




Отдел базидиомикотовые грибы (Basidiomycota)насчитывает более 30 000 видов, которые характеризуются наличием вегетативного тела в виде многоклеточного мицелия (гаплоидного первичного и дикарионтичного вторичного) и полового спороношения в виде базидиоспор.
Монадные стадии отсутствуют. Основные компоненты многослойной клеточной стенки – хитин и глюканы. Септы между клетками простые (с простой порой) или долипоровые (с бочковидным расширением вокруг поры).
У базидиальных грибов встречается вегетативное, собственно бесполое и половое размножение. Вегетативное размножение осуществляется с помощью неспециализированных участков мицелия либо особых спор образующихся на дикарионтическом мицелии (эцидиоспоры, оидиоспоры, уредоспоры и т. Д.). Собственно бесполое размножение встречается достаточно редко и представлено конидиальным спороношением.
Половых органов у базидиальных грибов не образуется. Половой процесс, в основном, осуществляется путем слияния двух вегетативных клеток гаплоидного мицелия, вырастающего из базидиоспор (соматогамия). Встречается половой процесс в виде слияния базидиоспор или продуктов их почкования (Tilletia), слияния гаплоидных клеток базидии без образования базидиоспор (Ustillago), половой процесс в виде сперматизации (Puccinia).
У гомоталличных видов могут сливаться гифы одного и того же мицелия. У гетероталличных, к которым относится большинство базидиальных грибов, сливаются клетки гиф, берущих начало от спор противоположных половых знаков: «+» и «–». При этом происходит слияние цитоплазмы, а ядра объединяются в пары – дикарионы, которые затем синхронно делятся. Такой дикариотичный мицелий может существовать длительное время.
При синхронном делении пары ядер дикариона одновременно начинает формироваться пряжка в виде небольшого бокового выроста (рисунок 5.1, А-В), причем оси деления ядер лежат на одном уровне и вдоль клетки. В результате деления в клетке образуются четыре ядра. Сама клетка также делится перегородкой, разделяющей одну пару ядер: одно ядро оказывается в верхней (апикальной) клетке, другое – в нижней клетке (клетке-ножке) (рисунок 5.1, Г). Из оставшейся пары ядер одно остается в верхней клетке, а второе попадает в боковой вырост – пряжку, еще не отделенную перегородкой.
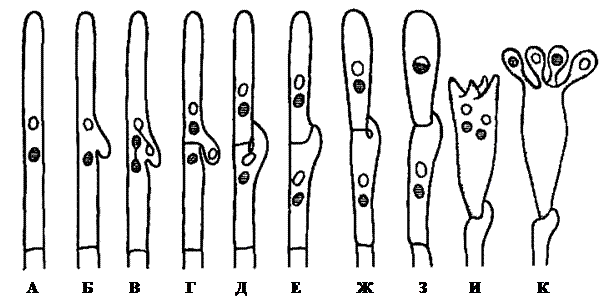
Рисунок 5.1 – Схема развития базидии
(объяснение буквенных обозначений в тексте)
В дальнейшем пряжка отделяется перегородкой и от апикальной клетки, а вершина ее, загибаясь книзу, прирастает к клетке-ножке. Между ними образуется отверстие, через которое ядро из пряжки переходит в клетку-ножку, восстанавливая ее двухъядерность (рисунок 5.1, Д-Ж). Благодаря этому клетка-ножка способна к дальнейшему функционированию: она может развивать дикариотичный мицелий и базидии. В верхней клетке также остается одна пара несестринских ядер. В дальнейшем она способна образовать базидию, при этом ядра сливаются (рисунок 5.1, Ж-З), и диплоидное ядро редукционно делится. Сама клетка обычно вытягивается, на ее вершине формируются выросты – стеригмы, которые, вздуваясь на конце, развиваются в базидиоспоры (рисунок 23, И). В каждую базидиоспору через узкий просвет стеригмы проникает по одному ядру (рисунок 5.1, К). При созревании базидиоспоры отбрасываются со стеригмы на небольшое расстояние.
По своему развитию базидия напоминает сумку и гомологична ей. Схема нарастания дикариотичного мицелия и развития базидии происходит с образованием пряжки, которая также гомологична структуре аскомикотовых грибов – крючку.
 |
Выделяют два типа базидий: холобазидии (одноклеточные) и фрагмобазидии (разделенные септами – продольными или поперечными). Ранее выделяемый тип гетеробазидия в настоящий момент рассматривается как одна из форм фрагмобазидии (рисунок 5.2).
Рисунок 5.2 – Типы базидий: А,Б – холобазидии; В,Г – фрагмобазидии
Холобазидии – одноклеточные базидии, чаще цилиндрической, булавовидной, овальной или неправильной формы. При этом если базидиоспоры располагаются на одном уровне, базидия является акроспоровой (характерна для грибов с активным отбрасыванием базидиоспор (рисунок 5.2)), а если на разных уровнях, беспорядочно – плевроспоровой (у грибов с пассивным распространением базидиоспор (рисунок 5.2, Б)).
Фрагмобазидия – базидия, разделенная поперечными перегородками на две или четыре клетки, на которых формируются базидиоспоры. Их структура разнообразна и специфична для таксонов.
У большинства базидиомикотовых базидии образуются на плодовых телах, сложенных из дикариотичного мицелия или внутри них. Реже они могут возникать прямо на мицелии или на особых покоящихся стадиях (телиоспорах или телейстоспорах).

Таким образом, в цикле развития базидиальных грибов наблюдается чередование гаплоидной, дикарионтичной и диплоидной ядерных фаз. В отличие от сумчатых грибов, преобладает дикарионтичная фаза: вторичный мицелий, плодовые тела (рисунок 5.3); гаплоидная фаза короткая: базидиоспоры и первичный мицелий, выросший из них; диплоидная фаза представлена молодой базидией (базидиолой).
Плодовые тела базидиальных грибов различны по форме и консистенции. Они бывают распростертые по субстрату – ресупинатные (паутинистые, рыхлые, плотно-войлочные, кожистые, в форме пленок, корочек), прямостоячие (палочковидные и коралловидные), копытообразные или состоящие из шляпки и ножки. У ряда видов плодовые тела отличаются своеобразным строением и иногда причудливой формой (гастеромицеты).
Спороносный слой плодового тела (гимений) у более примитивных видов располагается на верхней, а у более высокоорганизованных видов – на нижней стороне плодового тела (нижняя поверхность шляпки). Гимений базидиальных грибов состоит из базидии с базидиоспорами, базидиол (молодые или недоразвитые базидии) и стерильных клеток (парафиз), отделяющих базидии друг от друга и предохраняющих базидиоспоры от слипания. У некоторых видов в гимении имеются цистиды – крупные клетки, возвышающиеся над гимениальным слоем. Они защищают гимениальный слой и особенно базидии от давления сверху. Форма цистид для многих видов постоянна и часто служит систематическим признаком.
Поверхность плодового тела, несущая гимений, называется гименофором. У низших представителей он гладкий, у более высокоорганизованных имеет форму зубцов, складок, трубочек, пластинок.
В закрытых плодовых телах правильный гимений часто не образуется – на его месте формируется особая спороносная «ткань» (глеба), а базидии имеют разнообразную форму и разное расположение спор.
Гименофор базидиальных грибов может быть различно окрашен, что зависит в основном от цвета зрелых базидиоспор (от бесцветных, бледно-желтых и розовых до темно-фиолетовых и черных).
Большинство базидиальных грибов являются сапротрофами, микоризообразователями. Виды, ведущие паразитический образ жизни, встречаются реже, однако причиняют огромный вред сельскому и лесному хозяйству.
Отдел Basidiomycota по наличию плодовых тел, строению и образованию базидии, способностью спор к почкованию, строению септ принято делить на три класса:
1) класс базидиомицеты (Basidiomycetes) характеризуется наличием плодовых тел, всегда дифференцированных на стерильную и фертильную части. Базидии одноклеточные или септированные, развиваются на гимении или в глебе. Базидиоспоры не почкуются, а сразу прорастают в первичный гаплоидный мицелий. Половой процесс – соматогамия. Септы долипоровые с пареносомой (поровым мембранным колпачком);
2) класс телиомицеты (Teliomycetes), или ржавчинные грибы (Uredinomycetes). Плодовые тела отсутствуют. Базидия многоклеточная (фрагмобазидия), образуется из телейтоспоры. Базидиоспоры всегда прорастают гаплоидной гифой. Половой процесс протекает в виде сперматизации. Септы простые с открытой порой;
3) класс устомицеты (Ustomycetes) или головневые грибы (Ustillaginomycetes). Плодовые тела отсутствуют. Базидии (чаще разделенная перегородками фрагмобазидия, реже – одноклеточная) развиваются из толстостенной клетки – телиоспоры. Половой процесс протекает в виде копуляции гаплоидных клеток базидии, самих базидиоспор или продуктов их почкования. Септы простые, закрыты мелкими вакуолями.
Дополнение к разделу 5
Класс базидиомицеты(Basidiomycetes) включает большинство базидиальных грибов. Плодовые тела (карпофоры) закладываются на мицелии в виде зачатков (примордиев).
Систематика класса базидиомицеты является морфологической и построена на основании фенотипических признаков и цитологических признаков, основные их них: строение и способ образования базидии, тип и консистенция плодового тела, строение гимения, тип гименофора, наличие перфораций в парентосоме, наличие или отсутствие пряжки на дикарионтичном мицелии, орнаментация оболочки споры и ее химический состав.
В зависимости от сочетания перечисленных признаков выделяется до 14 порядков, которые объединяются в четыре несистематические морфологические группы:
– афилофороидные гименомицеты, включают порядки полипоральные (Polyporales), кантареллальные, или лисичковые (Cantarellales), телефоральные (Telephorales), гименохетальные (Hymenochaetales) и фистулинальные (Fistulinales);
– агарикоидные гименомицеты объединяют болетальные (Boletales), агарикальные (Agaricales) и руссулальные, или сыроежковые (Russulales);
– гастеромицеты: дождевиковые (Lycoperdales), ложнодождевиковые (Sclerodermatales), нидуллариальные, или гнездовковые (Nidulariales) и фаллальные, или веселковые (Phallales);
– фрагмобазидиомицеты со студенистыми плодовыми телами (ранее выделялись в таксон под названием гетеробазидиомицеты), которые объединяют порядки аурикулальные (Auricullales) и тремеллальные, или дрожалковые (Tremellales).
Необходимо отметить, что такое деление не в полной мере соответствует системе родственных отношений внутри класса, а больше отражает их морфологическую схожесть, сложившуюся конвергентно. Зачастую цитогенетический анализ постулирует значительно меньшее родство внутри перечисленных групп, чем между порядками разных морфологических групп. Так, компактную группу порядков формируют Agaricales (из агарикоидных гименомицетов), Nidulariales, Lycoperdales (порядки из группы гастеромицетов) и Fistulinales (из афилофороидных гименомицетов). Близкое родство имеет место между Boletales (агарикоидные гименомицеты) и Sclerodermatales (гастеромицеты). Одну филогенетически связанную группу составляют такие морфологически несхожие порядки, как Cantharellales, Hymenochaetales (афилофоровые гименомицеты) и Phallales (гастеромицеты). Обособленную позицию среди агарикоидных гименомицетов занимает порядок Russulales. В процессе эволюции наиболее удалились от других базидиомицетов порядки Auricullales и Tremellales.
Класс телиомицеты, или ржавчинные грибы (Teliomycetes, Uredinomycetes) облигатные паразиты высших растений, особенно мятликовых и бобовых. Плодовые тела у всех представителей класса отсутствуют вследствие паразитического образа жизни, многоклеточные базидии образуются из телейтоспоры (толстостенной многоклеточной покоящейся споры). Базидиоспоры прорастают гифой, половой процесс – сперматизация.
В составе ключевого порядка урединальные (Uredinales) насчитывается около 5 000 видов, в Беларуси отмечено более 400. У ржавчинных грибов известны несколько следующих друг за друec спороношений. Цикл развития ржавчинных грибов удобно проследить на примере возбудителя стеблевой, линейной, или черной ржавчины (Puccinia graminis), паразитирующей на барбарисе и на многих злаках
Другими представителями ржавчинных грибов являются пукциния скрытая (Puccinia econdite) – возбудитель бурой (листовой) ржавчины ржи и пшеницы, ржавчина малины (Phragmidium rubi-idaea), ржавчина рябины и ветреницы (Ochrospora sorbi) ржавчина льна (Melampsora lini), ржавчина сосны (Melampsora pinitorqua).
Класс устомицеты, или головневые грибы (Ustomycetes,или Ustilaginomycetes) включает базидиальные грибы, у которых отсутствует плодовое тело, базидии вырастают из толстостенной покоящейся клетки – устоспоры или телиоспоры (телиоспора по месту в жизненном цикле и ядерной фазе соответствует телейтоспоре ржавчинных грибов, зачастую между этими названиями ставится знак равенства). Данные споры являются фактором заражения растений, у большинства видов служат для перенесения неблагоприятных условий (в том числе для перезимовки).
Половой процесс представителей класса устомицеты протекает в виде копуляции базидиоспор, продуктов их почкования или в виде копуляции гаплоидных клеток базидии (без образования базидиоспор). В результате копуляции образуется дикарионтический мицелий. Появление таких форм полового процесса обусловлено паразитическим образом жизни головневых грибов и, как следствие, максимальной редукцией нежизнеспособных в этих условиях гаплоидных стадий. Данный тезис подтверждает тот факт, что полученные на стерильных искусственных средах головневые грибы существуют в гаплоидном состоянии.
Головневые грибы принято делить на два порядка: устилягинальные (Ustilaginales) и тиллециальные (Tilletiales). В последнее время к классу устомицеты все чаще относят порядок экзобазидиальные (Exobasidiales), в более старых системах причисляемый к классу базидиомицеты.

