




Лекція 8. СИНТЕЗ БІЛКІВ
1. Транспортні РНК
1.1. Структура тРНК
1.2. Аміноацилювання тРНК
2. Рибосома
2.1. Склад рибосоми
2.2. Структура рибосоми
3. Елонгаційний цикл
3.1. Елонгаційний фактор EF1
3.2. Зв'язування аа-тРНК з А-сайтом рибосоми
3.3. Транспептидація
3.4. Транслокація
4. Ініціація трансляції
4.1. Ініціація трансляції у прокаріотів
4.2. Ініціація трансляції в еукаріотів
5. Термінація трансляції
6. Регуляція трансляції
7. Формування просторової структури білка
7.1. Закономірності укладання білкової глобули
7.2. Шаперони hsp70
7.3. Шапероніни
Інформація щодо амінокислотної послідовності білка записана у вигляді послідовності нуклеотидів мРНК у відповідності з генетичним кодом. Зчитування цієї інформації та її переклад у амінокислотний текст (трансляція, translation) має розпочинатися зі стартового кодону, де при ініціації трансляції відбувається остаточне збирання головного пристрою трансляції – рибосоми (ribosome). Рибосома – комплекс рибосомної РНК і білків – сканує нуклеотидну послідовність мРНК, рухаючись уздовж неї кроками по три нуклеотиди від 5′- до 3′-кінця під час елонгації трансляції до стоп-кодону, де відбувається термінація процесу. Під час сканування рибосома працює як декодуючий пристрій, забезпечуючи впізнання кодонів комплементарними триплетами (антикодонами) у складі тРНК (транспортні РНК, tRNA ––transfer RNA), і як каталізатор процесу синтезу пептидного зв’язку між амінокислотами. Певний антикодон відповідає амінокислоті певного типу, яку несе на собі тРНК. Отже, тРНК виступають ключовою ланкою реалізації генетичного коду: саме вони забезпечують доставку амінокислот до рибосоми у порядку, який відповідає послідовності кодонів.
Транспортні РНК
Структура тРНК
Молекули тРНК містять 74 – 95 (найчастіше 76) нуклеотидів, серед яких зустрічаються неканонічні (утворюються внаслідок посттранскрипційних модифікацій) – тимідин, інозин (І), дигідроуридин (D), псевдоуридин (ψ) тощо.
У складі молекули формуються комплементарні дволанцюгові стебла та шпильки за єдиною для всіх тРНК схемою, що нагадує лист конюшини (рис. 1). Кінцеві фрагменти ланцюга об’єднуються у дволанцюгове стебло, причому чотири нуклеотиди на 3′-кінці залишаються неспареними. 3′-кінцевий триплет ССА є стандартним для всіх тРНК, до рибози кінцевого аденозину ковалентно приєднується амінокислота: відповідно, стебло називають акцепторним. 5′-кінцева частина акцепторного стебла переходить у шпильку з петлею, яка часто містить дигідроуридин – D-стебло і D-петля. У деяких тРНК D-петля може містити на один нуклеотид менше або на один–три нуклеотиди більше відносно типового розміру петлі, зображеного на рис. 1. D-стебло переходить у стебло з петлею, у складі якої розташований антикодоновий триплет (у позиції 34 – 36), – антикодонове стебло / антикодонова петля. За антикодоновим стеблом розташована варіабельна петля (V-петля). За розміром V-петлі всі тРНК можна поділити на два структурні класи: до першого належить більшість тРНК із короткою петлею (3 – 5 нуклеотидів), до другого – кілька тРНК із довгою (до 16 додаткових нуклеотидів) петлею. Нумерація нуклеотидів на рис. 1 є стандартною, додаткові нуклеотиди D- та V-петлі, якщо вони присутні, нумерують окремо. За варіабельною петлею міститься ТψС-стебло з петлею (у складі петлі часто зустрічається консенсус – ТψС), яке переходить у 3′-кінцеву частину акцепторного стебла.
тРНК конкретного типу, котра відповідає певній амінокислоті, позначають індексом, наприклад, тРНКAla. Якщо в молекулі тРНК є амінокислота, то таку аміноацильовану тРНК позначають як Ala-тРНКAla. Загальне позначення для аміноацильованих тРНК – аа-тРНК (aa-tRNA ––aminoacyl tRNA).
Еукаріотичні гени тРНК (близько 500 активних генів тРНК у геномі людини, частина яких зібрана в кластери) транскрибуються РНК-полімеразою ІІІ. Продуктом транскрипції генів є довші молекули попередники, деякі містять інтрон у межах майбутньої антикодонової петлі. Процесинг цих попередників з утворенням зрілих тРНК полягає у відщепленні певними нуклеазами зайвих фрагментів на кінцях, сплайсингу інтрона (здійснюється специфічними ендонуклеазами талігазою), приєднанні до 3′-кінця стандартного триплету ССА (у складі прокаріотичних тРНК-попередників цей триплет уже присутній), хімічній модифікації певних азотистих основ. Прокаріотичні гени тРНК (87 у геномі E. coli, не містять інтронів) або транскрибуються окремо (як у еукаріотів), або є частинами оперонів, і в цьому випадку первинний транскрипт містить кілька майбутніх молекул тРНК. Крім того, деякі прокаріотичні гени тРНК знаходяться у складі оперона генів рибосомної РНК. В усіх випадках тРНК вирізаються з попередників нуклеазами, після чого піддаються хімічним модифікаціям. Одна з нуклеаз, котра бере участь у процесингу тРНК у про- та еукаріотів, – РНК-аза Р – варта особливої уваги. Фермент складається з двох субодиниць, одна з яких є білковою, а друга – молекулою РНК зі складною просторовою структурою. Саме ця РНК-субодиниця має каталітичну активність, тобто РНК-аза Р – рибозим.
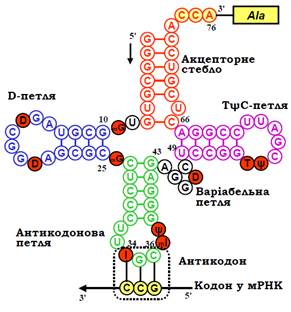
Рис. 1. Схема спарювання нуклеотидів у складі аланінової тРНК.
Схема лист конюшини (рис. 1) не дає уяви щодо просторової структури тРНК. Насправді акцепторне та ТψС-стебло, переходячи одне в одне, утворюють єдину майже пряму подвійну спіраль (рис. 2), під приблизно прямим кутом до якої розташована друга подвійна спіраль, сформована D- і антикодоновим стеблом. ТψС- і D-петлі при цьому наближаються одна до одної, між ними реалізується комплементарне спарювання основ. У результаті молекула тРНК має Г-подібну (або L-подібну) форму з двома плечами різної довжини: на кінці одного плеча акцептується амінокислота (акцепторне плече), на кінці іншого розташований антикодон (антикодонове плече).
У складі дволанцюгових спіральних зон тРНК міститься ~55 % нуклеотидів, але ~90 % основ залучено до стекінг-взаємодій. Отже, молекула характеризується високою впорядкованістю, жорсткістю своєї структури. Це стосується також і петель, серед яких лише антикодонова петля не залучена до взаємодій з іншими елементами. Але п’ять основ петлі (включаючи антикодон) утворюють досить жорстку стопку. Додатково структура тРНК стабілізується іонами Mg2+, тільки у присутності яких молекула є функціонально активною.
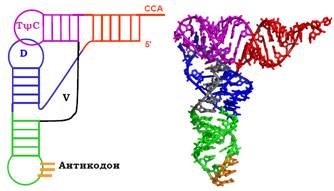
Рис. 2. Схема і просторова структура тРНК (тРНКPhe, 6TNA).
Елементи структури пофарбовані однаково ліворуч і праворуч.
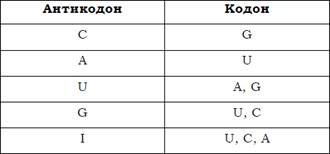
Загальна кількість типів тРНК, які обслуговують процес білкового синтезу, є близькою до 40 (наприклад, усі гени тРНК людини можна поділити на 49 родин за властивостями антикодонів). Оскільки типів тРНК більше, ніж амінокислот, одній амінокислоті може відповідати кілька тРНК – такі тРНК називають ізоакцепторними. Серед них є такі, що містять різні (але, звичайно, синонімічні) антикодони, – гетерокодонові. Є також гомокодонові тРНК, які можуть бути продуктами різних генів (розрізняються за послідовністю нуклеотидів), а можуть бути продуктами одного гена, розрізняючись модифікаціями основ. Оскільки типів тРНК менше, ніж кодонів, одна тРНК здатна впізнавати кілька синонімічних кодонів, що забезпечується неоднозначністю спарювання між першою позицією антикодона і третьою (за якою, головним чином, розрізняються синонімічні кодони) – кодона. А саме, U і G здатні впізнавати по два нуклеотиди у третій позиції кодона, I (який досить часто зустрічається в першій позиції антикодона) упізнає три нуклеотиди (табл. 1).
Таблиця 1.
Відповідність між нуклеотидами в першій позиції антикодона і третій позиції кодона
Аміноацилювання тРНК
Порядок залучення амінокислот до поліпептидного ланцюга, що утворюється при білковому синтезі, залежить лише від взаємодій між нуклеїновими кислотами – кодоном і антикодоном; амінокислота, яку несе тРНК, жодним чином не розпізнається рибосомою. Отже, акцептування певної амінокислоти молекулою тРНК відповідного типу (і тільки відповідного) є одним із найважливіших моментів білкового синтезу: від точності процесу акцептування буде залежати й точність синтезу білка в цілому.
Процес приєднання амінокислот до тРНК каталізується аміноацил-тРНК-синтетазами (АРС-аза, aaRS – aminoacyl-tRN- Synthetase). Кожна з 20 типів (за кількістю амінокислот) цих ферментів каталізує дві хімічні реакції (рис. 3):
• На першій стадії відбувається так зване активування амінокислоти – її приєднання до АМР з утворенням аміноациладенілату, коли пірофосфат (рр) у складі АТР замінюється на амінокислоту (аа). Активування амінокислоти супроводжується зниженням вільної енергії, але при цьому значна частина вільної енергії, що звільняється при відщеплені пірофосфату від АТР, заощаджується у формі аміноациладенілату – молекули, гідроліз якої також супроводжується великим енергетичним ефектом. Молекула АТР, що використовується на етапі активування амінокислоти, – єдине джерело енергії для майбутнього синтезу пептидного зв’язку на рибосомі.
• Аміноациладенілат утворює проміжний комплекс з активним центром ферменту й ефективно атакує ОН-групу рибози 3′-кінцевого аденозину тРНК (3′- або 2′- ОН-групу залежно від класу АРС-ази): відбувається перенесення амінокислоти на тРНК. Після дисоціації від АРС-ази у складі аа-тРНК має місце спонтанний обмін амінокислоти між двома ОН-групами рибози; при зв’язуванні з рибосомою амінокислота фіксується на 3′ ОН-групі. Аа-тРНК – також макроергічна сполука: руйнування зв’язку між амінокислотою та тРНК є енергетично вигідним, що й забезпечує утворення пептидного зв’язку на рибосомі.
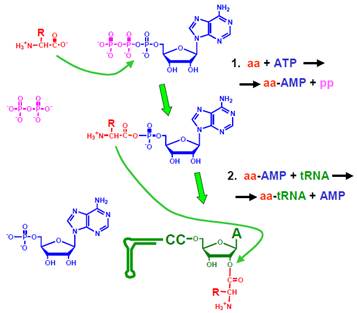
Рис. 3. Приєднання амінокислоти до тРНК, яке каталізується АРС-азою.
Отже, АРС-ази не тільки залучають амінокислоти до білкового синтезу, приєднуючи їх до тРНК, а й забезпечують заощадження вільної енергії, необхідної для приєднання амінокислоти до поліпептидного ланцюга. Двадцять типів АРС-аз, між якими немає майже нічого спільного на рівні первинної структури, можна поділити на два класи, по десять у кожному (табл. 2). Це завжди мультидоменні білки зі складною структурою (рис. 4), що зумовлено розмаїттям функцій – необхідністю специфічно зв’язати три субстрати й каталізувати дві хімічні реакції.
Крім того, еукаріотичні АРС-ази різних типів взаємодіють між собою та з мембраною ендоплазматичного ретикулуму, утворюючи так звану кодосому, що розташована поблизу від рибосом. Мультисубодиничні АРС-ази (гомодимери, гомотетрамери або гетеротетрамери) містять два ідентичні набори активних центрів та сайтів зв’язування.
Таблиця 2.

