




Зрительные рецепторные клетки — палочки и колбоч-ки —"расположёны во внутренней оболочке глаза-— сет-чатЕЕГ~Самый наружный слой сетчатки образТШн т!иг-" ментным эпйтеТтеТС~к~"~котот;о1^^
прГмыкает слой фоторецёпторов. Каждая палочка "и
колбочка состоит из наружного сегмента, КРТО'рыи имеет
соответственно палочкообразную или кол бочкообразную
форму, и внутреннего сегмента, содержащего ядро и ми
тохондрии обеспечивЖПГГЯе энергетические процессы, в"
фоторецепторнои клетке! ' '
Кнутри от слоя фоторецепторных клеток расположен слой биполярных нейронов, с которыми контактируют рецепторные клетки. Биполярные нейроны соединены с ганглиозными клетками, отростки которых составляют волокна зрительного нерва. Возбуждение, возникающее вфоторецепторе под действием света, передается на волокна зрительного нерва через биполярную и ганглиоз-'
ную клетки.
Электронно-микроскопические исследования показали, что наружные сегменты фоторецепторов—имеют слоистую структуру, представляя собой как бы стопку дисков, число которых достигает неаскольких сотен. Каж-дый диск состоит из сдвоенной белково-липидной мем-браны, соединенной по краям (рис. 59). Диски образуются путем впячивания клеточной мембраны внутрь сегмента и представляют собой образования, аналогичные кристам митохондрий. В результате такого процесса междисковые промежутки соответствуют цитоплазме
20 Медицинская биофизика
305

клетки и имеют высокое содержание калия, а внутриди-сковые промежутки являются замкнутыми участками внешней среды, богатой натрием. Следовательно, наружные сегменты фоторецепторов представлены структурами, обусловливающими многократно повторяющийся градиент концентрации К+ и Ка+, который поддерживается процессами активного переноса с участием Ка+—К+ — АТФ-азы. За счет образования дисков эф-фективная светочувствительная поверхность фоторецеп-тора возрастает во много раз, что обусловливает высокую вероятность ее взаимодействия с квантами света.
Исследования, выполненные в последнее десятилетие с помощью микроэлектродной техники, показали, что при действии светя в фоторецепторах формируются генераторные потенциалы1, величина которых пропорциональна интенсивности действующего света. В след за появлением генераторных потенциалов возникают импульсы в нервных волокнах. Большинство авторов в настоящее время считают, что передача возбуждения с рецепторных клеток на биполярные осуществляется с помощью холинергической системы. Суммарное изменение разности потенциалов в сетчатке при действии света — электроретинограмму (ЭРГ) — можно зарегистрировать с помощью электродов, приложенных непосред-
1 Многие авторы называют их рецепторными 'потенциалами, а генераторными обозначают потенциалы в нервных окончаниях.
306
ственно к сетчатке или к неповрежденному глазу. Только в самое последнее время в ЭРГ удается выделить компоненты, обусловленные электрическими изменениями в отдельных группах клеток сетчатки.
В настоящее время твердо установлено, что восприя-тие света начинается с поглощения света молекулами светочувствительных пигментов — белков, находящихся в наружных сегментах палочек и колбочек. Все светочувствительные белки являются хромопротеидами. Они состоят из полиенового хромофора ретиналя — альдегида витамина А — и молекулы белка опсина (полнен — углеводородная цепь с сопряженными двойными связями). Таким образом, все зрительные пигменты содержат одну и ту же группу — ретиналь — и отличаются химическим составом и структурой своей белковой части — опсином. Согласно последним данным, практически весь белок фоторецепторной мембраны представлен опсином (92-95%).
К настоящему времени в палочках всех видов животных обнаружен зрительный пигмент родопсин а в кол бочках многих животных найдены пигменты йодопсин, цианопсин и др. Наиболее изученным в настоящее время является родопсин. Биохимические и электронно-микроскопические данные позволяют предполагать, что молекулы родопсина покрывают наружную поверхность дисков с обеих сторон и составляют около половины всех белков наружного сегмента. В последнее время допускают возможность его локализации и на внутренней поверхности дисков.
Соединение опсина с ретиналем осуществляется только в том случае, если последний находится в виде «изогнутой» 11-цис-формы (см. рис. 59). Незначительное изменение в структуре хромофора, например удлиненение полиеновой цепи, уже препятствует образованию родопсина. Показано, что опсин соединяется с ретиналем путем взаимодействия альдегидной группы ретиналя и аминогруппы одной из аминокислот опсина. Современные методы исследования, в частности микроспектрофо-тометрия, показали, что молекул? родопсина ориентирована в мембране рецептора так, что ее хромофор — ретиналь— в 90% случаев лежит перпендикулярно направлению светового луча, т. е. перпендикулярно длин-ной оси палочки. Остальные 10% молекул располагаются параллельно оси палочки.
20* 307
Впервые фотохимические процессы в рецепторных клетках были исследованы Уолдом. Применялись спект-рофотометрические методы, метод электронного парамагнитного резонанса и биохимические методы. Уолдом было показано, что элементарный зрительный акт начинается с поглощения молекулой пигмента кванта электромагнитного излучения. При этом электроны в молекуле ретиналя переходят с основного синглетного уровня на возбужденный. В результате поглощения энергии кванта происходит процесс изомеризации: ретиналь из «изогнутой» 11-цис-формы превращается в «выпрямлен-ную» полностью транс-форму. Углеводородная цепь
полиостью выпрямляется и все части молекулы располагаются по обе стороны оси 11 — 12. В результате трансизомеризации ретиналь не может «вкладываться» в соответствующую «нишу» опсина, что приводит к ослаблению связи между молекулой ретиналя и опсином. Ко-нечным результатом реакции фотоизомеризации являет-ся распад родопсина на ретиналь и опсин. При этом происходит выцветание родопсина — он из розового становится желтоватым, а затем бесцветным.
Реакция изомеризации 11-цис-ретиналя полностью в транс-форму идет через ряд промежуточных стадий, в ходе которых образуются изомеры ретиналя, устойчивые только при низких температурах (пре-люмиродопсии, —140 °С; люмиродопсин, —40 °С; мета-родопсин, —15 °С и др.).
Распад родопсина на ретиналь и опсин приводит к возбуждению рецепторной клетки. В последующих ста-диях осуществляется подготовка к следующему восприятию квантов света. При этом освободившийся ретиналь вначале восстанавливается в ретинол — одну из форм
витамина А.
Ретинол под действием фермента ретиненизомеразы через ряд стадий, которые еще слабо изучены, вновь превращается в 11-цис-изомер.
Необходимо отметить, что в процессе фотохимических реакций родопсина происходят не только изменения структуры ретиналя, но и существенные изменения структуры белковой части молекулы — опсина.
В последнее время многими авторами было показано, что превращения йодопсинаколбочек на свету и в тем-ноте в общем совпадают с превращениями родопсина палочек.
308
Каким же образом молекулярные превращения зрительного пигмента в той или другой стадии его распада приводят к возникновению генераторного потенциала рецепторной клетки? Вопрос о механизме возбуждения зрительных рецепторов еще не получил своего оконча-тельного решения, так же как не выяснены механизмы возбуждения других рецепториых клеток. В настоящее время данная проблема является одной из наиболее важных в биофизике органон чувств. Несомненно, что ее решение будет возможным в результате глубокого изучения молекулярных процессов, происходящих в мембранах рецепториых клеток при действии внешних стимулов.
Из всех теорий, объясняющих механизм возбуждения зрительных рецепторов при распаде зрительного пигмента, основного внимания заслуживают две: энзиматиче-ская (Уолд) и ионная (Бонтинг, Бегем).
Первая теория исходит из представления, что родопсин является проферментом. Активизация фермента осуществляется воздействием фотона в результате освобождения активного центра, который в темноте «прикрыт» молекулой ретиналя. Опсин проявляет свою активность, что приводит к возникновению генераторного потенциала. При этом предполагается, что родопсин или сам обладает АТФ-азными свойствами, или связан с АТФ-азой.
Вторая теория основывается на ряде уже приводившихся данных о наличии ионных градиентов между ре-цепторными клетками и окружающей их средой и о природе связи между молекулами ретиналя и опсина. Согласно данной теории, вход натрия в наружные сегменты фоторецепторов и выход из них калия при освещении протекает на основе тех же закономерностей, которые установлены для возбуждения нерва и мышцы. Предполагается, что действие света приводит к распаду родопсина. Освобождающийся полностью транс-изомер ретиналя реагирует с положительно заряженными аминогруппами фосфолипидов мембраны фоторецептора. Блокирование аминогрупп фосфолипидов делает мембрану более отрицательно заряженной, что приводит к резкому повышению проницаемости мембраны для противоположно заряженных катионов. Повышение ионной проницаемости мембраны вызывает появление генераторного потенциала, который активирует холинергиче-
309
ский механизм в синапсе и вызывает возбуждение нервных клеток. В фазе восстановления ретиналь освобождается от фосфолипида, реизомеризуется в цис-форму и соединяется с опсином. Ионные градиенты восстанавливаются в результате процессов активного переноса ионов.
Хагинс предполагает, что необходимое для возбуждения изменение проницаемости мембраны может быть вызвано не только описанными процессами изменения заряда мембраны, но и изменением конформации молекул светочувствительных белков. Таким образом, молекулярные превращения зрительного пигмента могут рассматриваться как пусковой механизм для возбуждения фоторецепторов.
Кодирование информации в органе зрения
При исследовании процессов кодирования зрительной информации необходимо решить несколько задач: каким образом кодируется длина волны и интенсивность света, а также какие принципы лежат в основе кодирования информации о размерах и форме предметов.
Кодирование длины волны света. Способность глаза по разному воспринимать свет различной длины волны называется цветовым зрением. Еще в конце прошлого века было установлено, что палочки сетчатки являются рецепторами системы монохроматического (черно-белого, или серого) зрения, а колбочки — рецепторами системы полихроматического (цветового) зрения.
Наибольшим признанием пользуется трехкомпонент-ная теория цветового зрения. Эта теория предложеная
еще М.В. Ломоносовым, была разработана в прошлом столети Юнгом и Гельмгольцем. Согласно этой теории, колбочки сетчатки делятся на три вида и содержат раз-личные светочувствительные веществ. Всякий цвет ока-зывает действие на все три вида рецепторов, но в раз-
личной степени. При изолированном возбуждении колбочек одного вида возникло бы ощущение насыщен-ного красного, при изолированном возбуждений другого -- насыщенного зеленого, а при изолированном возбуждение третьего -- насыщенного синего. Если одновременно возбуждаются два вида рецепторов, то возни-
310
|
|

каст ощущение промежуточного цвета. Например, при возбуждении рецепторов зеленого «и синего цвета возникает ощущение голубого цвета. При одновременном возбуждении всех видов ре-цепторов возникает ощуще-ние белого или серого цве-та. Таким образом, согласно данной теории, кодирование длины волны света обусловлено наличием фоторецепторов, обладающих избирательной чувствительностью к электромагнитным колебаниям определенной
длины волны. Все многообразие цветовых ощущений обусловлено соотношением количества возбуждаемых рецепторов разных видов.
В последнее время трехкомпонентная теория получила много экспериментальных подтверждений благодаря применению электрофизиологических, спектрофотомет-рических, биохимических и других методов. Поэтому в настоящее время большинство ученых признают наличие в сетчатке трех видов колбочек.
Так, Гранит, исследуя чувствительность рецепторов глаза, установил, что есть рецепторы, имеющие максимум чувствительности в желтой области спектра, и рецепторы с максимумом чувствительности в красной, зеленой и синей областях спектра (рис. 60). Он прикладывал к обнаженной сетчатке кошек и лягушек микроэлектроды, позволявшие отводить токи действия от отдельных волокон зрительного нерва или от небольших групп таких волокон. Этим методом было обнаружено, что как бы ни перемещался электрод по исследуемой сетчатке животных, обладающих цветовым зрением, получаются кривые чувствительности одного из четырех типов. Первый из них дает колоколообразную кривую, охватывающую весь видимый спектр и имеющую максимум около 560 нм. Остальные три кривые имеют также колоколообразную форму, но охватывают ограниченные области спектра, имея максимумы в красном, зеленом и синем участках спектра. Считают, что первая кривая
311
чувствительности соответствует палочковому, монохроматическому, зрению, а остальные три — колбочковому, цветному, зрению.
Существование в одной сетчатке колбочек, обладающих различной спектральной избирательностью поглощения, было подтверждено также для птиц, черепах, ящериц и насекомых. При этом было выяснено, что у позвоночных нет морфологических отличий между различными видами колбочек. Но такие отличия были обнаружены у насекомых с полихроматическим зрением. Например, по данным Ф. Г. Грибакииа (1971), у рабочей пчелы имеются три типа морфологически отличающихся рецепторов с максимумами чувствительности в ультрафиолетовой, желто-зеленой и синей областях спектра.
Различия в спектральной чувствительности колбочек объясняются различиями в светочувствительности зрительных пигментов колбочек. Учитывая, что структура и свойства молекулы ретиналя неизменны, все разнообразие зрительных пигментов следует отнести за счет структурных особенностей белковой части пигментов — опсина. В 1959 г. Раштон обнаружил в сетчатке человека два светочувствительных пигмента с максимумами поглощения 540 и 590 нм. Стайлз получил три спектральные кривые с явно выраженными максимумами поглощения — 440, 540 и 590 нм, что, по его мнению, соответствует трем типам фоторецепторов.
Трехкомпонентная теория объясняет большинство фактов из физиологии и патологии цветного зрения. Например, Уолд феномены цветовой слепоты объясняет тем, что у протанопов не работают красночувствитель-ные рецепторы, у тританопов — синечувствительные, у дейтеранопов — зеленочувствительные или красночувст-вительные, объединенные в один механизм.
Некоторые авторы придерживаются мнения о наличии в сетчатке не трех, а большего числа типов рецепторов. Так, Гранит и Хартридж допускают, что в сетчатке имеется семь типов рецепторов, реагирующих на различные цвета. Гранит считает, что три кривые чувствительности, полученные при отведении потенциалов сетчатки, формируются в результате усреднения кривых спектральной чувствительности семи типов рецепторов.
Кодирование информации об интенсивности света. Кодирование информации об интенсивности света в ор-
312
гане зрения основано на тех, же принципах, что и кодирование информации об интенсивности раздражителей в других органах чувств. Как указывалось, при действии света в фоторецепторных клетках возникают, гене-
раторные потенциалы, величина которых пропорциональна
интенсивности действующего света. Генераторные по-_ тенциалы вызывают возбуждение биполярных клеток и волокон зрительного нерва. Частота возбуждения нерв - ных клеток пропорциональна величине генераторных потенциалов и, следовательно, интенсивности света. При этом между интенсивностью света и частотой нервных импульсов в волокнах зрительного нерва в определен-ных пределах имеется логарифмическая зависимость, выраженная уравнением 4 (стр. 286). Впервые это было показано Хартлайном и Грэмом на глазу мечехвоста.
Кодирование информации о расстоянии до предметов и о величине предметов. Оценка расстояния до предмета возможна как при зрении одним глазом (монокулярном), так и при зрении двумя глазами (бинокулярном). При монокулярном зрении основное значение в оценке расстояния имеет явление аккомодации, которое заключается в изменении кривизны хрусталика для получения четкого изображения на сетчатке. Кодирование расстоя-ния до предмета в данном случае осуществляется сте-пенью напряжения ресничной мышцы, изменяющий кри-
визну хрусталика. Для оценки расстояния имеет значе-ние и величинаизображения предмета на сетчатке.
При бинокулярном зрении основное значение в оценке расстояния имеет явление диспарации — расхождения изображений предметов на сетчатке глаз. При бинокулярном зрении, когда глаза конвергированы (сведены) на предмет, изображения всех его точек попадают на идентичные участки сетчатки обоих глаз и в представлении человека два изображения сливаются в одно, В то же время изображения предметов более удаленных и менее удаленных, чем тот, на который глаза конвергированы, попадают уже на неидентичные участки сетчатки и их изображения представляются раздвоенными. В зависимости от величины диспарантного расхождения изображении предметов на сетчатке возникают ощуще-ния большей или меньшей удаленности предметов друг от друга, т. е. глубина рельефа кодируется величиной диспарации. При бинокулярном зрении точность оценки расстояния значительно выше, чем при монокулярном.
313
Информация о величине предметов кодируется количеством возбуждаемых рецепторов. В зависимости от величины предметов на сетчатке будет возникать изо-бражение большей или меньшей величины и станет возбуждаться больше или меньше количество рецепторов. А так как величина изображения предмета на сет-чатке зависит от расстояния до предмета, то оценка величины предмета будет зависеть и от этого расстояния.
Вся зрительная информация, которую может получить мозг, содержится в распределении активности меж-ду фоторецепторами сетчатки (за исключением информации, представленной степенью напряжения мышц, участвующих в зрении). Но эта информация завуалиро-вана общим возбуждением рецепторов. Такая неявная информация становится явной в последующих стадиях переработки. Одним из механизмов переработки информации на уровне сетчатки является латеральное торможение, впервые изученное в сложном глазу мечехвоста.
Было обнаружено, что между омматидиями (рецепторами) глаза существуют взаимотормозные отношения. Тормозной эффект, оказываемый каждым омматидием на другие клетки, пропорционален его возбуждению и убывает с расстоянием между ними. Роль латерального торможения состоит в том, что оно обеспечивает контрастность и контурность изображения.
Впоследствии латеральное торможение было изучено в органах зрения других животных и человека. При латеральном торможении функциональную связь между рецепторами, биполярными и ганглиозными клетками осуществляют горизонтальные и амакриновые клетки сетчатки.
Латеральное торможение играет важную роль и при цветовосприятии. Окончательно механизмы латерального торможения в глазу человека еще не выяснены. По мнению Сомьена из трех типов колбочек два всегда реци-прокно связаны с одними нейронами, одна совокупность — возбуждающими, а другая — тормозными синапсами. За счет этого ганглиозные клетки обладают более высокой спектральной избирательностью, чем рецепторы, и сигналы от тех колбочек, которые не являются специфичными для данного света, не доходят до центральной нервной системы.
314
Глава 14 БИОФИЗИКА ВНЕШНЕГО ДЫХАНИЯ
Внешнее дыхание, или вентиляция легких, заключается в обмене воздуха между внешней средой и альвеолами. Вентиляция легких осуществляется в результате ритмических дыхательных движений грудной клетки.
Задачи биофизики внешнего дыхания заключаются главным образом в выяснении и описании связей между работой дыхательных мышц, давлением в разных частях аппарата дыхания, объемом легких и движением воздуха.
К настоящему времени разработан ряд методов, которые могут дать количественные сведения о механизме внешнего дыхания. Разработанные тесты получили широкое распространение не только в физиологии, но и клинике. Определение физических параметров внешнего дыхания помогает диагностировать многие заболевания и оценивать эффективность терапии.
БИОМЕХАНИКА ВДОХА И ВЫДОХА (ФИЗИЧЕСКИЕ ОСНОВЫ ВЕНТИЛЯЦИИ ЛЕГКИХ)
| (1) (2) |
Механика вдоха. Легкие, помещающиеся внутри грудной клетки, отделены от ее стенок плевральной полостью (щелью). В грудной клетке они находятся в растянутом состоянии. За счет того что легкие обладают эластичностью (эластичность — сочетание растяжимости и упругости), давление в плевральной полости1 ниже, чем в альвеолах. В условиях равновесия давление в альвеолах Рал уравновешивается суммой давления б плевральной полости Рпл и давления Рэл.л, создаваемого эластической тягой легких:
|
|

Отсюда
1 Здесь и далее, говоря о плевральном давлении или плевральной полости, имеется в виду давление в межплевральном щелевидном пространстве или само это пространство.
315
т. е. плевральное давление меньше альвеолярного на величину, обусловленную эластической тягой легких.
Атмосферное давление на грудную клетку Ратм уравновешивается суммой плеврального давления и давления Рэп.г.к, создаваемого эластичностью грудной клетки:
|
|

(3)
Плевральное давление, следовательно, меньше атмосферного на величину, развиваемую эластической тягой грудной клетки. Плевральное давление поэтому часто называют отрицательным, принимая уровень атмосферного давления за нуль.
Отрицательное давление в плевральной полости стремится сжать грудную клетку. Стало быть, тяга грудной клетки направлена в противоположную от тяги легких сторону (что облегчает вдох). Соотношение указанных сил определяет уровень спокойного дыхания и функциональную остаточную емкость. Когда глубина вдоха становится выше 70% жизненной емкости, эластичность грудной клетки начинает противодействовать вдоху и ее тяга уже направлена в ту же сторону, что и эластическая тяга легочной ткани.
Акт вдоха совершается в результате увеличения объема грудной полости, происходящего при подъеме ребер и опускании диафрагмы. Диафрагма является самой сильной мышцей вдоха, обеспечивающей примерно 2/з вентиляции. При сокращении она уплощается и объем грудной полости увеличивается в вертикальном направлении. Приподнимание ребер обеспечивается главным образом сокращением наружных межреберных мышц. Эти мышцы при сокращении должны были бы сближать ребра, так как сила, прилагаемая к точкам прикрепления на верхнем и нижнем ребрах, одинакова. Но при косом расположении мышц плечо и момент силы у верхнего ребра всегда меньше, чем у нижнего. Поэтому при сокращении мышц ребра в целом поднимаются, что увеличивает сечение грудной клетки как в передне-заднем, так и в поперечном направлении.
Увеличение объема грудной клетки при сокращении вдыхательных мышц приводит к уменьшению давления в плевральной полости. В результате этого воздух в легких, преодолевая сопротивление их растяжению, расширяется, а его давление в соответствии с законом Бой-
316
ля — Мариотта уменьшается и становится ниже атмосферного. Возникновение разности давлений между атмосферой и альвеолами легких служит непосредственной причиной движения в них воздуха.
•
Объемная скорость тока воздуха V, представляющая собой первую производную от объема проходящего через дыхательные пути воздуха по времени, описывается уравнением:
|
|

(4)
где ΔР — разность между Ратм и Рал; Rв — аэродинамическое сопротивление, обусловливаемое трением движущегося воздуха о стенки дыхательных путей и внутри
воздушного потока. V представляет собой объем воздуха, проходящего по дыхательным путям за единицу времени, и измеряется обычно в л/с. Объемы вдыхаемого и выдыхаемого воздуха легко измеряются, например, спирометром. Изменения же объемов циркулируемого воздуха во времени записываются спирографом; возникающая при этом кривая называется спирограммой. Объемная скорость воздуха измеряется пневмотахометром, а ее изменения во времени регистрируются пневмотахографом в виде кривой — пневмотахограммы.
Механика выдоха. Во время вдоха вдыхательные мышцы преодолевают ряд сил: 1) эластическое сопротивление грудной клетки (после достижения 70% жизненной емкости) и внутренних органов, отдавливаемых книзу диафрагмой; 2) эластическое сопротивление легких; 3) динамическое (вязкостное) сопротивление всех перемещаемых тканей; 4) аэродинамическое сопротивление дыхательных путей; 5) тяжесть перемещаемой части грудной клетки; 6) силы, обусловленные инерцией перемещаемых масс.
Энергия мышц, затраченная на преодоление всех видов динамического сопротивления (обусловленного трением), переходит в тепло и в дальнейшем процессе дыхания не участвует. Остальная часть энергии мышц вдоха переходит в потенциальную энергию растяжения всех эластических тканей и потенциальную энергию тяжести. При расслаблении вдыхательных мышц под действием эластических сил грудной клетки и внутренних органов и силы тяжести грудной клетки ее объем уменьшается. При форсированном выдохе к перечисленным
317
силам присоединяется сокращение внутренних межреберных мышц, задних нижних мышц и мышц живота.
Уменьшение объема грудной клетки приводит к по-вышению плеврального давления. В результате этого и под действием эластической тяги легких воздух в альвеолах сжимается, его давление становится выше атмосферного и он начинает выходить наружу. Когда эластическая тяга легких уравновесится понижающимся давлением в плевральной полости, выдох закончится. В легких остается некоторое количество воздуха, которое называют остаточным объемом. Таким образом, действие дыхательных мышц на легкие осуществляется не непосредственно, а через изменение давления в плевральной полости. Непосредственной же причиной движения воздуха через дыхательные пути являются циклические колебания альвеолярного давления.
Если вскрыть грудную клетку — произвести пневмоторакс, то тем самым будет выключен один из важных факторов механики дыхания; тогда стремление легких к спадению за счет эластической тяги проявится в полной мере — легкие займут так называемый объем коллапса, который значительно меньше, чем остаточный объем. Вентиляция легких при этом становится невозможной, поскольку при изменении объема грудной клетки воздух движется не через дыхательные пути, а через искусственно созданное отверстие в стенке грудной клетки.
РАСТЯЖИМОСТЬ ЛЕГКИХ
Исследование эластичности легких началось с работ Гутчинсона (1852) и Дондерса (1853). Эти исследователи, меняя давление воздуха в трахее и одновременно измеряя объем воздуха в легких (либо изолированных, либо при широко вскрытой грудной полости), получили первые данные об эластичности легких. В настоящее, время зависимость между давлением и объемом легких можно считать хорошо установленной. Как оказалось, она значительно отличается от таковой для идеально упругого тела. Объем идеально упругого полого тела, под-чиняющегося закону Гука, линейно возрастает при повышении давления внутри тела. Эластическое сопротив-ление такого тела характеризуется величиной растяжи-
318
|
|

мости С, которая равняется отношению изменения объема Δ V к изменению давления Δ Р:
|
|

(5)
В отличие от такой линейной зависимости для легких характерны S-образные кривые, различные в зависимо-сти от последовательности изменений давления. Если через вставленную в трахею спавшихся легких трубку постепенно повышать в них давление, то вначале объем легких увеличивается незначительно, затем быстро воз-растает и при приближении к предельному объему вновь маломеняется (рис. 61,1). Если теперь постепенно по-нижать давление, то объемы легких при тех же величинах давления, при которых проводился первый опыт, окажутся иными, значительно большими.
Отклонение поведения легких от поведения идеально упругих тел — нелинейность зависимости V — Р и наличие петли гистерезиса — обусловлено тем, что эластическая тяга легких обусловлена двумя факторами: 1)наличием в альвеолах эластических волокон и 2) поверхностным натяжением пленки жидкости, покрывающей внутрен-нюю поверхность альвеол. На долю поверхностного натяжения приходится 55—65% эластической тяги легких. В соответствии с законом Лапласа давление, создаваемое поверхностным натяжением, обратно пропорцио-
319
нально радиусу сферы. Поэтому мало заполненные воздухом альвеолы имеют высокое сопротивление, а с уве-личением объема альвеол их сопротивление уменьшается. Поэтому же легкие с малым исходным объемом трудно заполнить воздухом, но раздутые легкие сохраняют большой объем даже при снижении давления. Устранение поверхностного натяжения альвеол путем заполнения их не воздухом, а физиологическим раствором дает три эффекта: 1) растяжимость легких значительно повышается, т. е. при одинаковом давлении они занимают больший объем; 2) зависимость V — Р становится близкой к линейной; 3) гистерезис почти полностью устраняется (рис. 61).
Однако в своем первоначальном виде гипотеза поверхностного натяжения альвеол встретила серьезные возражения. Расчеты, основанные на законе Лапласа, показали, что при микроскопических размерах альвеол должно возникать столь большое поверхностное натяжение, что оно приводило бы к спадению альвеол. Для их расправления разность между альвеолярным и плевральным давлением должна была бы быть в несколько раз больше, чем реально существующая, а работа вдоха также должна была бы превышать нормальную в несколько раз. Высокое поверхностное натяжение приводило бы также к нестабильности альвеол. Так как альвеолы соединены между собой параллельно, то воздух из малых альвеол должен поступать в большие; последние должны еще больше увеличиться, а малые — спасться.
Описанные несоответствия были объяснены после того, как обнаружили, что клетки альвеолярного эпителия выделяют поверхностно-активное вещество — сурфак-тант, который снижает поверхностное натяжение. По своей природе он оказался липопротеидом. Внутренняя поверхность альвеол покрывается пленкой сурфактаита, который стабилизирует силы поверхностного натяжения. При уменьшении объема альвеол плотность молекул сурфактанта на поверхности альвеол увеличивается, что снижает коэффициент поверхностного натяжения и ограничивает действие закона Лапласа. В результате этого устраняется опасность ателектазов (спадения альвеол).
Исследование поверхностного натяжения легких имеет большое практическое значение, поскольку многие
320
заболевания дыхательной системы могут быть вызваны или сопровождаться изменениями поверхностного натяжения. Обнаружено, что в легких нежизнеспособных новорожденных, погибших от асфиксии из-за невозможности дыхания, снижено содержание сурфактанта. Установлено также, что его содержание в легких падает после ваготомии, прекращения кровотока по легочной артерии, при повышенном парциальном давлении кислорода.
СОПРОТИВЛЕНИЕ ДЫХАНИЮ
Эластическое сопротивление. Эластическим сопротивлением, наз. сопротивления которое должны преодолеть дыхательные мышцы при вдохе, является упруго-эластическая сила аппарата внешнего дыхания. Преодолеваемые эластические силы разделяются на два вида: 1) эластические силы легких; 2) эластические силы грудной клетки (и внутренних органов, смещаемых диафрагмой). Эластические силы, развиваемые легкими и грудной клеткой, зависят от их объемов и не зависят от скорости изменения этих объемов. Измерение растяжимости должно производиться в статических условиях, так как динамическое сопротивление ткани и воздухоносных путей и силы инерции равны нулю только при отсутствии тока воздуха. Для исключения динамического компонента сопротивления используют различные приемы.
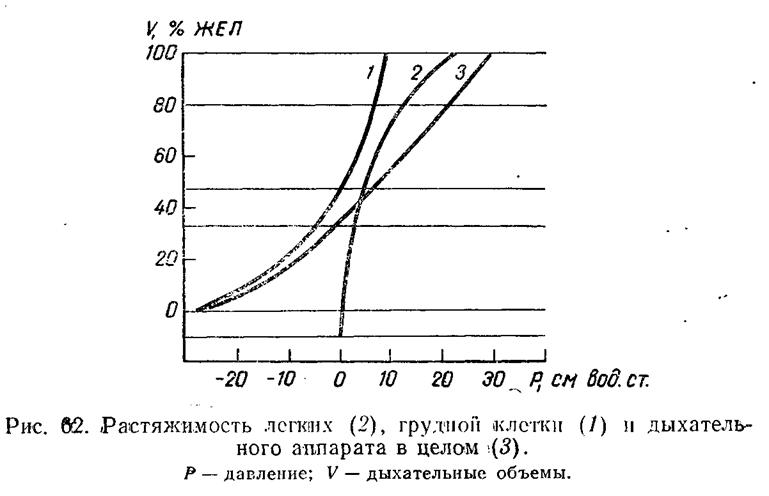
Эластические свойства дыхательного аппарата в целом и отдельных его частей исследуют различными методами. Наиболее распространенный из них заключается в следующем. Испытуемый вдыхает измеренный объем газа из спирографа, после чего трубку, соединяющую верхние дыхательные пути со спирографом, перекрывают зажимом. Затем испытуемый полностью расслабляет дыхательную мускулатуру. Теперь эластической тяге легких и грудной клетки не противодействует активное сокращение мышц. Эта тяга создает альвеолярное давление, которое измеряется манометром, присоединенным к трубке между дыхательными путями и зажимом. Повторение этой процедуры несколько раз при разных объемах дает возможность построить кривую V — Р (рис. 62).
Другой способ заключается в том, что легкие наркотизированного больного раздувают через интратрахе-альную трубку, плотно прилегающую к стенкам трахеи.
Медицинская биофизика
321
Затем трубку закрывают и измеряют давление в системе трубка — легкие. Повторяя процедуру несколько раз и измеряя объемы газа, устанавливают зависимость V
Ои Рал.
В среднем диапазоне изменений объема зависимость V — Р ал оказалась близкой к линейной, что позволило использовать уравнение (5) и характеризовать эластические свойства аппарата дыхания в целом одним параметром — растяжимостью. Последняя у здорового человека близка к 0,1 л/см вод. ст. Это означает, что для обеспечения спокойного вдоха (0,5 л) мышцы должны развить силу, создающую в легких отрицательное давление 5 см вод. ст.
Растяжимость дыхательного аппарата бывает уменьшенной при определенных заболеваниях (фиброз легких и грудной клетки, легочный застой и др.). Указанным можно объяснить те трудности, которые испытывают некоторые больные при вентиляции своих легких.
Для клиники представляет интерес исследование эластических свойств не только дыхательного аппарата в целом, но и его частей: легких и грудной клетки. Определение изменений их растяжимости при различных патологических состояниях дает возможность устанавливать причины, вызывающие затрудненное дыхание и обусловливающие нарушения частоты, глубины дыхания и структуры дыхательного цикла (Л. Л. Шик, 1973).
322
Из уравнения (1) следует, что для определения растяжимости легких необходимо построить кривую зависимости объема легких от транспульмонального давления, т. е. от разности между альвеолярным и плевральным давлением. Измерять плевральное давление непосредственно опасно из-за возможности прокола легкого. Задача облегчилась после того, как было установлено, что изменения давления в пищеводе равны соответствующим сдвигам его в плевральной полости. Давление в пищеводе измеряют с помощью маленького баллона, соединенного с манометром. Если одновременно регистрировать легочные объемы, альвеолярное давление (по описанной выше методике) и давление в пищеводе, то можно получить данные для построения зависимости объема от транспульмонального давления.
Растяжимость грудной клетки можно определить, если построить кривую зависимости объема легких от
разности между атмосферным и плевральным давлением. Для этого практически необходимо измерять только объемы воздуха и давление в пищеводе, поскольку атмосферное давление постоянно (при кратковременном исследовании).
Измерения показали, что растяжимость легких у здорового человека близка к 0,2 л/см вод. ст. Растяжимость грудной клетки также имеет величину, близкую к 0,2 л/см вод. ст. Таким образом, легкие и грудная клетка имеют примерно одинаковую растяжимость, которая в 2 раза превышает растяжимость дыхательного аппарата в целом.
Динамическое сопротивление. Сопротивления дыха-нию, возникающие исключительно во время дыхательных движений т.е. когда воздухидет в влегкие или из легких называют динамическими или вязкостными со-противлениями. Различают динамическое сопротиваление тканевое и воздушное (или аэродинамическое).
Тканевое динамическое сопротивление возникает в результате трения в тканях, которые премещаются во время дыхания. Это ребра, диафрагма, содержимое борюшнои полости, легкие. В отличие от эластического сбпротивления, определяемого изменением объема, оно определяется скоростью движения. В начале вдоха, когда ток воздуха еще не начался и ткани не перемещаются, эта сила равна нулю. Она становится наибольшей в момент максимальной скорости тока воздуха и вновь
21*
328
равна нулю в конце вдоха, когда ток и перемещение останавливаются. У людей со здоровыми органами дыхания тканевое динамическое сопротивление составляет 15—18% всего динамического сопротивления (Маршалл, 1956).
Аэродинамическое сопротивление. При дыхании требуется давление, чтобы преодолеть трение между стен-ками трахобронхиального дерева и воздушным пото-ком, а также внутри самого потока. Сопротивление ды-хательных путей зависит от объемной скорости и типа воздушного потока. Воздушный поток может быть лами-нарным или турбулентным. В гладких прямых трубках турбулентное течение возникает лишь при больших скоростях. Однако трахеобронхиальное дерево имеет сотни тысяч разветвлений, около которых могут возникать завихрения. Турбулентность, или вихреобразование, при малых скоростях особенно часто возникает там, где в трубках имеются неровности, вызванные, например, слизью, экссудатом, опухолью, инородными телами.
В общем случае давление АР (или разность давлений), необходимое для преодоления аэродинамического сопротивления, определяется уравнением:
|
|
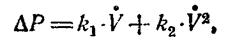
(6)
где k 1 и k 2— коэффициенты, характеризующие аэродинамические сопротивления соответственно для ламинарного и турбулентного потоков воздуха.
Коэффициенты k х и k 2 находят из уравнений:
|
|
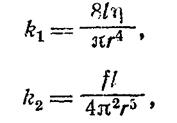
(7)
(8)
где l — длина трубки; r — ее радиус; ή —вязкость газа; f — коэффициент трения, зависящий от плотности газа.
Уравления (6), (7) и (8) показывают, аэроди-намическое сопротивление увеличивается с повышением объемной скорости воздуха, при переходе от ламинарного к турбулентному потоку и особенно при сужении просвета дыхательных путей. При астме и эмфиземе легких это сопротивление может быть в 7—8 раз больше, чем у здоровых людей.
324
Методы измерения динамического сопротивления. Наиболее точным можно считать метод общей плетиз-мографии, позволяющий исследовать одновременно мно-гие параметры внешнего дыхания. Принцип действия общего плетизмографа, или железных легких, заключается в следующем. Тело пациента с расслабленной дыхательной мускулатурой, за исключением головы, заключают в герметический металлический сосуд. В нем благодаря действию мощного насоса ритмически создается отрицательное и положительное давление, воздействующее на все тело. Отрицательное давление приводит к увеличению объема грудной клетки и вызывает вдох; при повышении давления совершается выдох.
В данном случае работа дыхательных мышц заменена действием искусственного насоса.
В опыте одновременно регистрируются разность давлений между плетизмографом и ротовой полостью, спирограмма и пневмотахограмма. Если построить зависимость давления от объема в статических условиях — при отсутствии тока воздуха, то будет получена прямая линия (в первом приближении). Это линия АВ на рис. 63. Построение этой же зависимости в динамических условиях даст кривую АЕВ, которая проходит ниже АВ, поскольку при движении к эластическому присоединяется и динамическое сопротивление. Расстояние между осью абсцисс и АВ в каждый момент показывает давление ΔРэл, расходуемое на преодоление эластического сопротивления, а расстояние между АВ и АЕВ — ΔРД — давление, затрачиваемое на преодоление динамического сопротивления всего дыхательного аппарата. Определяя
ΔРд и V (по пневмотахограмме) для каждого момента
времени, по уравнению  находят динамическое сопротивление Rд.
находят динамическое сопротивление Rд.
Общее динамическое сопротивление легких и дыхательных путей можно определить способом, аналогичным описанному. Только для этого необходимо строить зависимость дыхательных объемов от разницы давлений между пищеводом и ртом, а дыхание может осуществляться как естественным путем, так и с помощью общего плетизмографа.
Для определения одного только динамического сопротивления дыхательных путей надо знать: 1) объем-
325

ную скорость воздуха и 2) разность давлений между альвеолами и атмосферой. Прямое измерение последней величины невозможно, так как в альвеолы нельзя ввести зонд. В связи с этим прибегают к косвенным способам. Наиболее доступным является метод прерывания воздушного потока. Его суть заключается в том, что в любой фазе дыхательного цикла трубка, соединяющая верхние дыхательные пути человека с пневмотахогра-фом, перекрывается на короткий срок автоматическим затвором. В этот момент движение воздуха прекращается и манометр регистрирует давление в трубке между ртом и затвором. Это давление равно альвеолярному, так как скорость потока равна нулю. Короткий срок, на который прерывается поток, не нарушает дыхания. Разность давлений, измеренных при неподвижном воздухе и в момент, непосредственно предшествующий прерыванию потока, представляет собой величину падения давления в воздухоносных путях. Разделив величину падения давления на величину объемной' скорости воздуха (которая непрерывно регистрируется пневмотахогра-фом) для данного момента времени, получают величину аэродинамического сопротивления.
Сопротивления, связанные с гравитацией и инерцией. При дыхании происходят изменения положения грудной клетки и органов брюшной полости в гравитационном поле Земли. При этом в зависимости от фазы дыхательного цикла и ориентации положения тела в пространстве сопротивление, связанное с тяжестью перемещаемых органов, меняется. Вес грудной клетки при вдохе оказывает сопротивление мышцам вдоха; при выдохе этот
326
фактор способствует уменьшению объема грудной клетки. Сила тяжести брюшных внутренностей (особенно в положении стоя) воздействует на диафрагму, стремясь оттянуть ее вниз. Это в некоторой степени способствует вдоху и оказывает сопротивление выдоху. Влияние гравитационных факторов на дыхание можно установить при сравнении механических и вентиляционных показателей дыхания человека в положениях лежа и стоя.
Поскольку при дыхании приходит в движение система, обладающая массой, то при этом проявляется ее инерция. Сопротивление, обусловленное инерцией, возникает при изменении скорости движения, т. е. при ускорении. Давление Р и, необходимое для преодоления инерции, определяется уравнением:
|
|

(9)
где т — масса перемещаемых органов; V — вторая производная от объема по времени.
Инерционный фактор в обычных условиях дыхания весьма мал и приобретает некоторое значение лишь при увеличении ускорений, что бывает при очень частом дыхании и увеличенной вентиляции.
РАБОТА ДЫХАНИЯ
Работой дыхания называется работа по преодолению всех видов сопротивления, которая выполняется дыха-тельными мышцами при вентиляции легких. Роль от-дельных мышц, ихактивность в разные фазы дыхательного цикла и в различных условиях дыхания исследуются с помощью метода электромиографии. Непосредственно измерить работу дыхательных мышц не представляется возможным.
В связи с этим для определения работы дыхания пользуются косвенными методами.
Работа, как известно, измеряется произведением силы на путь, пройденный по направлению силы. Для случая дыхательной (трехмерной) системы работа выражается произведением давления на изменение объема. Поскольку давление в дыхательной системе величина переменная, определение работы А производят путем интегрирования:
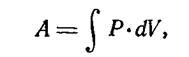
327
где Р — общее давление, приложенное к дыхательной системе в каждый момент дыхательного цикла; йУ — приращение объема системы.
Полная работа дыхания определяется методом об
щей плетизмографии. Испытуемого с выключенным са-
мопроизвольно или с помощью фармакологических
средств дыханием помещают в плетизмограф, в котором ритмически создается отрицательное и положительное давление. Для определения работы необходима одновременная запись спирограммы, показывающей легочные объемы, и разности давлений между плетизмографом и ротовой полостью. По полученным данным строят кривую зависимости давления от объема (см. рис. 63). Работа дыхания численно равняется площади фигуры, Образуемой замкнутой линией, соединяющей последовательные значения давлений и объемов в каждый момент дыхательного цикла.
Полученная фигура дает представление о работе по преодолению как общего сопротивления дыханию, так и его отдельных компонентов. Работа во время вдоха равняется площади АЕВС и состоит из двух частей: АВС — работы по преодолению статического сопротивления дыхательной системы; АЕВ — работы по преодолению динамического сопротивления дыханию. Во время выдоха совершается работа АВСД. Но так как АВС представляет собой потенциальную энергию, накопленную во время вдоха, то общая работа дыхания равна площади АЕВСД. При этом площадь АСД показывает работу по преодолению динамического сопротивления выдоху.
Определение общей работы дыхания по вышеописанной методике практически весьма сложно. Поэтому более широко изучается та часть работы дыхания, которая затрачивается на преодоления сопротивления легких и дыхательных путей (без учета сопротивления грудной
клетки). Для этого достаточно регистрировать динамику плеврального (пищеволного) давления во время естественного дыхания при одновременной записи дыхательных объемов. Последующее определение работы принципиально не отличается отописанного выше.
Величину дыхательной работы относят к 1 л вентиляции. В покое (вентиляция до 10 л/мин) работа
| 328 |
дыхания, по данным разных авторов, составляет величины 0,1—0,59 Дж/л, или 0,98—4,9 Дж/мии. По мере уве-
личения минутного объема работа дыхания возрастает не пропорционально, что связано главным образом с увеличением динамического сопротивления. Работа дыхания бывает повышенной при увеличенном эластическом или динамическом сопротивлении и особенно при сочетании обоих этих факторов. Предельная для каждого человека величина вентиляции лимитируется доступной для него предельной работой дыхательной мускулатуры. С резко увеличенной работой дыхания обычно связано тягостное ощущение одышки.
Показано, что обычные сочетания глубины и частоты дыхания близки к оптимальным, т. е. таким, которые обусловливают минимальную работу на 1 л вентиляции. Это обеспечивается функциями систем регулирования дыхания.
|
|

|
|

|
|

|
|

|
|

|
|


