза | 3,84×10-6 | 0,783 |
Для коротких інтервалів часу і для найбільш ранніх стадій ферментативної реакції можна припустити, що концентрація субстрату залишається майже постійною, особливо у випадках, коли субстрату міститься значний надлишок. У таких умовах проходження реакції підпорядковується закону нульового порядку, а кількість утвореного продукту пропорційна тривалості реакції
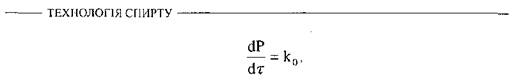
де 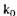 - константа реакції нульового порядку. У цих реакціях кількість кінцевого продукту подвоюється, якщо подвоюється тривалість реакції.
- константа реакції нульового порядку. У цих реакціях кількість кінцевого продукту подвоюється, якщо подвоюється тривалість реакції.
Якщо ж ферментативна реакція виходить за межу початкової стадії, необхідно враховувати безперервне зменшення концентрації субстрату, що приводить до сповільнення реакції. Більшість ферментативних мономолекулярних реакцій підпорядковується кінетичному закону реакції першого порядку
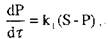
де 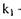 -константа реакції першого порядку;
-константа реакції першого порядку; 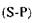 - концентрація субстрату, що за-
- концентрація субстрату, що за-
лишився в будь-який заданий час.
Швидкість реакції прямо пропорційна концентрації субстрату, що залишився. У рівні проміжки часу перетворюється половина кількості субстрату, що залишилась. Наприклад, якщо за першу годину перетворилося 50% субстрату, то за другу - 25% початкового, за третю - 12,5% і так далі.
Після інтегрування попереднього рівняння одержимо
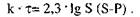
Таким чином, швидкість гідролізу поліцукридів при постійній концентрації ферменту і температурі пропорційна тільки концентрації поліцукридів у розчині. Гідроліз поліцукридів одним ферментом описується кінетичним рівнянням першого порядку.
У випадку дії на поліцукриди декількох амілаз у результаті дії 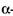 -амілази утворюється значна кількість нередукуючих кінців, що полегшує і прискорює дію інших амілаз -
-амілази утворюється значна кількість нередукуючих кінців, що полегшує і прискорює дію інших амілаз - 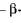 амілази або глюкоамілази.
амілази або глюкоамілази.
Температура
Температура впливає на швидкість реакції утворення фермент-субстратного комплексу і на всі наступні етапи перетворення субстрату, що призводить до прискорення каталізу. При енергії активації 48 кДж-кмоль 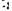 (для ферментативного гідролізу крохмалю вона дорівнює 45 кДж-кмоль
(для ферментативного гідролізу крохмалю вона дорівнює 45 кДж-кмоль  ) швидкість реакції подвоюється при збільшенні температури від 22 до 32° С.
) швидкість реакції подвоюється при збільшенні температури від 22 до 32° С.
При значному підвищенні температури проходить теплова денатурація ферментативного білку, що призводить до поступової втрати ферментом каталітичних властивостей, проявляється теплова інактивація.
Тому при температурах, вищих 50° С, денатурація ферментативного білка різко посилюється, і хоч швидкість перетворення субстрату продовжує рости, активність ферменту, виражена кількістю перетвореного субстрату за одиницю часу, падає.
Температура, при якій каталітична активність ферменту максимальна, називається його температурним оптимумом.
|
Температурний оптимум для різних ферментів неоднаковий.
Якщо визначити константу швидкості реакції при двох різних температурах, то енергія активації може бути виражена такою формулою (відповідно із рівняння Арреніуса)
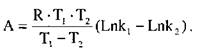
де А - енергія активації, кДж-кмоль 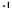 ;
;
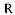 - універсальна газова постійна, дорівнює 8,3134 кДж/(моль • К);
- універсальна газова постійна, дорівнює 8,3134 кДж/(моль • К); 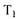 і Т2 - абсолютні температури, К;
і Т2 - абсолютні температури, К;
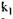 і
і 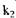 - константи швидкості реакцій відповідно при температурі
- константи швидкості реакцій відповідно при температурі 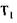 і Т2. Чим нижча енергія активації, тим досконаліше проходить процес. Зміни активності в залежності від температури різні у одного і того ж ферменту, але різного походження. Наприклад, термостабільність амілаз можна записати в такій послідовності: бактеріальна > солодова > грибна.
і Т2. Чим нижча енергія активації, тим досконаліше проходить процес. Зміни активності в залежності від температури різні у одного і того ж ферменту, але різного походження. Наприклад, термостабільність амілаз можна записати в такій послідовності: бактеріальна > солодова > грибна.
Температурний оптимум для ферментів солоду із різних культур зерна неоднаковий (рис. 4.2). Для ячмінного і просяного солодів він не перевищує 55° С, для вівсяного - 50° С.
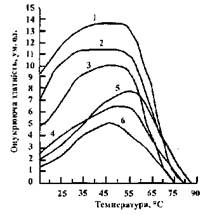
Рис. 4.2 Вплив температури на оцукрюючу здатність солодів:
1 - житнього; 2 - пшеничного; 3 - ячмінного; 4 - кукурудзяного;
5 - просяного; 6 - вівсяного
При подальшому підвищенні температури проходить теплова денатурація білкової молекули фермента і зниження а£о повна втрата (75-85° С) каталітичної активності (дані приведені для гідролізу 2%-ного розчину крохмалю). Підвищення концентрації вуглеводів у суслі сприяє збільшенню їх термостабільність Наприклад, оптимальна температура для дії 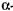 -амілази ячмінного солоду в суслі - біля 70° С,
-амілази ячмінного солоду в суслі - біля 70° С, 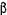 -амілази - 60° С. У виробничих умовах температуру оцукрювання тримають в межах 57-58° С. Небажана менша температура, бо вона може сприяти розвитку кислотоутворюючих бактерій.
-амілази - 60° С. У виробничих умовах температуру оцукрювання тримають в межах 57-58° С. Небажана менша температура, бо вона може сприяти розвитку кислотоутворюючих бактерій.

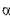 -амілази мікробного походження мають різні температурні оптимуми дії (в °С):
-амілази мікробного походження мають різні температурні оптимуми дії (в °С): 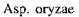 - 52,
- 52, 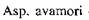 - 55,
- 55, 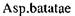 - 60,
- 60, 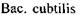 - 70,
- 70,
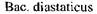 - 80. Глкжоамілази різних продуцентів мають температурний оптимум 55-60° С.
- 80. Глкжоамілази різних продуцентів мають температурний оптимум 55-60° С.
Іноземні фірми ( , Данія та ін.) виробляють концентровані ферме-
, Данія та ін.) виробляють концентровані ферме-
нтні препарати, які мають різні температурні оптимуми. Наприклад, ферментні препарати, які містять 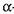 -амілазу,
-амілазу,  (продуцент селекційований штам
(продуцент селекційований штам
 ) відрізняється високою термостабільністю - 90-95° С;
) відрізняється високою термостабільністю - 90-95° С;  (продуцент селекційований штам
(продуцент селекційований штам  І - 75° С.
І - 75° С.
Ферментний препарат  , що містить глюкоамілазу (продуцент селекційо-
, що містить глюкоамілазу (продуцент селекційо-
ваний штам  ) має температурний оптимум 75° С, рН - 4,0-4,5. Ак-
) має температурний оптимум 75° С, рН - 4,0-4,5. Ак-
тивно діє протягом усього періоду бродіння.
Ферментний препарат  . містить глюкоамілазу, грибну
. містить глюкоамілазу, грибну 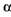 -амілазу і
-амілазу і
бактеріальну нейтральну протеіназу. Оптимум дії - 55° С.
Під час зброджування сусла (температура 28-30° С) бактеріальні 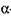 -амілази майже припиняють каталітичну дію, тоді як
-амілази майже припиняють каталітичну дію, тоді як 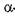 -амілаза мікроміцетів продовжує діяти.
-амілаза мікроміцетів продовжує діяти.
Оптимум температури дії ферментів залежить від тривалості реакції - для коротких періодів оптимальна температура більш висока, ніж для тривалих.
Температурний оптимум дії амілаз не співпадає з оптимальною температурою їх стабільності. Оптимальна температура стабільності ферментів нижча температурного оптимуму їх активності.
У першій основній стадії оцукрювання крохмаль гідролізується частково і гідроліз продовжується при зброджуванні сусла. Тому для зберігання активності амі-лолітичних ферментів температура в першій стадії оцукрювання не повинна перевищувати оптимальну. У виробничих умовах вона складає 57-58° С.
Продукти гідролізу крохмалю і білки сповільнюють інактивацію ферментів. Тому у виробничому суслі оптимальна температура оцукрювання може збільшуватися з підвищенням концентрації сухих речовин.
Величина рН середовища
Зміни рН середовища значно впливають на активність ферментів. Вплив концентрації іонів водню на каталітичну активність ферментів полягає у впливі на їх активні центри. При різних значеннях рН реакційного середовища активний центр може бути більше або менше екранований сусідніми з ним фрагментами поліпеп-тидного ланцюга білкової частини ферменту, більш сильно або слабоіонізований. Кислотні й основні групи ферментів здатні до іонізації. При зміні рН середовища внаслідок приєднання 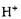
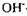 -іонів зменшується ступінь іонізації одних або
-іонів зменшується ступінь іонізації одних або
других груп. У тому випадку, коли ці групи є активними центрами ферментів, іони 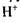 або
або 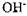 відіграють роль конкурентних інгібіторів ферментів. В інших випадках іони
відіграють роль конкурентних інгібіторів ферментів. В інших випадках іони 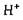 *бо
*бо 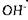 призводять до порушення конформації і комплементарності ферменту і субстрату, тобто виступають як реконкурентні інгібітори.
призводять до порушення конформації і комплементарності ферменту і субстрату, тобто виступають як реконкурентні інгібітори.
Крім того, рН середовища впливає на сутність іонізації субстрату, фермент-субстратного комплексу і продуктів реакції, має великий вплив на стан ферментно-

----------------------------------------- Одержання солоду та мікробних ферментних препаратів ------------
го білка, визначаючи співвідношення в ньому катіонних і аніонних центрів, що впливає на третичну структуру білкової молекули. Певна третична структура біл-ка-фермента необхідна для утворення фермент-субстратного комплексу.
Оптимальне значення рН дії ферменту залежить від природи ферменту і субстрату, його концентрації, від стабільності ферменту, температури середовища та тривалості каталітичної реакції.
Термін "оптимум рН" не має строго визначеної фізичної суті. Правильніше показувати межі рН, сприятливі для даної ферментативної реакції.
Максимальна активність 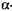 -амілази ячмінного солоду при рН 5,4-5,7,1
-амілази ячмінного солоду при рН 5,4-5,7,1 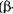 -амілази - 4,5-5,0.
-амілази - 4,5-5,0.
З підвищенням температури середовища оптимальне значення рН дії ферменту збільшується. Наприклад, оптимальне значення рН дії амілаз при температурі 50° С - 4,8, 60° С - 5,1, 70° С - 5,9.
Більшість глюкоамілаз 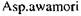 має оптимальне значення рН 4,5. Тому в
має оптимальне значення рН 4,5. Тому в
процесі зброджування сусла активність глюкоамілаз у значній мірі зберігається, навіть при зниженні рН до 4,0.
Амілази мікробного походження більш стабільні в порівнянні з амілазами солоду при знижених значеннях рН бражки, що дуже важливо для умов спиртового виробництва. Це дозволяє більш повно провести дооцукрення граничних декстринів бражки навіть при зниженні її рН.

