




В дистальный извитой каналец поступает гипото-ничная жидкость из петли Генле. Как правило, состав и объем канальцевой жидкости в этом отделе нефрона изменяется значительно. В отличие от более проксимальных отделов нефрона, клетки эпителия дистальных канальцев плотно прилежат друг к другу, что делает их относительно непроницаемыми для воды и натрия, за счет чего поддерживаются градиенты концентраций, сформированные петлей Генле. В дистальных канальцах реабсорбируется 5 % фильтруемого в клубочках натрия. Реабсорбция натрия обеспечивается за счет функционирования КауК+-зависимой АТФ-азы на обращенной к капилляру поверхности клеток, а на люминальной мембране осуществляется Na^/СГ-переносчиком. Реабсорбция натрия в дистальном канальце прямо пропорциональна его поступлению. Дистальные канальцы — основное место реабсорбции кальция; этот процесс регулируется паратиреоидным гормоном и витамином D.
Наиболее дистальный отдел канальца называют соединяющим сегментом. В этом участке, как и в более проксимальных, происходит опосредованная гормонами реабсорбция кальция, но в отличие от них здесь имеет место еще и регулируемая альдостероном реабсорбция натрия.
Собирательная трубочка
В собирательной трубочке реабсорбируется 5-7 % фильтруемого в клубочках натрия, в ней выделяют кортикальную и медуллярную части.
А. Кортикальная часть собирательной трубочки. Этот сегмент нефрона состоит из двух типов клеток: 1) главных клеток, секретирующих калий и участвующих в опосредованной альдостероном реабсорбции натрия; 2) вставочных клеток, ответственных за регуляцию кислотно-основного равновесия. Главные клетки реабсорбируют натрий с помощью электрогенной помпы, поэтому для поддержания электронейтральности должна происходить либо реабсорбция СГ, либо секреция K+.

|
Рис. 31-3. Реабсорбция фосфатов, глюкозы, аминокислот и электролитов в проксимальном извитом канальце. Отметим, что №+/Кт-зависимая АТФ-аза, поддерживая низкую концентрацию натрия внутри клеток канальце-вого эпителия, обеспечивает энергетические потребности для реабсорбции катионов
Рис. 31-4. Реабсорбция натрия и хлора в толстом сегменте восходящей части петли Генле. Белок-переносчик (расположенный на люминальной поверхности клетки петли Генле) функционирует, только если заняты все четыре места связывания. Концентрация хлора в канальцевой жидкости является фактором, ограничивающем скорость реабсорбции
|
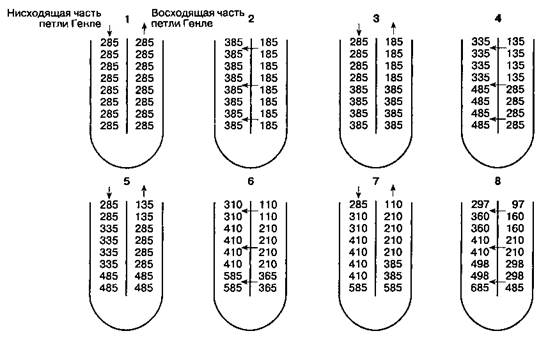
Рис. 31-5. Механизм противоточного умножения, обу слов лен ный различиями в проницаемости и трансмембранном транспорте, которые существуют между нисходящей и восходящей частями петли Генле. Нисходящая часть и тонкий сегмент восходящей части петли Генле проницаемы для воды, Na+, Cl и мочевины. Толстый сегмент восходящей части петли Генле непроницаем для воды и мочевины, в нем активно реабсорбируется Na+ и Cl. Эти факторы приводят к возникновению осмотического градиента. На рисунке представлен осмотический градиент между нисходящей и восходящей частями петли Генле, постепенно увеличивающийся от О до 200 мОсм/кг. Заметим, что в петле Генле по ходу тока канальцевой жидкости градиент не изменяется, в то время как осмоляльность увеличивается по мере приближения к точке перегиба. (По: Pitts R. F. Physiology of the Kidney and Body Fluids, 3rd ed. Year Book, 1974.)
Увеличение концентрации внутриклеточного калия способствует его секреции. В данном отделе нефрона альдостерон повышает активность NaV К+-зависимой АТФ-азы, увеличивая количество открытых K+- и Na^-каналов в люминальной мембране. Альдостерон также повышает активность ГГ-секретирующей АТФ-азы на люминальной поверхности вставочных клеток (рис. 31-6). Кроме того, во вставочных клетках люминальная Ю/Н+-зависимая АТФ-аза реабсорбирует K+ и секрети-рует ГГ. Некоторые вставочные клетки способны секретировать бикарбонат при большой щелочной нагрузке.
Б. Медуллярная часть собирательной трубочки — основная мишень антидиуретического гормона (АДГ, синоним — аргинин-вазопрессин). АД Г активирует аденилатциклазу через У2-рецеп-торы (активация V1-рецепторов повышает сосудистое сопротивление за счет усиления метаболизма фосфатидилинозитола). Проницаемость люминальной мембраны для воды полностью зависит от присутствия АДГ (гл. 28). Дегидратация приводит к повышению секреции АДГ, в результате
действия которого люминальная мембрана становится проницаемой для воды. По осмотическому градиенту вода удаляется из канальцевой жидкости, и образуется концентрированная моча (до 1400 мОсм/л). Напротив, адекватная гидратация подавляет секрецию АДГ; в этом случае осмоляльность канальцевой жидкости в медуллярной части собирательных трубочек не изменяется, и она остается гипотоничной (100-200 мОсм/л). В стенке медуллярной части собирательных трубочек содержатся основные и вставочные клетки, причем последние преобладают. Кроме того, эта часть нефрона отвечает за подкисление мочи; образующиеся в этих клетках ионы водорода выделяются в виде титруемых кислот (фосфаты) и ионов аммония (гл. 30). Предсердный натрийуретический пептид способен снижать реабсорбцию Na" в собирательных трубочках.
В. Роль собирательных трубочек в поддержании гипертоничности мозгового вещества. Кортикальная часть собирательных трубочек полностью проницаема для мочевины, тогда как медуллярная часть в норме для нее непроницаема. Этим разли-

Рис. 31-6. Кортикальная часть собирательной трубочки: секреция ионов водорода, реабсорбция бикарбоната и калия
чием в проницаемости практически наполовину обусловлена гипертоничность интерстициальной жидкости мозгового вещества. В присутствии АДГ значительно увеличивается проницаемость для мочевины наиболее дистальных отделов медуллярной части собирательных трубочек. Таким образом, при секреции АДГ вода покидает собирательные трубочки, и моча становится более концентрированной. Затем мочевина диффундирует в интерстициальную жидкость мозгового вещества, увеличивая ее осмоляльность.

