




¬ излагаемом ниже эксперименте было использовано известное психофизиологическое €вление, заключающеес€ в возникновении ощущени€ света при раздражении глаза электрическим током. Ёто ощущение и носит название фосфена. Ѕольшинством авторов принимаетс€, что нервным субстратом фосфена, во вс€ком случае при пороговых воздействи€х, €вл€етс€ второй нейрон сетчатки, состо€щий из бипол€ров, горизонтальных и амакринных клеток, расположенных на глубине нескольких дес€тков микрон под слоем рецепторов ( . Motokawa et al., 1957). “аким образом, измерение параметров реакций глаза на электрическую стимул€цию позвол€ет миновать фотохимический аппарат сетчатки и получить пр€мую информацию о состо€нии и функции вышележащих собственно нервных элементов (ј.». Ѕогословский, 1944; ѕ.ќ. ћакаров, 1952). — этой целью определ€етс€ порог электрического возбуждени€ глаза (реобаза), электрическа€ оптическа€ хронакси€, критический интервал дискретности, критическа€ частота мельканий (частота исчезновени€ фосфена) и некоторые другие показатели (ј.». Ѕогословский, 1944; ѕ.ќ. ћакаров, 1952; ≈.Ќ. —еменовска€, 1963; и др.). ѕо каждому из этих показателей наблюдаетс€ широкий диапазон индивидуальных различий. ћожно было предполагать, что такой индикатор, как порог фосфена при раздражении глаза одиночными импульсами посто€нного тока, €вл€€сь показателем порога ощущени€, обнаружит такую же св€зь с параметрами силы нервной системы, как и другие исследованные ранее пороги. ѕравда, прежние работы проводились при адекватных раздражени€х органов чувств, а электрическа€ стимул€ци€ не €вл€етс€ адекватной дл€ глаза (как и дл€ любого другого анализатора), но она давала возможность измерить непосредственно возбудимость нервных элементов анализатора, оставл€€ в стороне первичный рецепторный аппарат. ѕоэтому измерение порогов фосфена и сопоставление их с показател€ми силы нервной системы €вились одной из задач нашей работы.
ƒругой задачей Ц и притом основной Ц было определение зависимости критической частоты мелькающего фосфена ( „‘) от амплитуды раздражающих импульсов и сопоставление получаемых индивидуальных функций (Ђкривых „‘ї) с результатами определени€ силы нервной системы.
ритическа€ частота мельканий фосфена представл€ет собой феномен, весьма сходный с критической частотой мельканий ( „ћ), измер€емой при адекватном раздражении глаза ритмическими световыми импульсами. ќтличие „ћ от „‘ состоит в том, что вместо €сного сли€ни€ мерцаний в непрерывно свет€щеес€ п€тно испытуемый при исследовании „‘ отмечает при некотором значении частоты ритмических импульсов исчезновение мерцаний в поле зрени€ и вообще исчезновение ощущени€ света. ќтсюда, кстати, проистекают существенные методические сложности измерени€ „‘, обусловленные трудностью улавливани€ самих мерцаний в поле зрени€, особенно при интенсивност€х, близких к порогу, а также трудностью фиксации момента исчезновени€ мерцаний. Ќеобходима довольно длительна€ тренировка дл€ того, чтобы испытуемый научилс€ отдавать себе и экспериментатору €сный отчет о характере своих ощущений и давать при измерени€х устойчивые однообразные результаты.
|
|
|
явление исчезновени€ мелькающего фосфена характеризуетс€ одной существенной особенностью: оно обнаруживает зависимость от интенсивности стимула Ц от напр€жени€ электрического тока или от его силы. ѕоскольку это так, можно ожидать индивидуальных различий в динамике кривой „‘ в зависимости от величины порога данного испытуемого, а значит, от силы его нервной системы: индивиды с высокими порогами и сильной нервной системой должны демонстрировать кривые с медленным нарастанием эффекта и поздним приближением к предельным значени€м функции, в то врем€ как кривые испытуемых с низкими порогами и слабой нервной системой должны характеризоватьс€ стремительным ростом эффекта и ранним достижением предела. »ными словами, можно ожидать тех же самых про€влений динамики „‘ как функции интенсивности стимула в зависимости от силы нервной системы, которые наблюдаютс€ и в опыте со временем реакции.
ќднако феномен „‘ имеет весьма специфическое физиологическое содержание, и это обусловливает возможность других гипотез по поводу зависимости кривой „‘ от силы нервной системы. явление исчезновени€ фосфена при определенной частоте импульсов может рассматриватьс€ как частное выражение пессимума ¬веденского специально в работе зрительного анализатора (Ќ.¬. —еменов, ¬.¬. оноплина, 1937; ≈.Ќ. —еменовска€, 1963). ѕоскольку пессимум Ц это феномен, в каком-то смысле родственный €влению запредельного торможени€, можно в принципе предполагать, что индивидам с сильной нервной системой, возможно, свойственны более высокие значени€ частот, при которых достигаетс€ пессимальный эффект, а дл€ Ђслабыхї эти значени€ будут ниже (ƒ.ѕ. Ќеумывака- апустник, ј.». ѕлаксин, 1964), и тогда при изучении критической частоты мелькающего фосфена можно было бы ожидать в противоположность гипотезе, сформулированной выше, более высоких значений „‘ (более высокой лабильности) у испытуемых с сильной нервной системой, меньших значений этого показател€ Ц у индивидов со слабой нервной системой.
Ќельз€ исключить также того, что оба указанных фактора Ц чувствительность и лабильность, Ц облада€, как видно, разнонаправленным действием, могут интерферировать между собой и как бы взаимно погашать друг друга. ¬ этом случае, по всей видимости, никаких €сных зависимостей между силой нервной системы и динамикой кривых „‘ получить не удалось бы.
— целью проверки высказанных предположений был организован эксперимент, в котором показатели порогов фосфена и кривые „‘ при изменении интенсивности стимула были сопоставлены с характеристиками силы нервной системы (¬.ƒ. Ќебылицын, 1960в).
ћетодическа€ сторона опыта заключалась в следующем.
ѕороги фосфена определ€лись при помощи импульсного электронного стимул€тора »—Ё Ц 0,1, генерирующего пр€моугольные импульсы. ƒл€ исследовани€ „‘ использовалс€ хронаксиметр производства мастерских »нститута физиотерапии, который удобен тем, что позвол€ет плавно измен€ть частоту подаваемых импульсов от одного до нескольких сотен в секунду при раздельной регулировке длительности импульса и частоты стимул€ции.
ќпыты проводились в звуконепроницаемой камере с черными стенами, при слабой освещенности пор€дка 0,002 лк (глаза испытуемого в течение всего опыта были открыты). ќпыт начиналс€ через 10 мин после начала адаптации, после того как понизивша€с€ электрическа€ чувствительность глаза устанавливалась на посто€нном уровне.
|
|
|
—еребр€ные электроды, обернутые влажной ватой, укрепл€лись: активный Ц над правой бровью испытуемого, индифферентный Ц на ладони одноименной руки.
¬ начале опыта путем предъ€влени€ одиночных импульсов возрастающей амплитуды производилось измерение порогов фосфена. Ёта процедура повтор€лась 3 Ц 4 раза, после чего выводилось среднее значение порога.
ќпределение критической частоты мелькающего фосфена производилось при нескольких фиксированных значени€х интенсивности импульса, начина€ от 3 ¬ и выше, ступен€ми по 2 ¬; длительность импульсов сохран€лась посто€нной и была равна 7 мс.
»змерени€ происходили следующим образом. ”становив минимальную интенсивность тока (3 Ц 5 ¬ в зависимости от измеренной перед этим реобазы) и подав сигнал Ђ¬нимание!ї, экспериментатор включал ток и вращал ручку плавной регулировки частоты импульсов до тех пор, пока испытуемый ни сообщал об исчезновении световых мерцаний; повторив измерение еще 2 Ц 3 раза, экспериментатор переходил к следующей интенсивности. “аким образом, крива€ измерений „‘ в зависимости от напр€жени€ импульса строилась в данном опыте по 3 Ц 4 измерени€м на каждой ступени интенсивности.
— каждым испытуемым было проведено 6 Ц 10 опытов; показатели из 4 последних опытов брались дл€ вычислени€ средней реобазы, средней хронаксии и средней кривой интенсивности Ц частоты.
¬ качестве испытуемых в опытах прин€ли участие 18 человек обоего пола в возрасте от 18 до 45 лет. »спытуемые поступали на опыты после определени€ у них силы нервной системы.
—ила нервной системы испытывалась при помощи четырех индикаторов. ќдним из них было угашение с подкреплением условного фотохимического рефлекса, причем в качестве условного сигнала использовалс€ световой раздражитель красного цвета. “рем€ другими были три варианта индукционной методики: Ђутомлениеї, Ђповторениеї и Ђформа кривойї. роме того, у всех испытуемых в отдельных опытах определ€лись показатели абсолютной зрительной чувствительности. оэффициенты коррел€ции рангов между всеми силовыми индикаторами были достаточно высоки Ц от 0,63 до 0,87.
ѕрежде чем перейти к изложению результатов основного эксперимента, остановимс€ на соотношении индикаторов силы нервной системы и электрической чувствительности глаза.
¬ табл. 37 приведены коэффициенты коррел€ции рангов между порогами электрической и световой чувствительности глаза и силой нервной системы наших испытуемых, определенной по четырем методикам. ак видно из таблицы, коэффициенты в двух случа€х из четырех (при Ђутомленииї и Ђповторенииї) значительно выше, когда показателем возбудимости зрительного анализатора €вл€етс€ реобаза, т. е. когда измер€етс€ непосредственно возбудимость нервных элементов анализатора, в одном случае Ц Ђформа кривойї Ц коэффициенты одинаковы, и только угашение с подкреплением несколько лучше коррелирует с абсолютной зрительной чувствительностью. “аким образом, преимущество в коррел€ции лежит в целом на стороне реобазы. Ёто св€зано, по-видимому, как раз с тем Ђобходом периферииї, который осуществл€етс€ при использовании электрической чувствительности глаза в качестве общего показател€ возбудимости анализатора. оэффициенты коррел€ции между порогами и силовыми показател€ми указывают на достаточно €сную тенденцию, котора€ подтверждаетс€ применением и других статистических критериев. “ак, оценка значимости различи€ между средними значени€ми порогов электрической чувствительности Ђсильныхї (2,9 ¬) и Ђслабыхї (2,1 ¬) испытуемых (по данным угашени€ с подкреплением) свидетельствует о статистической значимости этого различи€ на уровне р < 0,01.
“аблица 37
оэффициенты коррел€ции рангов между абсолютными порогами зрени€, порогами при электрической адекватной стимул€ции и силой нервной системы по данным четырех методик (¬.ƒ. Ќебылицын, 1960в)
| ”гашение с подкреплением | Ђ”томлениеї | Ђѕовторениеї | Ђ‘орма кривойї | |
| –еобаза | 59* | 74*** | 54* | |
| јбсолютный зрительный порог | 68** | 51* | 53* |
ѕримечание. Х р < 0,05; ** р < 0,01; ***р < 0,001.
|
|
|
—ледовательно, положение о наличии взаимосв€зи между чувствительностью и силой нервной системы находит себе подтверждение не только при применении адекватных раздражителей, но и в том случае, когда в качестве показател€ чувствительности нервных клеток выступает электровозбудитель зрительного индикатора.
–ассмотрим теперь результаты сопоставлени€ с силой нервной системы показателей динамики кривых „‘ как функции величины стимул€ции. ¬се индивидуальные кривые изменени€ критической частоты мелькающего фосфена в зависимости от амплитуды импульса показаны на рис. 50. ѕо данным угашени€ с подкреплением все испытуемые могли быть распределены по двум основным группам Ц Ђсильныхї и Ђслабыхї, а кроме того, двое испытуемых с промежуточными данными составили группу Ђпромежуточныхї.
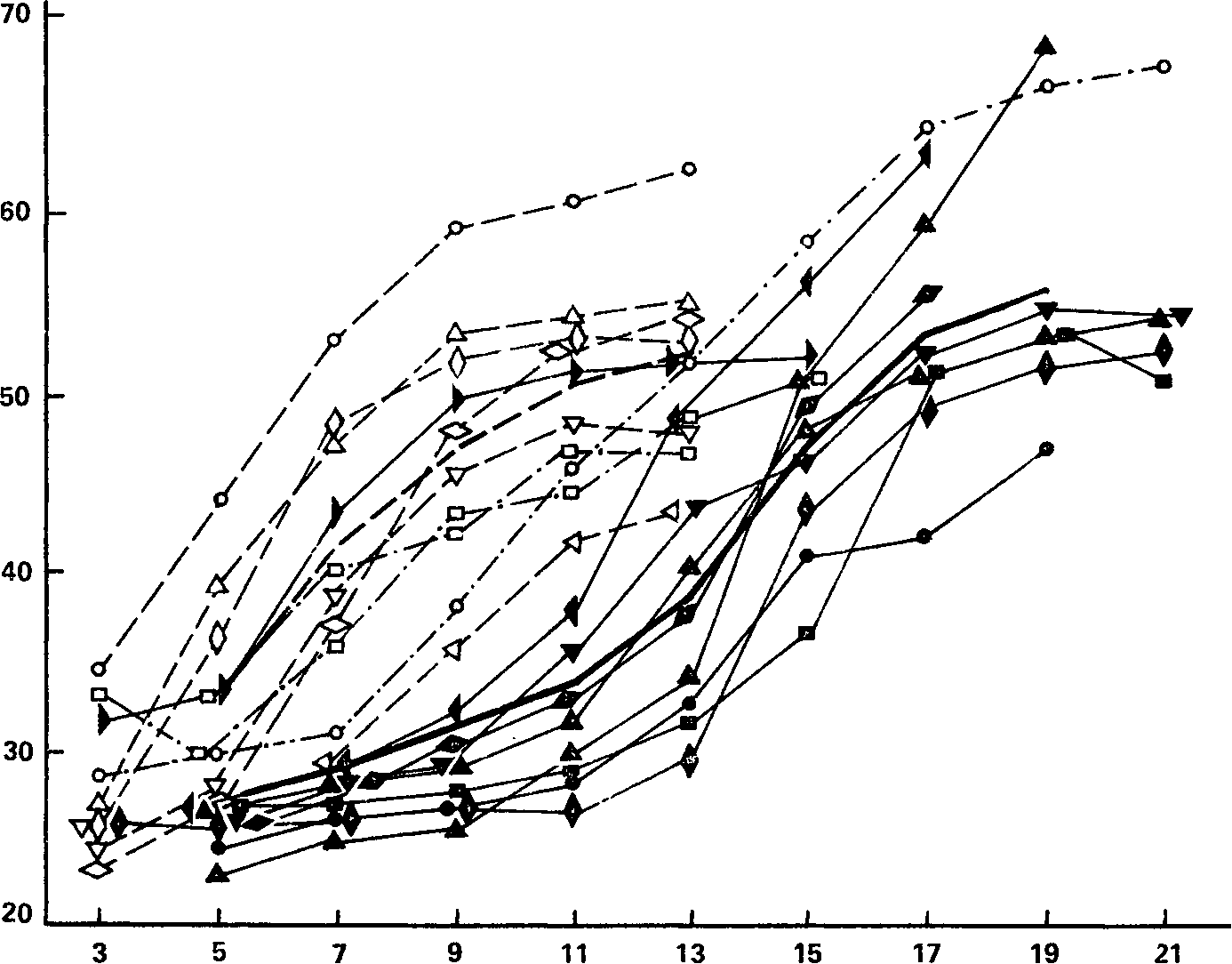
–ис. 50. ритическа€ частота мелькающего фосфена ( „‘) как функци€ интенсивности раздражающего электрического тока. —плошные. линии принадлежат испытуемым с сильной нервной системой; штриховые линии Ц испытуемым со слабой нервной системой; штрихпунктирные линии Ц испытуемым с промежуточным уровнем силы нервной системы. ∆ирные сплошна€ и штрихова€ линии Ц соответствующие средние.
ќсь абсцисс Ц напр€жение (в ¬); ось ординат Ц значени€ „‘ (¬.ƒ. Ќебылицын, 1960в).
ривые испытуемых с сильной нервной системой на рис. 50 обозначены сплошными лини€ми, со слабой Ц штриховой, Ђпромежуточныхї Ц штрихпунктиром. аждую из этих кривых можно характеризовать двум€ показател€ми: во-первых, пределом частоты, т. е. той максимальной высотой, которой достигает крива€ при постепенном увеличении интенсивности тока и за которой она у большинства испытуемых практически становитс€ горизонтальной, и, во-вторых, пределом интенсивности, т. е. той величиной раздражител€, при которой достигаетс€ частотный предел кривой данного испытуемого. –азличи€ по первому показателю Ц предельной величине усвоенного ритма Ц мы пока оставим в стороне, поскольку они, по-видимому, не завис€т от силы нервной системы: высота кривой в момент приобретени€ ею пологости (котора€, впрочем, достигнута не у всех испытуемых) может быть самой разнообразной у испытуемых как с сильной, так и со слабой нервной системой. „то же касаетс€ различий по второму показателю, то они, видимо, пр€мым образом св€заны с силой нервной системы, и это видно уже из непосредственного сравнени€ Ђсильныхї и Ђслабыхї кривых.
¬ самом деле, сплошные кривые, принадлежащие испытуемым с сильной нервной системой, имеют перегиб[35], демонстриру€ сначала положительное, а затем отрицательное ускорение, и приближаютс€ (кроме одной) к пределу лишь при интенсивности 17 Ц 21 ¬. Ќекоторые кривые Ђслабыхї испытуемых также показывают кратковременное положительное ускорение, но оно заканчиваетс€ уже в интервале 3 Ц 7 ¬, а при дальнейшем усилении тока эти кривые (как и другие Ђслабыеї кривые) быстро набирают высоту и приобретают пологость уже при напр€жени€х 11 Ц 13 ¬, т. е. уже в тот момент, когд^ кривые Ђсильныхї индивидов только еще начинают свой перегиб. “аким образом, в диапазоне 5 Ц 13 ¬ все кривые испытуемых со слабой нервной системой имеют отрицательное ускорение и большую высоту над осью абсцисс, а кривые всех испытуемых с сильной нервной системой, за исключением одного, имеют положительное ускорение и малую высоту над абсциссой. ¬ чем причина этой противоположности вида кривых в данном диапазоне стимул€ции?
Ћогично было бы предположить, что причина здесь заключаетс€ в различной физиологической действенности физически равных раздражителей, падающих на различные по своей чувствительности сильные и слабые нервные системы (вспомним, что это последнее различие выше было подтверждено статистически).
|
|
|
” более чувствительных слабых нервных систем один и тот же раздражитель (в нашем материале ток напр€жением 9 Ц 13 ¬) вызывает эффект, близкий к предельному, у менее чувствительных сильных нервных систем Ц эффект, от предельного еще весьма далекий. ќбщим результатом этой разницы и €вл€етс€ то, что в одном и том же диапазоне раздражителей (5 Ц 13 ¬) Ђсильныеї и Ђслабыеї кривые изменени€ „‘ имеют существенно различную высоту над осью абсцисс.
Ёти различи€ весьма хорошо коррелируют с силой нервной системы. ¬ табл. 38 представлены коэффициенты коррел€ции рангов между частотой исчезновени€ мелькающего фосфена при четырех интенсивност€х раздражител€ ( „‘ 7, 9, 11, 13 ¬) и результатами определени€ силы нервной системы по четырем примен€вшимс€ методикам. »з таблицы видно, что эти коррел€ции достаточно высоки.
¬озвраща€сь к предположени€м, высказанным в начале этого раздела относительно роли силы нервной системы, а точнее, св€занных с нею характеристик чувствительности и Ц гипотетически Ц лабильности в динамике кривых „‘, следует, видимо, признать, что индивидуальные вариации функции „‘ при изменении интенсивности стимул€ции идут именно в том направлении, которое диктуетс€ взаимосв€зью между силой и абсолютной чувствительностью. ≈сли бы это было не так, если бы основную роль играла предполагаема€ св€зь между силой и лабильностью, то тогда, очевидно, распределение кривых Ђсильныхї и Ђслабыхї испытуемых на рис. 50 было бы существенно иным.
ќднако вли€ние фактора лабильности нервной ткани на характер кривых „‘ все же существует. ѕо-видимому, продуктом именно этого вли€ни€ €вл€ютс€ уже упом€нутые выше индивидуальные различи€ в частотном пределе кривых, €вно не завис€щие от силовой характеристики индивида. —уд€ по кривым, максимально достигаема€ испытуемыми частота исчезновени€ фосфена колеблетс€ в среднем около 50 Ц 55 имп/с, что хорошо совпадает с данными других авторов (≈.Ќ. —еменовска€, 1963). ћожно предполагать, что лабильность элементов зрительного анализатора, насколько она может быть измерена приемами определени€ „‘, €вл€етс€ независимым по отношению к силе нервной системы фактором мозговой де€тельности. этому вопросу позже мы еще вернемс€.
“аблица 38
оэффициенты коррел€ции рангов между силой нервной системы, определенной по четырем методикам, и критической частотой мелькающего фосфена на четырех ступен€х интенсивности раздражител€ (¬.ƒ. Ќебылицын, 1960в)
| ћетодики определени€ силы | „‘, ¬ | |||
| и | ||||
| ”гашение с подкреплением | 81*** | 80*** | 84*** | 74*** |
| Ђ”томлениеї | 83*** | 81*** | 82*** | 79*** |
| Ђѕовторениеї | 68** | 69** | 71** | 57* |
| Ђ‘орма кривойї | 63** | 60** | 61** | 52* |
ѕримечание. Х p < 0,05; ** р < 0,01; ***p < 0,001. 253
Ќовое подтверждение того, что крива€ „‘ при вариаци€х интенсивности импульса есть показатель силы нервной системы, было получено «. √. “уровской (19636) в работе по сравнительному изучению р€да индикаторов некоторых свойств нервной системы. ѕоказатель наклона кривой „‘ коррелировал в этой работе с двум€ вариантами индукционной методики и с другими силовыми индикаторами; почти все коэффициенты были статистически значимыми, два из них Ц на 1 %-ном уровне. ќднако порог фосфена в этой работе фактически не коррелировал ни с одним показателем силы нервной системы. ѕоследнее обсто€тельство можно объ€снить тем, что измерение порога фосфена в экспериментах «.√. “уровской проводилось лишь в двух-трех опытах, а этого дл€ определени€ такой нестабильной характеристики, видимо, недостаточно (напомним, что в наших опытах значение порога фосфена выводилось как среднее из 4 опытов, шедших уже после значительной тренировки).
ѕодвод€ итоги сказанному относительно роли силы нервной системы в функции „‘ при измерении интенсивности стимула, нетрудно заметить, что полученные здесь результаты в значительной степени аналогичны результатам эксперимента с временем реакции и так же, как последние, видимо, вполне удовлетвор€ют тем следстви€м из правила взаимосв€зи между чувствительностью и силой, которые были сформулированы в начале этой главы. ƒействительно, при малых Ц близких к среднему порогу Ц интенсивност€х раздражени€ у испытуемых с сильной нервной системой наблюдаетс€ значительно меньший реактивный эффект, чем у Ђслабыхї индивидов, затем Ц при возрастании стимула Ц Ђслабыеї опережают Ђсильныхї в движении функции к пределу и раньше достигают его, и лишь затем при значительно больших величинах стимула к пределу приближаютс€ кривые индивидов с сильной нервной системой. ¬се эти различи€ обусловлены различи€ми самих нервных систем по уровню абсолютных порогов.
|
|
|
Ќужно сказать, однако, что в одной из работ, специально проведенной с целью сопоставлени€ методики „‘ с другими индикаторами силы нервной системы, эта методика, как оказалось, не коррелировала ни с одним из этих индикаторов (¬.ƒ. Ќебылицын и др., 1965). ¬ объ€снение этого факта были выдвинуты несколько предположений, из которых, пожалуй, самое основательное заключаетс€ в том, что все силовые индикаторы в этой работе относились к слуховому анализатору, а методика „‘, естественно, €вл€етс€ индикатором силы нервных клеток зрительного анализатора. роме того, могло сыграть свою отрицательную роль и то обсто€тельство, что кривые „‘ получались в данной работе в одном лишь опыте, без вс€кой предварительной тренировки. Ќаконец, не исключено, что в данной выборке испытуемых могла про€витьс€ та тенденци€ Ђсильныхї испытуемых к более высокой лабильности (а Ђслабыхї Ц к более низкой), о которой говорилось в начале этого раздела и которой достаточно обозначитьс€ лишь у некоторых испытуемых, чтобы свести к нулю обычно наблюдаемую коррел€цию.
“аким образом, исследование показателей электровозбудимости глаза в аспекте проблемы, обсуждаемой в насто€щей главе, дает подчас недостаточно определенные результаты. Ёто, очевидно, во многом св€зано со спецификой самих исследуемых параметров этой психофизиологической функции, которые, с одной стороны, могут быть определены только в результате оценки испытуемым своих иногда трудноуловимых ощущений и в которых, с другой стороны, действуют, веро€тно, вли€ние целой группы специфических дл€ этой методики факторов.
ѕоследнее, надо полагать, справедливо и дл€ той физиологической функции, котора€ известна под названием нав€зывани€ ритма и котора€, нар€ду со временем реакции и критической частотой мелькающего фосфена, была использована нами дл€ проверки следствий, вытекающих из положени€ о св€зи между силой нервной системы и абсолютной чувствительностью.

