




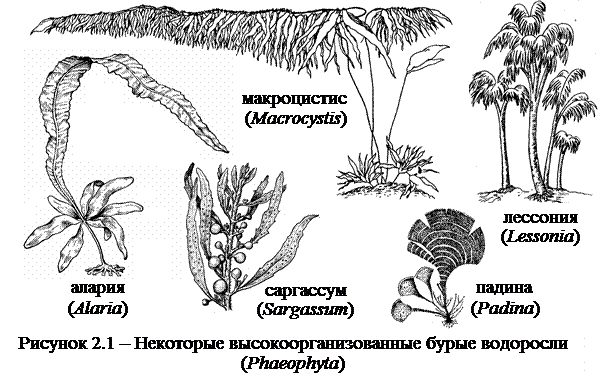
 Подавляющее большинство из около 2000 видов представителей отдела бурые водоросли (Phaeophyta) – морские, макроскопические организмы (рисунок 2.1).
Подавляющее большинство из около 2000 видов представителей отдела бурые водоросли (Phaeophyta) – морские, макроскопические организмы (рисунок 2.1).
В настоящий момент известно только восемь пресноводных видов, однако, ввиду их малых размеров и редкой встречаемости, они остаются слабо изученной группой.
Наиболее просто организованные талломы представлены однорядными беспорядочно ветвящимися нитями, иногда формирующими кустистые формы, основанием которых служат стелющиеся ризоиды (порядок Ectocarpales). У высокоорганизованных бурых водорослей талломы дифференцированы (тканевые) и напоминают высшие растения (Laminaria, Sargassum, Fucus). У них имеются корневидные, стеблевидные и листовидные части, некоторые крупные представители имеют воздушные пузыри, удерживающие ветви в вертикальном положении. В таких слоевищах наблюдается специализация клеток с образованием тканей (ассимиляционная, запасающая, механическая и проводящая) (рисунок 2.2). В простейшем случае в талломе различают кору и сердцевину. У более сложно организованных бурых водорослей (Laminariales) в коре выделяют меристодерму (делящуюся покровную ткань), в центральной бесцветной части – промежуточный слой и сердцевину. Сердцевина выполняет механическую функцию и функцию транспорта продуктов фотосинтеза. Транспортная функция осуществляется с помощью ситовидных трубок или трубчатых нитей. Часть трубчатых нитей идет поперек слоевища от клеток промежуточного слоя, другие ориентированы по длинной оси слоевища, при этом клетки последних очень длинные и узкие, и только у поперечных перегородок они воронкообразно расширены. В перегородках трубчатых нитей находятся поры, а сами перегородки называются ситовидными  пластинками.
пластинками.
Среди бурых водорослей имеются виды с однолетними и многолетними талломами. Многолетние бывают нескольких типов. У одних таллом целиком многолетний, ежегодно отмирают только побеги, на которых развивались органы размножения (Fucus). У других многолетними являются осевая часть и органы прикрепления (Laminaria). У третьих многолетний только базальный диск, а остальная часть таллома ежегодно отрастает вновь (Sargassum).
Увеличение слоевищ спорофитов в длину, в основном, обеспечивается деятельностью интеркалярной зоны, которая расположена в основании пластинчатой части слоевища, на вершине ствола (в более просто организованном нитчатом талломе может одновременно присутствовать как интеркалярный, так и верхушечный рост). В разветвленном таллом интеркалярная зона имеется в основании каждой пластины. Интеркалярная зона не имеет четкой границы, деление клеток слоевища происходит и на некотором расстоянии от нее. Наибольшая скорость роста таллома наблюдается у ламинариевых (до 10 – 13,5 см в сутки), но с ростом таллома одновременно происходят процессы разрушения его верхушки. Многолетние органы периодически нарастают в толщину за счет деления клеток коры, что приводит к формированию концентрических слоев, напоминающих годичные кольца высших растений (рисунок 2.3).
 |
Оболочка клеток бурых водорослей двухслойная. Внутренний слой целлюлозный, однако, целлюлоза бурых водорослей несколько отличается по свойствам от целлюлозы цветковых растений, и потому ее называют альгулезой. Внешний слой пектиновый, обычно состоит из белковых комплексов с альгиновой кислотой и ее солями. Благодаря такому строению оболочка бурых водорослей может набухать, превращаясь в слизистую массу иногда значительного объема.
Ядро в клетке одно, типично эукариотического строения. Митоз полузакрытый, веретено деления формируется центриолями.
Окраска талломов от желтовато-бурой до почти черной, обусловленной своеобразным набором пигментов (хлорофиллы а и с, β - и ε -каротины, а также несколько ксантофиллов – фукоксантин, виолаксантин, антераксантин и зеаксантин), среди которых наиболее специфичным является фукоксантин интенсивно бурого цвета. Пластиды вторично симбиотические родофитного типа, покрыты четырьмя мембранами. Две наружные мембраны структурированы в пластидную эндоплазматическую сеть, которая непосредственно переходит в ядерную оболочку. Две внутренние мембраны образуют собственно оболочку хлоропласта. Между второй и третьей мембранами есть перипластидное пространство. Тилакоиды внутри пластид расположены по три, имеется опоясывающая ламелла. Пластиды большей частью многочисленные, мелкие, дисковидные, реже лентовидные или пластинчатые. Пиреноиды характерны не для всех представителей. У подвижных стадий в пластиде имеется стигма. Запасные вещества (ламинарин, реже обнаруживаются масла и многоатомные спирты (например, econ)) откладываются за пределами пластид. Митохондриальный аппарат представлен разветвленным митохондрионом, внутренняя мембрана которого структурирована в трубчатые кристы.
Для бурых водорослей характерно бесполое и половое размножение. Вегетативное размножение специализированными выводковыми почками характерно только для некоторых бурых водорослей (род Sphacelaria). Вегетативное размножение путем фрагментации таллома нельзя считать безусловным. Оторвавшиеся участки талломов продолжают вегетацию только при попадании в наиболее благоприятные условия, не образуя при этом новых органов размножения. Исключением являются саргассовые водоросли (род Sargassum), обитающие в открытых акваториях Атлантического океана. Бесполое размножение у большинства бурых водорослей происходит с помощью разножгутиковых зооспор (жгутики прикреплены с брюшной стороны, двигательный направлен вперед, покрыт ретронемами, рулевой жгутик гладкий, направлен назад), у отдельных – неподвижными спорами, которые по количеству их в спорангии называются тетраспорами или моноспорами. Половой процесс изо -, гетеро - и оогамный.
Жизненный цикл гаплодиплофазный с чередованием поколений (изоморфным или гетероморфным, с регулярной или нерегулярной сменой поколений) или диплофазный без смены поколений.
Бурые водоросли являются основным поставщиком органического вещества в прибрежных экосистемах морей и океанов, макроскопические представители способны формировать подводные леса и плантации, что массово используется человеком. Бурые водоросли и продукты их переработки (альгинаты, фукоидаты, йод) нашли применение в сельском хозяйстве (кормопроизводство и производство удобрений), в пищевой, текстильной, целлюлозно-бумажной промышленности, в металлургии и радиоэлектронике. Изучается возможность их переработки в метан. В медицине они применяются не только как наполнители таблеток и гели-носители, но и как самостоятельные препараты (кровоостанавливающие субстанции и сорбенты токсических и радиоактивных элементов, в том числе стронция). Исследуются их противоопухолевые и противовирусные свойства.
Отдел включает один класс Феофициевые (Phaeophyceae) и восемь порядков. Деление на порядки осуществляется по следующим признакам: наличие пиреноида и его строение, особенности размножения, наличие зооспор, тип полового процесса, особенности протекания жизненного цикла. Наиболее характерные порядки отдела бурые водоросли: Эктокарпальные (Ectocarpales), Диктиотальные (Dictyotales), Ламинариальные (Laminariales) и Фукальные ил Фукусовые (Fucales). Разнообразие жизненных циклов можно рассмотреть на примере одноименных представителей данных порядков.
Род эктокарпус (Ectocarpus) характеризуется талломом в виде желтовато-бурых кустиков, достигающих в длину нескольких сантиметров (иногда до 30 см и более). Талломы состоят из стелющихся нитей, от которых отходят вертикальные, обильно ветвящиеся, однорядные нити. Рост нитей у большинства видов диффузный, лишь у некоторых обосабливается интеркалярная зона деления у основания боковых веточек (волосков). Еще реже встречается верхушечный рост. Ввиду этого длина клеток в нитях неодинакова: более молодые клетки – более мелкие. Прикрепление талломов осуществляется ризоидами, которые у крупных экземпляров образуют своеобразную кору у основания ветвей. К вершине ветвей клетки сужаются и заканчиваются длинным бесцветным волоском. Эти водоросли произрастают на скалах или на других водорослях и встречаются во всех морях мира, предпочитая холодные воды Севера. Размножаются бесполым и половым путем.
В жизненном цикле наблюдается нерегулярная изоморфная смена поколений (рисунок 2.4). Гаметофит и спорофит одинаковы по размерам и строению, но гаметофит гаплоидный и образует только многогнездные гаметангии, где формируются морфологически одинаковые гаметы.
Половой процесс – изогамия, но гаметы отличаются поведением: одни теряют подвижность (женские) и выделяют вещество, привлекающее другие (мужские) гаметы. После оплодотворения из зиготы вырастает диплоидный спорофит. На спорофите при температуре выше 20˚С образуются только нейтральные многогнездные спорангии, где без редукционного деления формируются диплоидные зооспоры, прорастающие в еще одно диплоидное поколение (спорофит). В диапазоне температур от 13 до 20˚С образуются и гаплоидные и диплоидные зооспоры. При температуре ниже 13˚С развиваются только одногнездные спорангии, в которых происходит редукционное деление, что приводит к формированию гаплоидных зооспор. Они прорастают в гаплоидные гаметофиты
Часто наблюдаются дополнительные отклонения от регулярной смены поколений (на схеме – пунктирные стрелки). Гаметы, образованные на гаметофите, могут функционировать как зооспоры и партеногенетически (без оплодотворения) прорастать в еще одно гаплоидное поколение. Поэтому многогнездные гаметангии также называются микроспорангиями. Кроме того, гаплоидные зооспоры, образующиеся на спорофите, могут вести себя как гаметы и попарно копулировать с образованием зиготы и, соответственно, диплоидного растения. Иногда, в дополнение к гаплоидным и диплоидным, наблюдается спонтанное появление и тетраплоидных
    |
генераций.
Регулярное чередование поколений наблюдается в жизненном цикле представителей рода диктиота (Dictyota) (рисунок 2.5).
Их таллом представляет собой трехслойное лентовидное образование с моноподиальным ветвлением. Однако, вследствие быстрого роста боковых ветвей, складывается впечатление дихотомического ветвления. Таллом развивается из цилиндрического ризома, прикрепленного к субстрату ризоидами. Внутренний слой таллома состоит из крупных сердцевинных клеток, бесцветных с немногими хроматофорами. С обеих сторон располагаются по слою клеток «коры», более мелких с многочисленными хроматофорами.
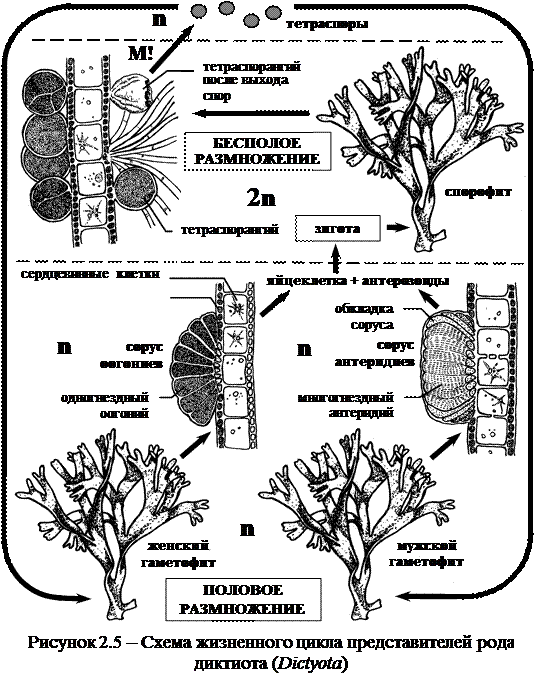 |
Жизненный цикл. На диплоидном спорофите диктиоты из поверхностных клеток «коры» развиваются шаровидные спорангии, где путем редукционного деления образуются по четыре неподвижные крупные тетраспоры. Тетраспоры прорастают в гаплоидный гаметофит. Диктиота – растение двудомное. Несмотря на однородность тетраспор, из них формируются мужские гаметофиты с сорусами антеридиев и женские гаметофиты с сорусами оогониев (группы оогониев). Антеридии имеют вид многокамерных вместилищ, в каждой камере которых формируется по одному одножгутиковому сперматозоиду (антерозоиду). Антеридии собраны в группы (сорусы антеридиев), которые покрыты обкладкой. Одногнездные оогонии (также собранные в сорусы, но без обкладки) образуют по одной яйцеклетке в каждом. Она выпадает из гаметангия, и в воде оплодотворяется антерозоидом. Образовавшаяся зигота покрывается оболочкой, и без периода покоя прорастает в спорофит.
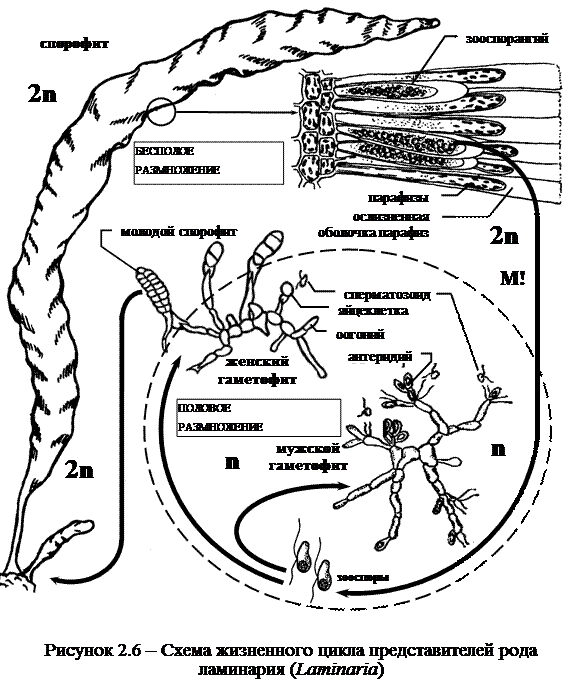 |
Род ламинария (Laminaria) или морская капуста имеет паренхиматозный тип таллома, здесь появляются настоящие ткани; в жизненном цикле наблюдается гетероморфное чередование поколений.
Жизненный цикл. Спорофит представляет собой листовидное слоевище с плотным стеблевидным черешком, прикрепленное к субстрату мощными когтевидными ризоидами. Листовая часть ежегодно сбрасывается и вырастает вновь за счет деятельности меристематических клеток, находящихся между черешком и пластинкой. На поверхности листовидной пластины формируются сорусы, состоящие из парафиз и зооспорангиев. Оболочка парафиз на вершине сильно ослизняется, образуя своеобразный толстый слизистый колпачок. Слизистые колпачки соседних парафиз смыкаются, благодаря чему получается сплошной толстый слой слизи, защищающий сорус. В зооспорангии развивается в зависимости от вида по 16-128 одинаковых зооспор. Первое деление ядра редукционное (рисунок 2.6).
Зооспоры выходят из зооспорангия, недолго поплавав, останавливаются и прорастают в микроскопические раздельнополые нитевидные гаметофиты (заростки). Мужской – ветвящийся с одноклеточными антеридиями, в каждом из них созревает по одному сперматозоиду. Женский заросток состоит из нескольких клеток, образующих короткую нить, он формируется в более благоприятных условиях. Половой процесс оогамный. Созревшая в оогонии яйцеклетка выходит и закрепляется на его верхнем конце. В таком положении и происходит оплодотворение, после которого зигота без периода покоя прорастает в спорофит. Таким образом, женский гаметофит обеспечивает место прикрепления будущего спорофита.
К бурым водорослям, не имеющим смены поколений и характеризующимся только сменой ядерных фаз, относятся представители рода фукус (Fucus). Их талломы ремневидные, дихотомически разветвленные, до 1 м длиной и до 5 см шириной (рисунок 2.7).
По середине таллома проходит толстая средняя жилка, которая в нижней части переходит в короткий стволик с подушкообразно расширенным основанием (базальным диском), с помощью которого и происходит прикрепление к субстрату. В верхней части многих видов по бокам жилок имеются воздушные пузыри, удерживающие талломы в вертикальном положении.
Жизненный цикл. Нормальное воспроизведение фукуса возможно только половым путем. На концах ветвей образуются вздутия (рецептакулы), в которых образуются скафидии – вместилища половых органов. Скафидии могут быть мужские и женские. Между оогониями в женском скафидии имеются парафизы, которые часто выходят за его пределы. В каждом оогонии образуется 8 яйцеклеток. В мужском скафидии антеридии размещаются на концах особых однорядных веточек, вырастающих из стенки скафидия, парафизы значительно короче. Необходимо отметить, что весь скафидий образуется из одной клетки (проспоры), но редукционное деление происходит непосредственно перед образованием гамет.
Существуют и другие точки зрения относительно особенностей смены ядерных фаз у данного представителя, однако, представленная схема жизненного цикла фукуса является классической.
 |
Рассмотренные типы жизненных циклов не охватывают всего их разнообразия у бурых водорослей. Так, например, существуют и виды с гетероморфным чередованием поколений и преобладанием макроскопического гаметафита, спорофит несколько меньших размеров (Cutleria). У большого числа бурых водорослей значительного разнообразия смен форм развития добавляют вариации жизненных циклов в зависимости от экологических условий и нерегулярности смены поколений.

