




В основі різноманітності типів археологічних культур пізнього плейстоцену і раннього голоцену, а також їх подальшої спеціалізації лежали екологічні зміни, але основним стимулом були культурні контакти, дифузія і пересування народів [42]. Спільні риси багатьох культурних традицій свідчать про те, що максимальна пристосованість до середовища забезпечувалась ідентичними методами. Незначні варіанти культурного розвитку виникали внаслідок специфічності умов існування різних колективів. Давні культури починають швидко змінювати одна одну. На базі Сангойської культури формуються інші. Атерійская культура (мустье на ножці) існувала на півночі Сахари та в регионі гір Атласу 90-20 тис. р. т. Це одна з найперших культур, в якій почали використовувати лук та стріли. В мезоліті її змінює іберо-мавританська культура, зв’язок між атером та іберо-мавританською культурою залишається дискусійним. В Африці також існували культури Стілбей (71тис. 900 р. т. – 71 тис. р. т.) та Ховьєсонс Пурт (64 тис. 800 р. т – 59 тис. 500 р. т.), які відрізнялися особливо яскравими зразками мистецтва (прикраси з раковин, геометричні візерунки на камінні та шкаралупі страусів). Першою знахідкою людей в Азії стали знахідки людей в Омані (Джебель-Файя). Саме вони є скоріш за все безпосередніми предками Азійських людей, що висадились вперше на територію Аравії (127- 95 тис. р. т) (рис. 73) [42].

Рис. 73. Знаряддя праці людей із Джебель-Файя: 1 – подвійно загострене листовидне знаряддя; 2 – Левалуазський відщеп; 3 – заготовка для подвійно загостреного рубила (біфасу)[42].
На території Індії люди зявилися вже 80-74 тис. р. т., ще до виверження сумнозвісного вулкана Тоба на о. Суматра (різке похолодання, що скоротило чисельність людської популяції) [42, 128]. Емірійська культура (36-47 тис. р. т.) вже чітко притаманна для Близького Сходу. Виросла на базі місцевого мустьє, під впливом вихідців із Африки, еволюціонувала в ахмаріан (місцева культура верхнього палеоліту). Барадостська культура (36 тис.р. т. Була притаманною для Ірану та Іраку. Костенківсько-стрілецька культура (32-30 тис. р. тому) притаманна для Північного Причорномор’я. Еволюціонувала на межі мустьє та прийшлих азійських культур, стала родоначальником великої групи верхньопалеолітичних культур. Для Ц. Азії та Сибіру у відповідності з Мочановим Ю. А. характерні дві основні групи культур, що можливо мають відношення до предків частини населення Європи (Мальтинсько-буретська, Афонтовська, Кокорівська, Сроскінська, Ошурковська, Самномиська к-ри, віком 24000-14750 р. т) та Америки (Дюктайська, Сумнагітька к-ри, 35000-10500 р. т.). Верхній палеоліт – культур вже дуже багато, найбільш важливими в Європі є:
· Шательперон – територія Франції 35-29 тис р. т. Інтерстадіал Вюрмської епохи I\II, переходна культура між місцевим мустьє і солютре (вихідці з Африки та Азії). Можливо це була частина неандертальців, що намагалася адаптуватись до прийшлих неоантропів.
· Селет (40-28 тис р. т.) – Угорщина, Чехословачина, півд-зах. Польщі. Походить від місцевого варіанта мустьє, змінилась граветтськими культурами.
· Оріньяк – частина країн Західної та Центральної Європи. Повністю культура неоантропів (рис. 74) [144]. Співіснує з перигорською, змінюється солютрейскою культурою.
· Граветтськая культура – 28-21 тис. р. т. Третій етап в оріньяко-перигорському комплексі культур. Існував перед мадленом.
· Солютре (18-15 тис. р. т.) – змінює оріньяк та перигор на території Франції та Північної Іспанії. Можливо, солютрейці проникли в Північну Америку та прийняли участь в створенні культури кловіс (рис. 75) [144] (проіснувала дуже недовго), тощо.
·  Мадлен (15-8 тис.р. т.) – змінює попередні культури та поширюється від Зах. Європи тдо Уралу.
Мадлен (15-8 тис.р. т.) – змінює попередні культури та поширюється від Зах. Європи тдо Уралу.
Рис. 74. Оріньякська Мадонна[42].

Рис. 75. Знаряддя культури Кловіс – гостряки (Півн. Америка) [42].
Особливості трудової діяльності кроманьйонців [20, 42, 46, 60, 130]:
· виготовлення спеціальних засобів праці, які були представлені різцями, відбійниками і ретушерами, котрі змінили весь процес праці (рис. 74, 76, 77, 78) [42, 144];
· виготовлення прикрас з каменю, мушль, кісток, шкаралупи страусів (саме прикраси з мушль та шкаралупи страусів, гравіювання на каменях та шкаралупі є одним із найважливіших ознак неоантропів, що свідчать про значну зміну свідомості);
· оволодіння прийомами роботи з посередником (засобом праці), котрий починає займати основне місце в обробці каменя і поступово замінює безпосереднє оббивання нуклеусу, яке також суттєво змінюється. Якщо палеоантропи оббивали камінь, утримуючи його в руках, то неоантропам для виконання цієї ж процедури вже потребувався весь нервово-м'язовий аппарат організму: сильний короткий поштовх корпусом, спрямований на рукоятку віджимника (посередник праці, засіб праці), повинен був поєднуватися з дуже точним і дуже близьким до краю положенням наконечника віджимника. Пластинка відщеплялася з першого разу лише у виняткових випадках. Пізньопалеолітична людина повинна була виконувати багаторазові спроби, виявляючи завзятість у досягненні мети і чималу майстерність. Вона базувалася вже не тільки на обізнаності з властивостями матеріалу, але і на правильному розрахунку рухів, скоординованості дій тулубу, рук і ніг. Функціональна складність подібних прийомів дуже висока;
· необхідність великої підготовчої роботи з нуклеусом з одночасним спрощенням самого процесу одержання знарядь, який складається практично з одного вдалого натиснення або удару по посереднику. Пластинка-відщеп сама по собі вже становила готове дволезове знаряддя праці. Вторинна обробка набувала протилежного значення: ретуш повинна була не загострити, а затупити лезо, щоб край пластинки не різав руку під час роботи;
· надзвичайна економія матеріалу, коли використовувався майже кожен відщеп-пластинка і навіть залишки нуклеусу застосовували як відбійник під час ретушування;
· подальший розвиток техніки ретуші, з'являються нові її різновиди: ударна, контрударна, точкова, віджимна;
· зміна техніки роботи з кісткою, засвоюються нові прийоми її обробки: пиляння, свердління, шліфування. Це дозволяло виготовляти голки, шила, кістяні наконечники стріл, які змочували отрутою, тощо;
· виникнення нових прийомів роботи зі шкірами: шиття, лощення для забезпечення їх непроникності для води тощо;
· спроби виправити зіпсовані знаряддя і повернути їм робочі властивості.
Таким чином, техніка пізнього палеоліту була дуже раціональною, а процес виготовлення знарядь значно удосконалився за рахунок їх мінімізації і появи виробів, які одержали назву мікролітів. Мікроліти – невеличкі пластинки або відрізки подібних до ножів пластин і відщепів, які часто мали визначену геометричну форму. З одного кінця за допомогою ретуші вони затуплювалися, а з другого – формувалася гостра робоча частина. Затуплені кінці дозволяли прикріплювати держак.

Рис. 76. Знаряддя Орінякської епохи Європи [144].

Рис. 77. Знаряддя епох протосолютре та солютре Європи [144].
З декількох мікролітів, закріплених за допомогою смоли, виготовляли складні види знарядь. Природні рослинні смоли використовували в розплавленому вигляді. Після затвердіння вони давали жорстке і міцне з'єднання. Мікроліти використовували як зубці для гарпуноподібних дротиків, наконечники стріл, складові частини для ножів, серпів, пилок тощо. Таким чином, у неоантропів з'являються перші складні знаряддя і здійснюється перехід до синтетичного уявлення про технологічну послідовність операцій. Виготовлення таких принципово нових знарядь потребувало і застосування нового матеріалу. З цього приводу кроманьйонці першими звернули увагу на такі дрібнозернисті породи, як халцедон, кристалічний кварц, агат, кременистий вапняк тощо.
Суттєве зростання ефективності трудової діяльності неоантропів було б неможливим без відповідного розвитку нервово-психічних процесів, який у людей пізнього палеоліту здійснювався в напрямку переважання процесів, пов'язаних із оволодінням уявленнями про складні взаємодії між предметами і явищами в часі. Мета (одержання якогось знаряддя) віддаляється від початку роботи на довший час, ніж у мустьєрців. Причини: виготовлення знарядь праці і сам процес труда, з погляду психологічних механізмів, стають усе більш опосередкованими і абстрактними, оскільки тепер для виготовлення знаряддя людини повинна мати уявну мету, задум не тільки щодо цього знаряддя, але і відносно посередника – засоба праці. Кінцева абстрактна мета досягається вже не тільки за допомогою безпосереднього порівняння задуму з тим конкретним предметом, що виходить, а більш складним шляхом, коли до цього ланцюга нервових механізмів додається ще одна ланка, котра також базується на абстрактному задумі (одержання засобу праці) і його практичному виконанні (конкретна форма засобу праці). Така складна форма інтеграції пов'язана, головним чином, із функціонуванням префронтальної ділянки мозку за участю заднього, тім'яно-висково-потичного відділів кори. Вона є результатом третинного аналізу сигналів, які вже пройшли неодноразову переробку в первинних і вторинних зонах. Задум, який формується на основі аналітико-синтетичної діяльності кори задніх відділів півкуль, передається в лобову зону, де здійснюється його перешифрування в серію організованих у часі та просторі рухових актів. Нервово-психічні процесів ускладнювались паралельно із засвоєнням нової техніки виготовлення знарядь праці. Високого ступеню досягла координація всіх рухів, особливо тих, що зумовлені функцією кори. Техніка роботи з посередником потребувала значної координованості дій усього організму. Віднайдені численні сліди невдалих багаторазових спроб розщеплення каменю. Вони свідчать про те, що людина пізнього палеоліту досить важко засвоювала руховий комплекс, необхідний для одержання відповідних знарядь. Для цього потребувалися узгоджені рухи, зумовлені діяльністю кірку гловного мозку, й пов'язані з волею, настирливістю, раціональним підходом до виготовлення знарядь тощо. Забезпечити подібні властивості здатна була лише більш висока самоорганізація нервових процесів, зумовлена сумісною еволюцією передньолобових відділів, тім'яної частки і мозочку.
Загальні особливості сумісної еволюції мозкових структур, нервових процесів і трудової діяльності гомінід протягом антропогенезу для зручності сприйняття подані в Додатку А.
Відбувався паралельний розвиток нервової системи, психічного відображення, трудової діяльності й суспільної організації, що став основою для принципово нового напрямку еволюції живого світу: суто біологічні фактори ставали все менш важливими для виживання людей, а функцію регуляції життєздатності особин приймав на себе соціум. Змінювалися всі форми взаємовідносин нового виду (людина розумна) з довкіллям. Так, неоантропи першими застосували знаряддя не тільки як інвентар, але й як зброю. Лук і стріли з'явилися приблизно 90 тис. років тому в палеоліті (давній кам'яний вік) і вже 20 тис. р. т. швидко поширилися по всій Ойкумені. Завдяки цим революційним винаходам неоантропи змогли полювати і на прудконогих звірів, оскільки за допомогою лука вони могли влучити в ціль навіть за 100 м і зробити близько 20 пострілів за хвилину. Почали складатися етнічні й расові особливості, формувалася родова організація суспільства. Виникло мистецтво у вигляді печерних малюнків, гравюр тварин і людей, глиняних і кам'яних скульптур і рельєфів. Перші представники нашого виду характеризуються саме активним використанням прикрас, що активно виготовлялися з мушель та шкаралупи стравусиних яєць. Можливо в їх свідомості прикраси також були своєрідними оберігами, виконували певну магічну захисну функцію (для створення ефекту «плацебо», самозаспокоєння). Вони досить часто зустрічаються на берегах океанів, що чітко ідентифікує шляхи міграції перших людей. З’являються чисельні картини з зображеннями звірів, на яких людина полює. Так, відомі «печерні галереї» у Франції, Іспанії, Африці, на Уралі й в інших регіонах Землі. Деякі малюнки зображують постаті напівлюдей – напівтварин. Припускають, що така штучно створена віртуальна реальність мала магічне значення і використовувалася чаклунами і шаманами під час певних ритуалів. Не виключається, проте, і можливість того, що це просто результат фантазії первісного художника. Одночасно подібні фігурки несли і певне психологічне навантаження, призводячи до приблизно такого висновку: якщо те, чого немає насправді, зображується, то можливо такі істоти живуть в якомусь іншому світі. Велике значення в мистецтві мав образ жінки, що, швидше, асоційовався з матріархатом. Важливий матеріал, який висвітлює духовне життя людей того часу, дають поховання і пов'язані з ними ритуали. На стоянках верхнього палеоліту знайдені могили у яких знаходилися різні предмети, дорогі прикраси тощо. Отже, у кроманьйонців ідеологічні уявлення вже суттєво ускладнилися, що і відбилося в появі більш різноманітних обрядів.
Людина сучасного типу, як було зазначено, покинула межі Африки відносно недавно – близько 127- 95 тис. р. тому [42]. Можливо весь цей час вона не могла вийти за межі Африки через активний спротив конкурента – неандертальця. Проте, після того як людина вийшла за межі Африки, відбулося мінімум 1-2 гібридизації з близькосхідними неандертальцями, про що свідчать дані палеогенетики (1-4% специфічних для неандертальця генетичних маркерів у людей неафриканських популяцій). Крім того на даний час існують знахідки віком від 130 до 90 тис. р. т. на Близькому Сході, що можуть бути свідотством гібридизації неспецифічних неандертальців та людей сучасного типу, що прийшли на Бл. Схід по коридору р. Ніл з Африки (рис. 78) [130]. Людина спочатку заселяє Аравійський півострів, куди потрапляє через Аденську затоку, потім Індію, а звідти – Індокитай. Важливим для неї було приручення свого азійського ворога – вовка, котрого вона приручила, ще до того, як потрапила в Австралію (собака Динго є вторинно здичавілим видом, про що свідчать дані взаємовідносин їх з популяцією аборигенів – симбіоз). Переважна більшість геному домашньої собаки – це геном Індійсько-Індокитайської популяції вовків [20]. Можливо саме собака сприяла певною мірою становленню людини та сприяла винищенню нею популяції останніх неандертальців Європи (як розносник інфекційних хвороб, до яких розумна людина встигла адаптуватись, або як засіб полювання на своїх ворогів, та нічного захисту від нападів неандертальцю). Европейський неандерталець ще 3-4 тис. р. співіснував з сучасною людиною в Європі, поки не був знищений близько 35 тис. р. тому. До речі, рештки людей в вогнищах неандертальців мають пошкодження, які свідчать про те, що люди були вбиті підступно, переважно в спину, можливо під час сну. А решки неандертальців свідчать про те, що їх вбили під час прямого нападу. Тобто між неандертальцями та людьми сучасного виду відбувалися активні зіткнення [42].



Рис. 78. Черепи давніх людей: верхній ряд – людина розумна (ліворуч), неандерталець (праворуч) (www.nkj.ru/news/24252/), нижній ряд – гібридні люди з печер Джебель Кафзех 11 та Схул V в Палестині[130].
Практикувався конкурентний канібалізм. Можливо розповсюдженню людей також сприяло те, що близько 72 тис. р. тому відбулося виверження супервулкану Тоба на о. Суматра, після якого відбулося швидке та сильне похолодання в межах Ойкумени, що призвело до скорочення популяції ворога, який жив набагато північніше, ніж люди нашого виду. До того ж є свідчення того, що генетичне різноманіття неандертальцю значно знизилось (відомі всього три жіночі лінії мітохондріальної ДНК неанденртальцю на просторі від Європи до Алтаю, у людей нашого виду – понад сорок).
Існують свідчення того, що люди нашого виду гібридизували також і з іншими пізніми антропоїдами, такими як Денисівська людина, зразки ДНК якої були вилучені із кісткових знахідок в Денисовій печері на Алтаї, і можливо останніми представниками людини випрямленої зі Східної Азії [42]. Це другий вид вимерлих представників гомінін, для якого став відомий повний мітохондріальний і майже повний ядерний геноми, які й дозволили пролити світло на приналежність цих решток. Вид жив близько 40 тисяч років тому і мав ареал, що перетинається за часом і місцем з територіями, що населялись неандертальцями і сучасними людьми. Команда вчених з Лейпцигського Інституту еволюційної антропології товариства Макса Планка під керівництвом шведського біолога Сванте Пяябо (Svante Pääbo) секвенували ДНК, вилучену з фрагмента кістки фаланги дитячого пальця, знайденого в 2008 р. російськими археологами в Денисовій печері на Алтаї [42]. З'ясувалося, що мітохондріальна ДНК цього зразка відрізняється від мт-ДНК сучасної людини по 385 нуклеотидах, у той час як мітохондріальна ДНК неандертальців відрізняється від ДНК Homo sapiens по 202 нуклеотидах. Статтю, присвячену цьому відкриттю, було опубліковано в журналі Nature 24 березня 2010 р. Пізніше, коли були оброблені послідовності, пов'язані з ядерним геномом, виявилося, що Денисівська людина все ж ближче до неандертальця, та їх еволюційне розходження відбулося близько 640 тис. років тому. Можливо сам Денисівець є гібридом між гейдельбержцями та азійськими еректусами.
Вік знахідок, виявлених в печері в тих же самих шарах, був визначений за допомогою радіовуглецевого аналізу в 40 тисяч років.
Геном Денисівської людини, прочитаний «начорно» в 2010 році, вдалося відсеквенувати «набіло» (з 30-кратним покриттям) завдяки новим методам роботи з древньою ДНК. Аналіз Денисівського генома підтвердив, що сапієнси після свого виходу з Африки схрещувалися з архаїчним євразійським населенням – неандертальцями і денисівцями. Дослідження також показало, що денисівці були нечисленною популяцією з украй низьким рівнем генетичної різноманітності. Найважливішим результатом є складання детального каталогу генетичних змін, що закріпилися у сапієнсів після їх відділення від спільних з денисівцями предків. Деякі з цих змін торкнулися генів, що впливають на роботу нейронів і розвиток мозку (ген мікроцефалін). Це означає, що між психікою сапієнсів і денисівців могли бути генетично зумовлені розходження. Знайдено сліди гібридизації Денисовської людини з Homo sapiens. У геномі меланезійців, тибетців, австралійців виявлено близько 5% спільних генів з прочитаним ядерним геномом Денисівської людини. У той же час спільних генів у сучасної людини і неандертальця від 1% до 4% у різних популяціях (за винятком Африки, де неандертальські гени практично відсутні) [42].
Цікавою є також знахідка «людей з печери червоного оленя» в Китаї, які є можливо гібридами між людьми розумними з якимись місцевими гомінідами, такими як еректуси, чи може й власне Денисівцями. Людина Печери червоного оленя – це наймолодші рештки доісторичних людей не виду Homo sapiens, відомі на 2012 рік. Згідно з радіовуглецевим аналізом вони датуються 14500 – 11500 р-р. т. Вивчені люди мали мішанину архаїчних і сучасних ознак. У них було округле склепіння черепа з товстими кістками, виступаючі надбрівні дуги, досить коротке і пласке обличчя, широкий ніс. Нижні щелепи цих людей виступали вперед, але у них не було такого підборіддя, як у сучасних людей. Також у людей цього виду були дуже великі корінні зуби. Мозок у них середнього розміру (лобові долі схожі за будовою на долі сучасної людини, а от тім'яні частки погано розвинені). Вчені вважають, що ці люди були вищими за неандертальців у своєму розвитку, так, томограма черепа показала, що фронтальні частки мозку мали бути схожі з тими, що маємо ми сьогодні. З причини того, що зоною їх проживання був регіон, що відрізнявся сонячною активністю, швидше за все, вони мали темну шкіру. Як відзначають вчені, даний вид однозначно пов'язаний з Homo sapiens, але є принципово новою гілкою його еволюції.
Аналіз черепів показав: ці гомініди мали унікальну суміш стародавніх і сучасних анатомічних рис, констатує Даррен Карно з Університету Нового Південного Уельсу (рис.79) [42, 130].


Рис. 79. Реконструкція зовнішності людини з печери червоного оленя та її череп.
[130].
Певні риси цього типу людей можливо помітити у корінного населення Австралії та Меланейзії. Саме вони є нащадками першої хвилі сапієнсів, що прийшли першими в Індокитай (носії чоловічої у-хромосомної гаплогрупи – С, що є альтернативною до основної для сучасного людства – гаплогруппи F та її більш пізніх варіантів, див. вище [124]). Цікавою є картина первинного розселення людей нашого виду по світу. Прослідкувати цей процес за соматичними генами – майже неможливо, тому що ділянки батьківської та материнської ДНК рекомбінуються внаслідок кроссінговеру та танку хромосом в мейозі. Проте в геномі людини є частини ДНК, що не рекомбінує в процесі розмноження. Це так звана мітохондріальна ДНК, що передається в потомстві тільки по жіночій лінії, через мітохондрії яйцеклітини (сперматозоїди не містять функціональних мітохондрій) та у-хромосомна ДНК(NRY-маркер), що передається тільки по чоловічій лінії (у-хромосома коньюгує з х-хромосомою лише частково, на проксимальному кінці). Тільки з часом в цих лінях відбувається накопичення мутаційних змін, швидкість та час яких можна прослідкувати за методом молекулярного годинника. Внаслідок мутацій утворюються нові варіанти ДНК, які називають гаплотипами, що за своєю спорідненістю об’єднуються в гаплогрупи [124, 125, 160, 181].
Гаплогрупа (з грец: ἁπλούς, haploûs – однократні, одиночні, прості) – група схожих гаплотипів, які поділяють також загальний поліморфізм одного нуклеотиду.
Цей термін широко застосовують в генетичній генеалогії (популяційній генетиці), де вивчають Y-хромосомні гаплогрупи (Y-ДНК), мітохондріальні ДНК (мтДНК) і ГКГ-гаплогрупи. Гаплогрупи є спільні для певної групи людей – нащадків одного предка (рис. 80) [168].

Рис. 80. Принцип успадкування гаплогруп мт-ДНК (за жіночою лінією) та у-ДНК (за чоловічою лінією) [168].
З цієї схеми добре зрозуміло, що якщо у матері немає дочки, то жіноча гаплогрупа далі не успадковується, якщо у батька немає сина, то його у-хромосомна гаплогрупа теж не успадковується. Всі нинішні люди мають загального предка по жіночій лінії – «мітохондріальна Єва», та по чоловічій лінії – «у-хромосомний Адам». Проте це не значить, що всі люди є лише їх нащадками, тому що існує ще соматична ДНК з нестатевих хромосом. Просто ці Єва та Адам залишили потомство саме за своїми чоловічими та жіночими статевими лініями, т. я. була певна перевагу їх потомству саме за цими ознаками. Решта – передали свої соматичні гени, які прослідкувати набагато важче, через кросинговер та танок хромосом (мейоз).
В 2002 році спільнота генетиків (Y Chromosome Consortium) виробила загальну класифікацію та номенклатуру за ліниями Y-хромосоми (STR). Виділено 21 основний мутаційний кластер в хромосомі, так звані клади, що позначаються латинськими буквами від A (Аоо) до Т (рис.81) [124, 160, 181]. Порядок букв відбиває послідовність виникнення мутацій. Ці клади в свою чергу гілкуються на гаплогруппи, які нумеруються цифрами та буквами.

Рис. 81. Загальна схема чоловічих ліній сучасних людей за у-хромосомною ДНК (У-гаплогрупи) [124].
Зокрема в світі вони розподіляються таким чином (рис. 82) [48, 67, 71, 72, 79, 86, 101, 103, 105, 108, 111, 112, 113, 124, 129, 160, 161, 181]:
Найдавніші групи без мутації М168:
Гаплогрупа А (М91) –Африка, зокрема койсанці, ефіопи, нілоти.
Гаплогрупа В (М60) – Африка, зокрема піґмеї і хадзапе.
Групи з мутацією М168 (сталася бл. 70-50 тис. до нашої ери):
Гаплогрупа C (М130) – Океанія (С5), Північна, Центральна, Східна Азія, Північна Америка (С3); Австралія (С4), трапляється в Індії (С2), дужє рідко в Європі (С1).
Гаплогрупи YAP+:
Гаплогрупа DE (М1, М145, М203):
Гаплогрупа D(М174) – Тібет, Японія, Андаманські острови.
Гаплогрупа E (М96)
Гаплогрупа E1b1a (V38) – Зах. Африка, відома раніше як Е3а, переважно у банту.
Гаплогрупа E1b1b (М215) – Східна та Північна Африка, Близький Схід, Середземномор'я, Балкани, великий відсоток у греків (раніше Е3b). Поширена в Україні (до 4%).
Групи з мутацією М89 (сталася бл. 45 тис. до нашої ери):
Гаплогрупа F (P14, M213) – Південна Індія, Шрі-Ланка, Ц. Азія, Кавказ, Китай, Корея.
Гаплогрупа G (M201) – наявна в багатьох етнічних групах Євразії, звичайно у невеликій пропорції: Кавказ, Іран, Афганістан, Кашмір, Анатолія, Греція, Італія, Тіроль, Чехія; дуже рідко у Північній Європі. На Україні до 4%.
Гаплогрупа G2 – основна підгрупа групи G в Європі
Гаплогрупа H (М69) – Індія, Шрі-Ланка, Непал; рідко в Пакистані, Ірані, Центральна Азія, Аравія. В Європі – серед циган.
Групи з мутаціями L15, L16 (сталися бл. 50-42 тис. до нашої ери):
Гаплогрупа IJK (L15, L16):
Гаплогрупа IJ (S2, S22):
Гаплогрупа I (М170, P19, M258) — поширена в Європі, зрідка на Близькому Сході:
Гаплогрупа I1 (М253, М307, Р30, Р40) – Північна Європа. У потомків скандинавів. В Україні – до 2-4%.
Гаплогрупа I2 (S31) – Центральна і Південно-Східна Європа, Сардинія. Поширена на Балканах. На Україні – до 18%.
Гаплогрупа J (М304) – Близький Схід, Туреччина, Кавказ, Італія, Греція, Балкани, Східна Європа, Північна і Північно-Східна Африка. В Україні складає 6%.
Гаплогрупа J* – о. Сокотра, зрідка у Пакистані, Омані, Греції, Чехії, зустрічається серед тюркських народів.
Гаплогрупа J1 (М267) – переважно семітські народи, вірмени, також Близький Схід, Ефіопія, Північна Африка, Дагестан.
Гаплогрупа J2 (М172) – Західна, Центральна, Південна Азія, Південна Європа, Північна Африка, острови Середземного моря.
Групи з мутацією М9 (сталася бл. 40 тис. до нашої ери):
Гаплогрупа K:
Гаплогрупа LT (L298/P326):
Гаплогрупа L (М11, M20, M22, M61, M185, M295) – Південна, Південно-Західна, Центральна Азія, Середземномор'я.
Гаплогрупа T (М70, M184/USP9Y+3178, M193, M272) – Північна Африка, Африканський Ріг, Південна і Південно-західна Азія, Середземномор'я, раніше відома як К2. В Україні – до 2%.
Групи з мутацією М526:
Гаплогрупа М (Р256) – Нова Ґвінея, Меланезія, Східна Індонезія.
Гаплогрупа NO(М214).
Гаплогрупа N (М231) – крайня північ Євразії, особливо серед уральських народів, тюрки (Якути), Японія, Корея (Фіни. Гуни). В Україні – до 6%
Гаплогрупа О (М175) – Центральна, Східна, Південно-Східна, Південна Азія, Тихий Океан.
Гаплогрупа P (М45, 92R7, M74/N12) (мутація М45 сталася бл. 35 тис. р. до нашої ери).
Гаплогрупа Q(МEH2, M242, P36) – ендемічна в Америці, серед кетів, селькупів Сибіру, челканців Алтаю; появу у Європі пов'язують з гунами; мутації сталися у 20-15 тис. до нашої ери
Гаплогрупа R (M207, P36).
Гаплогрупа R1a (M17) – Центральна, Південна Азія, Центральна, Північна, Східна Європа, Ц. Азія (росіяни, українці, поляки, білоруси, словаки, словенці, моравські чехи, лужичі, татари, киргизи, каракалпаки, хакаси, шорці, телеути, теленгіти, алтай-кижі, уйгури, півд. башкири, тубалари, скандинави та ісландці, пакистанці та афганці, хунджанські та пенджикентські тадики, ішкашимці Паміру, сх. Перси, представники вищих каст Індії, ойрати, узбеки, хотони Ц. Азії, талиші Кавказу). В Україні – 54%.
Гаплогрупа R1b (M343) – Європа, Кавказ, Центральна, Південна Азія, Центральна, Північна Африка (англійці, шотландці, баски, зах. німці, голландці, французи, півн. італійці, албанці, вірмени, ассірійці, євреї-сефарди, грузини, туркмени, турки, кумандини, кумики, півн. башкири, казахи-кипчаки, уйгури, частина населення Камеруну). В Україні – 2%.

Рис. 82. Карта розподілу чоловічих гаплогруп до епохи великого переселення народів[124].
Гаплогрупа R2(М124) – Сх. Індія, Кашмір, Індокитай, Пакістан, Афганістан, Бурятія, Півн. Кавказ, у циган.
Гаплогрупа S (M230, P202, P204) -Нова Ґвінея, Меланезія, Східна Індонезія.
Подібно розподіляється по поверхні Землі і носії мт-ДНК (NRI). Проте вони відрізняються більшою стабільністю в плані переселення і є більш регіональними та осілими. Це можливо пов’язано зі специфікою поведінки жіночої половини людства, їх любов’ю до стабільності та зручностей (рис 83) [152].

Рис. 83.Схема первинного розселення жіночих ліній людей [152].
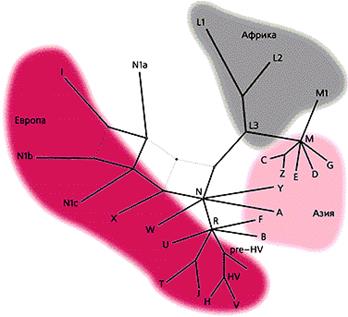
Рис. 84. Розподіл мт-ДНК в світі[129].
В Африці є близько 7 мт-ДНК гаплогруп, проте лише нащадки однієї з них – L3 вийшли за межі континенту в тому напрямку, що й носії чоловічої гаплогрупи СF, та розділились на 2 групи – М та N [71, 129, 152].
Перша хвиля людей, що рухалась в Індокитай мала переважно мт-гаплогруппу М. В Індії залишилась гаплогруппа N. Тому на заході Євразії переважють потомки носіїв гаплогруппи N, та її гілки R. На сході – є обидві лінії, що пов’язано з можливим поглинанням Індійською популяцією (носії чоловічої F-гаплогрупи) Індокитайців (носіїв чоловічої макрогруппи С та жіночої М), які в цей же час почали розселятися на острови Тихого океану.Це добре видно на наступній схемі, де серед народів заходу східні гаплогрупи мт-ДНК майже не зустрічаються.
В Західній Євразії, як ми бачимо, домінують вісім мт-гаплогруп – HV, H, V, U, K, I, W, J, T, X [130, 153]. При цьому в Сх. Європі (Україна, Росія, Білорусь) домінують H (40%), T (11-12 %), J (8%) [180]. Аборигенними тут є гаплогрупи – H, V, U, HV. В період неоліту зявилися з Бл. Сходу – J, T, K, з Ц. Азії – Х, I, W. Проте вони складають меншість. В Америці зустрічаються як східні гаплогрупи (D, C), так і західні (A, B, X). При цьому зрозуміло, що носійки Індійських за походженням гаплогруп разом з завойовниками також рухались на схід (мт-гаплогрупи – D,C, S, Y, P, F), розповсюджуючись на сході Азії, а потім і в Америці. Це доводить тезу про те, що корінне населення Америки має подвійне походження (Зах. Євразійське та Сх. Азійське). У відповідності до цього було дві групи переселенців. Перша – носії у-ДНК гаплогрупи – С3 та мт-ДНК – С, D (можливо через Полінейзію). Друга – з Алтаю. Таким чином ми можемо помітити, що в процесі розселення часто відбувається процес змішання носіїв різних гаплогруп, активна метисизація. Все це свідчить про певну відсутність чистих рас у людства. Мова йде про різні поєднання різних чоловічих та жіночих ліній на поверхні Землі, носії яких утворюють популяції, які мають комбіновані риси ще й внаслідок поєднання їх соматичного геному в процесі асиміляції.
Тому вірною може вважатися популяційна концепція людських рас у відповідності до якої існують певні континууми різних генів (ознак, ними індукованих) у просторі, що має досить нечіткі межі. Завдяки аналізу гаплогруп, до того ж, ми з’ясовуємо лише незначний етап розвитку людства, що не перевищує в даний час вік 325 тис. років (перевано 200-160 тис. років – вік більшості ліній). Проте у відповідності до данних, представлених на рис. 85 гени аутосом також активно мігрують між популяціями. Їх міграцію можна прослідити до більш давніх часів завдяки даним палеогенетики. Цікаво, що чоловічі лінії характеризуються більшою рухомістю, ніж жіночі, які мають більш осідлий характер. Наприклад, в Європі більшість жіночих ліній, як ми це уже спостерігали, мають походження від першопоселенців (близько 80 %), а ось чоловічі лінії – переважно лінії зайд (теж біля 80 % – мігранти з Ц. Азії).
В Європі люди з’явились близько 45 тис. р. тому, в Австралії – близько 50 тис. р. т., в Ц. Азії – 50 тис. р.т., в Америці – біля 14-16 тис. р. т. В Європі у відповідності до знань перша хвиля поселенців зявилась з Індії – 45 тис. р. т. (носії у-гаплогрупи І, у нас складають 20% чоловічого населення), друга – 10-12 тис. р. т. з Ц. Азії (R1a та G, відповідно 54% та 3%), третя – 5-6 тис. р. т. з Близького Сходу та Кавказу (R1b, J2, J1, Т відповідно 5-15%, 2-3%, 2-3%, 1-2%), четверта – 3,5 тис. р. т. з Півд. Уралу (N1c – 5-15%), п’ята – на межі нашої ери разом з гуннами з Ц. Азії (Q – 1-2%). Ще носії однієї з гаплогруп прибули в Європу з П. Африки, при посередництві греків (Е3в – 5%). Серед жіночих гаплогрупами прийшлих є мт-ДНК: Х, J, T (Ц. Азія та Бл. Схід). На даний момент не існує жодного народу, який би характеризувався лише однією із цих гаплогруп по чоловічій та жіночій лінії (рис. 86) [1, 10, 11, 153].

Рис. 85. Міграція генів людей в ойкумені за А. Темплтоном[42].

Рис. 86. Розподіл гаплогруп у-хромосом серед чоловічого населення Європи.
[153].
Це вже результат вторинного розселення людей та їх тривалої метисизації. Зразком цього є їх розподіл гаплогруп у-хропосом в Європі.
Після активного первинного розселення, у мезоліті клімат став більш теплим, з'явилися нові знаряддя, човни, весла, лижі, сани тощо. Тривалість життя збільшилася в жінок до 35 років, а у чоловіків – до 50. Під час неоліту розвивалися землеробство і скотарство, близько 7-9 тис. р. тому на Близькому Сході з'явилися міста (Ієрихон, Чатал-Гююк, Чайєню-Тапезі), а 6тис. р. тому перші держави (Шумер, Єгипет).

