




Тот факт, что реакция навязывания ритма в принципе подчиняется «закону силы», стал известен после работ некоторых авторов, изучавших динамику этого биоэлектрического явления в условиях изменяющейся интенсивности мелькающего светового раздражителя (М.Н. Ливанов, 1944; В.А. Ильянок, 1959, 1961; Н.Н. Данилова, 19616, 1963; и др.). В их работах можно найти образцы записей и диаграммы, свидетельствующие о том, что увеличение яркости стимула приводит к возрастанию амплитуды электрических ответов коры и повышению коэффициента синхронизации. Однако определение общего вида зависимости корковой реакции на мелькающий свет от интенсивности раздражения в литературе отсутствовало. В то же время из тех индивидуальных или «типичных» графиков этой зависимости, которые приводятся упомянутыми и – другими авторами, явствует, что индивидуальные различия в общем эффекте навязывания и в соблюдении правила силы могут быть весьма значительны, однако авторы воздерживаются от какой-либо интерпретации этих различий по их возможному механизму.
Все это побудило нас провести экспериментальную, работу (В.Д. Небылицын, 1964), целью которой было, во-первых, определить общий характер зависимости реакции навязывания ритма от интенсивности стимула, а во-вторых, установить ту роль, которую играет сила нервной системы в индивидуальных вариациях проявления этой зависимости.

Рис. 51. Энергия частотных полос ЭЭГ при действии непрерывного светового раздражителя. Ось абсцисс – интенсивность раздражителя (в лк); ось ординат – суммарная энергия, (усл. ед.) (В.Д. Небылицын, 1964).
Аппаратура, методика опыта и способ получения количественных индикаторов реакции навязывания (энергетических индексов навязывания ритма – ЭИНР) уже были описаны раньше (см. гл. V). Здесь мы напомним только, что мы пользовались анализатором частот, позволявшим выделить из ЭЭГ и регистрировать тета-, альфа- и бета-активность (см. образцы записи на рис. 18–21), и интегратором биотоков, дававшим возможность выражать в количественной форме эффект действия мелькающего светового раздражителя. Напомним также, что получаемые индексы (ЭИНР) представляют собой разность между энергией колебаний, измеренной в данной частотной полосе при действии ритмического светового раздражителя (например, в тета-полосе при мельканиях частотой 6 имп/с или в бета-полосе при частоте стимуляции 20 имп/с), и энергией колебаний, измеренной предварительно в той же частотной полосе при действии непрерывного света; таким образом, ЭИНР представляет собой оценку «чистого» эффекта ритмической световой стимуляции. Первоначально при подсчете ЭИНР для отдельных интенсивностей мы вычисляли указанные разности между эффектом ритмической стимуляции данной интенсивности и эффектом непрерывной стимуляции той же самой интенсивности. Впоследствии мы убедились, однако, что наше «вычитаемое» – энергия частотной полосы при действии непрерывного света – в пределах использованного нами диапазона интенсивностей (25, 50, 100, 200, 500 и 1000 лк от фотостимулятора ЭФС‑01) практически мало зависит от яркости стимула (рис. 51). Поэтому в качестве вычитаемого мы стали использовать при подсчетах среднюю энергию колебаний данной частотной полосы, вычисленную из результатов предъявления всех шести интенсивностей. По полученным таким образом индексам усвоения ритма строились соответствующие кривые.
Остановимся сначала вкратце на общих данных, получаемых путем простого усреднения показателей.
Динамика средних для всех 25 испытуемых численных значений индексов навязывания ритма как функции интенсивности раздражителя показана на рис. 52, где представлены кривые навязывания для каждой частоты стимуляции в отдельности (все кривые выравне-ны по способу взвешенной скользящей средней). Видно, что наименьшим наклоном обладают кривые навязывания частот, лежащих в диапазоне тета-ритма, особенно 5 и 6 имп/с, а также двух частот, относящихся к альфа-ритму, – 9 и 10 имп/с. Возрастание интенсивности с 25 до 1000 л к, т. е. в 40 раз, при данных частотах стимуляции почти не сказывается на приросте биоэлектрического эффекта. Кривая частоты 10 имп/с обнаруживает даже тенденцию к снижению. Наиболее крутым подъемом обладают кривые частот 11, 12 и особенно 18 и 20 имп/с. В настоящее время трудно сказать, являются ли эти различия закономерными или только случайными, свойственными лишь данной выборке испытуемых. Учитывая, однако, что эта выборка не так уже мала, можно думать, что в основе различий в крутизне кривых лежит действие физиологически существенных факторов.
Те же тенденции сохраняются и при усреднении кривых согласно частотным диапазонам (путем суммирования индексов соответствующих частот и последующего деления сумм на число частот, входящих в данную полосу). На рис. 53 приведены эти кривые; при этом мы даем две раздельные кривые для частот, соответствующих низкочастотному и высокочастотному альфа-ритму (9 – 10 и 11 – 12 имп/с), так как, согласно описанным в гл. V данным, эффекты навязывания двух этих групп частот, по-видимому, определяются различными факторами мозговой деятельности. Моментом, общим для всех кривых, является уменьшение прироста по мере увеличения интенсивности стимуляции (следует иметь в виду, что на всех упомянутых графиках ось абсцисс дана в логарифмированном виде); это заставляет предположить, что общим правилом динамики навязывания ритма как функции интенсивности является приближение к асимптоте.

Рис. 52. Зависимость реакции навязывания ритма от интенсивности стимуляции по отдельным частотам раздражения. Обозначения: а – тета-ритм, б – альфа-ритм, в-бета-ритм. Ось абсцисс – интенсивность раздражения (в лк); ось ординат – энергетические индексы навязывания ритма (усл. ед.) (В.Д. Небылицын, 1964).

Рис. 53. Зависимость реакции навязывания ритма от интенсивности стимуляции по отдельным физиологическим ритмам.
Значения абсцисс и ординат те же, что на рис. 52. (В.Д. Небылицын, 1964).

Рис. 54. Зависимость навязывания ритма от интенсивности стимуляции. Суммарная кривая.
Значения абсцисс и ординат те же, что на рис. 52 (В.Д. Небылицын, 1964).
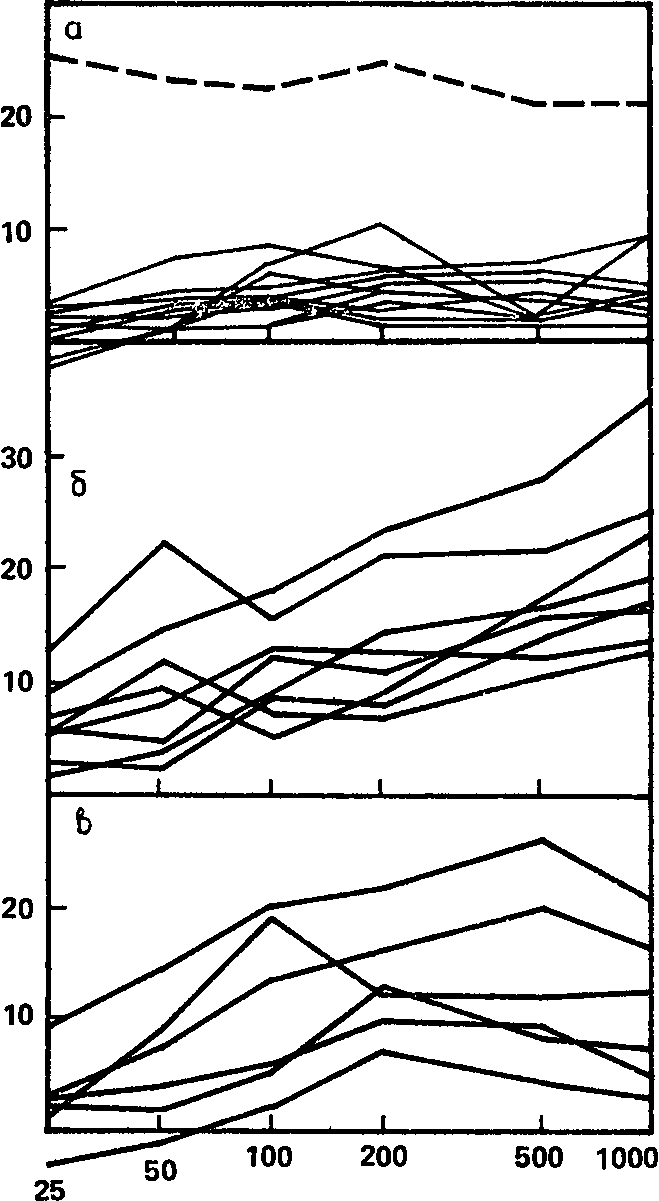
Рис. 55. Зависимость реакции навязывания ритма от интенсивности стимуляции. Индивидуальные графики, сгруппированные по трем основным типам реакций на возрастания яркости стимула (а, б, в).
Значения абсцисс и ординат те же, что на рис. 52 (В.Д. Небылицын, 1964).
Действительно, построив эмпирический ряд по среднесуммарным данным (т. е. в расчете на одну частоту стимуляции), мы находим, что точки этого ряда располагаются вдоль кривой, близко напоминающей асимптотическую функцию. Аналитическим выражением этой функции является равенство

Уравнение регрессии, параметры которого были вычислены в соответствии с этой формулой, приведено на рис. 54, где показаны также эмпирический ряд точек и построенная по этому ряду теоретическая кривая регрессии (шкала интенсивностей дается на этом графике в виде натурального ряда). Как видно, расчетная кривая проходит чрезвычайно близко к экспериментально найденным значениям.
Таким образом, судя по нашим данным, при использовании примененных нами техник анализа биотоков мозга и подсчета индексов навязывания ритма наилучшим аналитическим выражением зависимости навязывания ритма от интенсивности раздражения является асимптотическая функция. Экстраполирование полученной расчетной кривой в сторону более низких интенсивностей дает примерное значение нижнего порога навязывания ритма: оно лежит где-то около 10 лк. Что касается верхнего порога (предела функции), то, судя по кривой, он достигается уже при интенсивностях, равных приблизительно 400 – 500 лк.
Обратимся теперь к индивидуальным графикам, из которых, пожалуй, лишь некоторые более или менее точно соответствуют асимптотической функции.
Индивидуальные графики можно строить как по суммарным индексам, взяв средние для всех использованных частот, и по индексам для групп частот, соответствующих физиологическим ритмам, а также по индексам для отдельных частот стимуляции. Остановимся сначала на данных первого рода – кривых, отражающих индивидуальные различия в реакции навязывания ритма по среднесуммарной оценке этой реакции. Анализируя эти кривые, мы видим, что они по своему виду вполне отчетливо распадаются на три основные группы и что кривые всех испытуемых, кроме одного, могут быть отнесены к какой-либо из этих групп. Эта группировка показана на рис. 55.
Можно видеть, что для одного типа кривых (а) характерна ясно выраженная пологость и вместе с тем очень низкие ординаты: кривые идут практически параллельно оси абсцисс и на очень малой высоте над нею. Для другого типа кривых (б) характерен, напротив, довольно высокий прирост значений от низких интенсивностей к высоким, а также относительно высокие абсолютные значения ЭИНР, особенно при больших яркостях раздражения: кривые обладают значительной крутизной и заканчиваются на ординатах, в несколько раз превосходящих таковые для кривых первого типа. Наконец, в последнюю и самую немногочисленную группу (в) входят кривые, частью сходные с кривыми первого, а частью – с кривыми второго типа; однако они имеют одну характерную особенность, которая и позволяет выделить их в отдельную группу. Эта особенность – наличие ясно выраженного пика при одной из некрайних интенсивностей и последующее снижение кривой, обычно достигающее максимума при наибольшей интенсивности.
Для определения зависимости указанных особенностей кривых навязывания ритма от силы нервной системы было проведено сопоставление этих особенностей с результатами угашения с подкреплением (ЭЭГ вариант).
К сожалению, из 25 участвовавших в опыте испытуемых только 20 могли быть подвергнуты испытанию силы нервной системы при помощи ЭЭГ варианта угашения с подкреплением, у 5 остальных испытуемых альфа-ритм не был выражен настолько, чтобы у них можно было получить какие-либо условнорефлекторные ЭЭГ показатели.
Сопоставление у этих 20 испытуемых особенностей динамики ЭИНР и результатов угашения с подкреплением показало следующее (табл. 39):
Таблица 39
Сопоставление показателей силы нервной системы с типом кривой навязывания ритма (В.Д. Небылицын, 1964)
| Номер испытуемого | Сохранение условной реакции к концу угашения с подкреплением, в % к исходной величине | Тип кривой |
| Среднее | 131,5 | |
| Среднее | ||
| Среднее |
1) из 8 кривых первого типа («пологих») 5 принадлежат испытуемым, у которых в результате угашения с подкреплением условная реакция увеличилась (первая группа), а другие 3 – испытуемым с маловыраженным падением условного эффекта (с сохранением 70–84 % первоначального уровня условной реакции – вторая группа);
2) из 7 кривых второго типа («крутых») 4 также принадлежат испытуемым второй группы, а 3 других – испытуемым с сильным снижением реакции к концу угашения с подкреплением (третья группа);
3) 4 кривых третьего типа (с «перегибом») принадлежат испытуемым последней, третьей группы;
4) наконец, одна «нетипичная» кривая (обозначается пунктиром на рис. 55) была получена у испытуемого первой группы.
Из сопоставления следует, что между силой нервной системы и особенностями динамики реакции навязывания ритма при возрастании интенсивности стимула существует достаточно отчетливая зависимость.
Конкретные примеры проявления этой зависимости даются на рис. 56–59, представляющих собой кривые записи реакций биотоков мозга в ответ на предъявление ритмических световых импульсов возрастающей интенсивности. На рис. 56 и 57 показана динамика реактивного эффекта испытуемых с сильной, а на рис. 58 и 59 – испытуемых со слабой нервной системой при частотах, соответствующих тета- и бета-ритмам ‑7 и 16 имп/с. Как электроэнцефалографические кривые, так и графики, построенные по результатам интегрирования, свидетельствуют о наличии подчас весьма резких различий между «сильными» и «слабыми» испытуемыми.
Для испытуемых с сильной нервной системой характерны в целом кривые с малым эффектом и малым приростом эффекта навязывания на протяжении всего диапазона интенсивностей. Испытуемым со слабой нервной системой свойственны кривые с несколько большим эффектом навязывания уже при минимальной интенсивности стимула и с заметным возрастанием этого эффекта по мере увеличения интенсивности; кроме того, некоторые из «слабых» кривых, видимо, отражают момент достижения предела и перехода за этот предел (в виде снижения эффекта).
Эти различия иллюстрируются рис. 60, на котором изображены средние кривые для групп «сильных» и «слабых» испытуемых.
Вычисление корреляционного отношения  (эта), применяемого для измерения тесноты связи между двумя переменными в том случае, когда одна из них представляет собой градуальный ряд, а другая распределяется по нескольким качественным группам (P. Edwards, 1960), дает значение связи между кривой и силой нервной системы
(эта), применяемого для измерения тесноты связи между двумя переменными в том случае, когда одна из них представляет собой градуальный ряд, а другая распределяется по нескольким качественным группам (P. Edwards, 1960), дает значение связи между кривой и силой нервной системы  = 0,723 (р < 0,01). Таким образом, и статистический индикатор дает достаточно высокую меру связи между двумя сопоставляемыми характеристиками в том случае, когда для сравнения берутся кривые, усредненные по всем ритмам. Можно полагать, что на основе измерения ЭИНР как функции яркости мелькающего раздражителя может быть разработана достаточно «короткая» методика определения силы нервной системы. Мы проанализировали с этой целью кривые ЭИНР по отдельным физиологическим ритмам; для получения таких кривых обычно достаточно одного опыта с каждым испытуемым.
= 0,723 (р < 0,01). Таким образом, и статистический индикатор дает достаточно высокую меру связи между двумя сопоставляемыми характеристиками в том случае, когда для сравнения берутся кривые, усредненные по всем ритмам. Можно полагать, что на основе измерения ЭИНР как функции яркости мелькающего раздражителя может быть разработана достаточно «короткая» методика определения силы нервной системы. Мы проанализировали с этой целью кривые ЭИНР по отдельным физиологическим ритмам; для получения таких кривых обычно достаточно одного опыта с каждым испытуемым.


Рис. 56. (А, Б). Индивидуальная запись (исп. Щ., сильная нервная система). Стимуляция частотой 7 имп/с при различных уровнях интенсивности стимула не вызывает эффекта навязывания.
Обозначения: 1 – отметка раздражения, 2 – тета-ритм, 3 – альфа-ритм, 4 – бета-ритм, 5 – неанализированная ЭЭГ, 6 – отметка времени (1 с). Цифры справа обозначают результат интегрирования соответствующей частотной полосы (усл. ед.).


Рис. 57. (А, Б). Индивидуальная запись (исп. Щ., сильная нервная система). Стимуляция частотой 16 имп/с при различных уровнях интенсивности не вызывает эффекта навязывания. Обозначения те же, что на рис. 56.


Рис. 58. (А, Б). Индивидуальная запись (исп. К.О., слабая нервная система). Стимуляция частотой 7 имп/с вызывает отчетливо выраженную реакцию навязывания в тета-полосе, в целом возрастающую по мере увеличения интенсивности. Обозначения те же, что на рис. 56.


Рис. 59. (А, Б). Индивидуальная запись (исп. К.А., слабая нервная система). Стимуляция частотой 16 имп/с вызывает отчетливо выраженную реакцию навязывания в бета-полосе, возрастающую по мере увеличения интенсивности. Обозначения те же, что на рис. 56.

Рис. 60. Зависимость реакции навязывания от интенсивности раздражителя. Средние кривые для групп испытуемых с сильной и слабой нервной системой. Значения абсцисс и ординат те же, что на рис. 52 (В.Д. Небылицын, 1964).
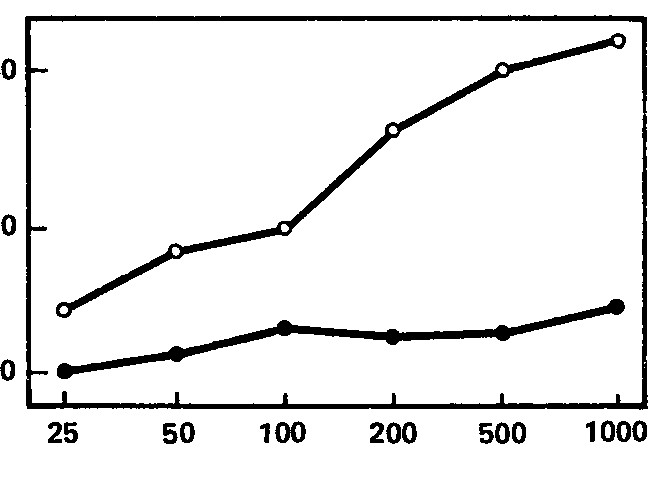
Рис. 61. Зависимость реакции навязывания ритма в тета-полосе от интенсивности раздражителя. Средние кривые для групп испытуемых с сильной (черные кружки) и слабой (белые кружки) нервной системой. Значения абсцисс и ординат те же, что на рис. 52.

Рис. 62. Зависимость реакции навязывания ритма в альфа-полосе от интенсивности раздражителя. Средние кривые для групп с сильной и слабой нервной системой.
Обозначения те же, что на рис. 61.

Рис. 63. Зависимость реакции навязывания ритма в бета-полосе от интенсивности раздражителя. Средние кривые для групп с сильной и слабой нервной системой.
Обозначения те же, что на рис. 61.
Эти кривые, для групп «сильных» и «слабых» испытуемых, показаны на рис. 61, 62, 63, из которых видно, что для частот, соответствующих тета- и особенно бета-ритму, кривые индивидов с сильной и слабой нервной системой существенно различны по своему характеру, особенно по степени наклона; правда, для диапазона альфа-ритма (рис. 62) различия между кривыми гораздо менее заметны. Отсюда вытекает, что для целей разработки короткой методики наибольшую эффективность обещает применение частот стимуляции, совпадающих с диапазоном тета- и бета-ритмов (по крайней мере, в пределах использованных нами частот). Причины же особого поведения диапазона альфа-ритма подлежат дальнейшему выяснению; возможно, они связаны со спецификой примененной методики оценки эффекта навязывания, тем более что другой метод оценки этой реакции, примененный Э.А. Голубевой, как мы увидим далее, дает при частотах полосы альфа-ритма довольно высокие величины связи с силой нервной системы.
Определенную перспективу в смысле разработки «короткой» методики обещает, как обнаруживается, и такой метод оценки реакции навязывания, как регистрация «кривых реактивности» по М.Н. Ливанову. Этот метод, состоящий в предъявлении индивиду ритмического светового раздражителя фиксированной частоты, но постепенно возрастающей яркости, удобен тем, что позволяет на протяжении одного «цуга» стимуляции, длящегося (в нашем случае) 38 с, наблюдать динамику реактивного эффекта на весь обычно применяемый диапазон интенсивностей светового раздражителя. При этом количественная оценка эффекта навязывания представляет определенные трудности, однако применение анализатора спектра позволяет все же выявить эту динамику с достаточной отчетливостью.
Опыт по определению индивидуальных особенностей «кривых реактивности» головного мозга и возможной зависимости их динамики от параметра силы нервной системы был поставлен нами с использованием в качестве источника световых импульсов постепенно возрастающей яркости фотостимулятора ЭФС‑01. От каждого из 25 испытуемых было получено 11 «кривых реактивности» – по числу примененных в опыте частот стимуляции, указанных в гл. V.
Сопоставление этих кривых с результатами угашения с подкреплением, основанное на визуальном анализе записей ЭЭГ составляющих и их изменений, позволяет, на наш взгляд, сделать вполне обоснованный вывод о существовании зависимости между силой нервной системы и проявлением эффекта навязывания в «кривых реактивности» коры больших полушарий.
Для испытуемых со слабой нервной системой оказалось характерным наличие эффекта навязывания почти на каждой из примененных частот стимуляции, кроме, может быть, минимальной ‑5 имп/с. При этом во многих случаях вершина эффекта наблюдалась не при максимальной яркости стимула, а при некоторых средних ее значениях. У испытуемых с сильной нервной системой реактивный эффект при данном способе стимуляции имел место гораздо реже, большей частью лишь в отдельных частотных диапазонах, а у некоторых из этих испытуемых он отсутствовал совершенно.
Мы не можем здесь за недостатком места сколько-нибудь широко иллюстрировать конкретными записями эти различия между «сильными» и «слабыми» испытуемыми. Приведем лишь для сравнения по одной характерной для каждой из групп записи биоэлектрической активности и ее изменений в ходе предъявления мелькающего света частотой 7 имп/с и постепенно возрастающей яркости. У «сильного» испытуемого (рис. 64) наблюдается только реакция блокады всех ритмов ЭЭГ на протяжении всего цуга стимуляции; у «слабого» (рис. 65) – отчетливо выраженная реакция навязывания первой гармоники, особенно при сильных значениях яркости стимула, а кроме того, временами и третьей гармоники (см. полосу бета-ритма).
Таким образом, при любом способе выявления эффекта навязывания обнаруживается зависимость динамики этой реакции в ходе изменения интенсивности стимула от параметра силы нервной системы.
И снова для объяснения природы этой зависимости нам приходится привлекать правило взаимосвязи между силой нервной системы и чувствительностью. Как и в опытах с временем реакции, а также с критической частотой фосфена, причиной различного хода «сильных» и «слабых» кривых при увеличении стимуляции, видимо, являются различия в абсолютной чувствительности сильных и слабых нервных систем. Большая чувствительность слабой нервной системы обусловливает больший эффект навязывания по сравнению с сильной нервной системой при минимальной интенсивности стимула (рис. 60) и более выраженный прирост этого эффекта («опережение») при увеличении стимула, с более стремительным приближением к пределу. Меньшая чувствительность сильной нервной системы и соответственно меньшая физиологическая эффективность стимуляции ведут к тому, что у «сильных» даже максимальная из применяемых интенсив

Рис. 64. Индивидуальная запись (исп. К., сильная нервная система). Отсутствие реакции навязывания ритма при стимуляции по методу «кривых реактивности», с частотой 7 имп/с.
Обозначения: А – начало, Б, В, Г – продолжение записи. 1 – тета-ритм, 2 – альфа-ритм, 3 – бета-ритм, 4 – неанализированная ЭЭГ, 5 – отметка времени (1с). Стрелками обозначены моменты включения и выключения стимуляции.
ностей стимула дает результат, лишь немного превышающий результат «слабых» при наименьшей яркости раздражителя (рис. 60). Таким образом, и эксперимент с навязыванием ритма, как это явствует из приведенных материалов, подтверждает следствия, вытекающие из правила взаимосвязи между чувствительностью и силой нервной системы, а следовательно, и само это правило.
К этому можно добавить, что прямой подсчет корреляции между суммарными индексами навязывания и абсолютными порогами, произведенный Э.А. Голубевой (1965), а также по ее данным в рамках работы по сопоставлению «коротких» методик (В.Д. Небылицын и др., 1965) приводит к довольно высоким, во многих случаях статистически значимым величинам коэффициентов (табл. 40), указывающим на то, что большая чувствительность зрительного анализатора способствует лучшему проявлению эффекта навязывания. Напомним, что методика оценки реакции навязывания, применяемая Э.А. Голубевой, значительно отличается от той, которая была использована нами. Тем более существенным кажется тот факт, что и при этой методике зависимость между показателями чувствительности (а также силы нервной системы – см. табл. 40) и индексами навязывания (которые в этой работе служат косвенными показателями подъема кривой, так как представляют собой сумму индексов для трех различных яркостей стимула) выявляется с достаточной отчетливостью.

Рис. 65. Индивидуальная запись (исп. Ц., слабая нервная система). Отчетливо выраженная, имеющая максимум при средних значениях яркости стимула реакция навязывания ритма при стимуляции по методу «кривых реактивности», с частотой 7 имп/с. Обозначения те же, что на рис. 64.
Таблица 40
Коэффициенты корреляции между показателями силы нервной системы и индексами навязывания ритма (по данным Э.А.Голубевой; В.Д.Небылицын и др., 1965)
| Показатели силы нервной системы | Суммарные индексы навязывания | |||||
| общий | дельта | тета | альфа | бета | гамма | |
| Абсолютный зрительный порог (24 испытуемых) | –409 ** | –253 | –390 * | –424 ** | –473 ** | –338 |
| ЭЭГ вариант угашения с подкреплением (19 испытуемых) | –470 ** | –220 | –275 | –562 ** | –451 * | –224 |
Примечание. р < 0,1; **р < 0,05.
Некоторым дополнением к данным настоящего раздела могут еще служить результаты эксперимента по определению эффекта навязывания как функции интенсивности стимула у детей. Этот эксперимент, поставленный К. Войку (1964), не имел целью сопоставление данных с показателями силы нервной системы, а поэтому мы не можем сказать точно, связаны ли индивидуальные особенности полученных функций с силой нервной системы испытуемых-детей. Однако весьма любопытно, что общая кривая зависимости эффекта навязывания от интенсивности стимула (рис. 66) очень близка к той асимптотической функции, которая была получена ранее для взрослых (рис. 54), а индивидуальные кривые демонстрируют примерно тот же характер индивидуальных различий, что и у взрослых. Это дает основания думать, что и у детей индивидуальные вариации проявления закона силы в реакции навязывания ритма, возможно, связаны с силой нервной системы (через связь этого параметра с абсолютной чувствительностью).

Рис. 66. Зависимость реакции навязывания ритма от интенсивности стимуляции у детей. Суммарная кривая. Ось абсцисс – интенсивность раздражения (в лк); ось ординат – ЭИНР (усл. ед.).
Что можно сказать по поводу материалов, изложенных в настоящей главе?
В трех различных экспериментах мы стремились получить опытные данные по вопросу о том, будут ли наблюдаться различия между сильными и слабыми нервными системами в количестве и динамике изменения исследуемого эффекта при возрастании стимуляции от каких-то минимальных значений (близких к порогу данной функции) до максимальных (находящихся на уровне предела данной физиологической функции). По-видимому, можно считать установленным, что эти различия действительно имеют место. Для слабых нервных систем характерны больший начальный эффект, более быстрое приближение к пределу и более раннее достижение предела данной функции, в то время как сильные нервные системы характеризуются, напротив, меньшим эффектом при минимальных значениях стимула, более медленным приближением к пределу функции и более поздним достижением этого предела. Для нас не существует иного объяснения этого явления, чем то, которое исходит из положения о взаимосвязи между силой нервной системы и абсолютной чувствительностью, можно полагать, что именно разница в уровне абсолютных порогов обусловливает указанные выше различия между сильными и слабыми нервными системами в количестве и динамике реактивного эффекта. Таким образом, через проверку следствий, вытекающих из закономерности связи между чувствительностью и силой, мы пришли к новому подтверждению самой этой закономерности.
Разумеется, каждый раз, как мы начинаем решать эту проблему с помощью конкретной методики, в дело вмешиваются специфические факторы этой методики, способные подчас сильно исказить ожидаемые «правильные» соотношения. В трех исследованных методиках такими интерферирующими факторами были (гипотетически) скорость проведения возбуждения по волокну в методике с временем реакции, лабильность периферических нервных элементов зрительного анализатора в методике КЧФ, лабильность корковых элементов зрительного анализатора в методике с навязыванием ритма. Устранить или обойти влияние этих специфических факторов при изучении обсуждаемой проблемы не всегда удается, но учитывать этот момент необходимо хотя бы для того, чтобы не делать поспешных отрицательных выводов.
В начале этой главы был поставлен частный вопрос относительно «параллельности», или «пропорциональности», прироста эффекта у сильных и слабых систем в промежуточной зоне стимуляции. Более или менее определенные данные для ответа на этот вопрос получены только в эксперименте с временем реакции, в той серии, где сопоставляются средние кривые «сильных» и «слабых» испытуемых при стимуляции в единицах порога, и, поскольку кривые фактически совпали, ответ на этот вопрос должен быть, по-видимому, положительным. Две другие примененные нами методики не дают такой легкой возможности для решения этой проблемы аналогичным образом. В эксперименте с фосфеном ввести стимуляцию в единицах порога чрезвычайно трудно, так как величина порога сама зависит от частоты стимуляции; поскольку опыт заключается как раз в плавном изменении частоты импульсов по мере нанесения раздражения, определить последнее в. единицах порога не представляется возможным (по крайней мере, без существенных технических усовершенствований аппаратуры). Что же касается навязывания ритма, то здесь для предъявления стимула в единицах порога следовало бы иметь прибор, представляющий собой комбинацию адаптометра со стробоскопом, а таким прибором мы не располагаем. Таким образом, от окончательного – положительного или отрицательного – ответа на вопрос о «пропорциональности» прироста эффекта в промежуточной зоне стимуляции следует пока воздержаться.

