




Заболевания
Репродуктивной системы
 Викки Мейерс-Валлен
Викки Мейерс-Валлен
РЕДАКТОР-КОНСУЛЬТАНТ
Врожденные заболевания репродуктивной системы у собак и кошек................................................ 994
ДНК-тестирование наследственных заболеваний собак........................................................................ 999
Использование прогестерона сыворотки крови для регуляции времени овуляции у сук........... 1004
Искусственное осеменение сук.................................................................................................................... 1006
Диагностика беременности у сук................................................................................................................ 1008
Использование релаксина сыворотки крови для диагностики беременности у сук..................... 1013
Бесплодие у сук................................................................................................................................................. 1015
Бесплодие у кошек........................................................................................................................................... 1019
Современные терапевтические рекомендации в отношении беременных сук.............................. 1021
Медикаментозное лечение патологических родов и показания для кесарева сечения у сук...1023
Влияние гипотиреоза на бесплодие кобелей............................................................................................ 1029
Заболевания семенников у кобелей............................................................................................................ 1031
Обзор методов прерывания беременности у сук..................................................................................... 1036
Врожденные заболевания репродуктивной системы
У собак и кошек
Викки Мейерс - Валлен
НОРМАЛЬНОЕ ПОЛОВОЕ РАЗВИТИЕ
Нормальное половое развитие зависит от успешного завершения трех последовательных этапов: (1) формирования половых хромосом, (2) развития половых признаков на гонадном уровне и (3) развития половых признаков на фенотипическом уровне. Набор половых хромосом, который соответствует генетического полу здоровых животных, устанавливается при оплодотворении. Зигота получает или две Х-хромосомы, или одну X, а другую Y-хромосому, и этот хромосомный набор поддерживается во всех клетках путем митоза. На ранних стадиях морфология XX- и XY-эмбрионов одинакова. У обоих есть генитальный гребень, из которого в дальнейшем развиваются семенники или яичники. У них также есть мюллеровы и вольфовы протоки, урогенитальный синус, генитальный бугорок и генитальные выпуклости, из которых развиваются внешние и внутренние половые органы (рис. 1). Дифференциация генитального гребня в семенники или яичники определяет пол на гонадном уровне и является окончанием индифферентной стадии развития эмбриона. Хотя для нормального развития индифферентной стадии необходимо несколько генов, но в половом развитии гены, определяющие пол, играют центральную роль.
В нормальном состоянии половые признаки на гонад ном уровне определяются по набору половых хромосом: присутствие Y-хромосомы приводит к развитию семенников, а ее отсутствие - к развитию яичников. Опредв ющий пол региональный Y-ген, Sry, расположен] Y-хромосоме. В этом гене закодирован фактор, опред ющий развитие семенников, и он является генетиче сигналом для начала дифференциации семенников генитального гребня (обзор Goodfellow and Lovell-b 1993). Sry полностью отличается от гена для H-Y-a на, которого больше не считают способным ишш вать развитие семенников. Трансгенетические экс менты на мышах показали, что Sry единственный Y-ген необходимый для индукции семенников (Коортап et al. 1991), есть и другие доказательства этой теории (о Goodfellow and Lovell-Badge, 1993). При отсутствии Y-хромосомы и Sry генитальный гребень в норм состоянии становится яичником. Ген Daxl обычно располагается на каждой Х-хромосоме в двойном количестве может играть роль в подавлении мужских генов во время дифференциации яичников {Swain et al., 1996).
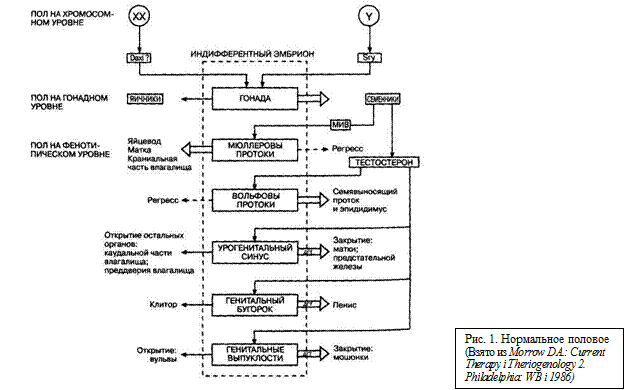 Фенотипические половые признаки в нормальном с стоянии контролируются гонадными половыми признаками. Если перед гонадной дифференциацией уд генитальный гребень из XX- и XY-эмбрионов, то будет развиваться женский фенотип. Это показывает, что эмбрион запрограммирован как на развитие женского пола, так и его развитие может быть переориентировано на развитие мужской особи. Критической стадией переорентации является развитие семенников. Семенники выделяют два вещества, которые действуют весь критический период эмбрионального развития и побуждают эмбрион развиваться в мужскую особь: (1) мюллерово ингибирующее вещество (МИВ), которое вызывает регресс мюллеровых протоков, и (2) тестостерон, который стимулирует формирование семявыносящих сосудов и эпидидимуса из вольфовых протоков (рис. 1). В наружных половых органах тестостерон (Т) конвертируется в дигидротестостерон (ДГТ) ферментом 5-альфа-редук-тазой. Дигидротестостерон стимулирует образование предстательной железы, мужской уретры, пениса и мошонки соответственно из урогенитального сину-са, генитального бугорка и генитальных выпуклостей (рис. 1). Опускание семенников в мошонку завершает образование наружных мужских половых органов, но генетический и гормональный контроль всех этих процессов еще до конца не выяснен. При отсутствии семенников и их мужских гормонов развиваются женские половые органы (рис. 1).
Фенотипические половые признаки в нормальном с стоянии контролируются гонадными половыми признаками. Если перед гонадной дифференциацией уд генитальный гребень из XX- и XY-эмбрионов, то будет развиваться женский фенотип. Это показывает, что эмбрион запрограммирован как на развитие женского пола, так и его развитие может быть переориентировано на развитие мужской особи. Критической стадией переорентации является развитие семенников. Семенники выделяют два вещества, которые действуют весь критический период эмбрионального развития и побуждают эмбрион развиваться в мужскую особь: (1) мюллерово ингибирующее вещество (МИВ), которое вызывает регресс мюллеровых протоков, и (2) тестостерон, который стимулирует формирование семявыносящих сосудов и эпидидимуса из вольфовых протоков (рис. 1). В наружных половых органах тестостерон (Т) конвертируется в дигидротестостерон (ДГТ) ферментом 5-альфа-редук-тазой. Дигидротестостерон стимулирует образование предстательной железы, мужской уретры, пениса и мошонки соответственно из урогенитального сину-са, генитального бугорка и генитальных выпуклостей (рис. 1). Опускание семенников в мошонку завершает образование наружных мужских половых органов, но генетический и гормональный контроль всех этих процессов еще до конца не выяснен. При отсутствии семенников и их мужских гормонов развиваются женские половые органы (рис. 1).

