




Минералокортикоиды:
1) стимулируют активный транспорт Na+ в дистальных канальцах. Это обусловлено тем, что минералокортикоиды: а) увеличивают число Na+-каналов на апикальной стороне эпителиальных клеток; б) повышают количество и активность ряда митохондриальных ферментов, что способствует наработке молекул АТФ, необходимых для работы Na+/K+-насоса на серозной стороне клеток;
2) стимулируют выделение почками ионов калия. Секреция ионов калия в мочу из клеток дистальных канальцев запускается электрохимической движущей силой, создаваемой активной реабсорбцией ионов натрия.
3) влияют на транспорт ионов натрия и калия в других эпителиальных клетках: потовых и слюнных железах, слизистой кишечника.
Основная роль в реализации минералокортикоидных эффектов принадлежит альдостерону. Минералокортикоидная активность 11-дезоксикортикостерона в 25 раз ниже, чем у альдостерона.
Гормоны гипоталамуса, задней доли гипофиза.
Классификация гипоталамических нейрогормонов способность стимулировать (либерины) или ингибировать (статины) высвобождение соответствующего гормона гипофиза. К первой группе корти-колиберин –тиреолиберин –соматолиберингонадолиберинпролактолиберин –меланолиберин Ко второй группе относятся соматостатин,пролактостатин,меланостатин Все гипоталамические гормоны, это вещества пептидной природы. Гипоталамо-гипофизарные гормоны очень быстро исчезают из крови, так как не имеют специфических белков-переносчиков. Период их полураспада составляет несколько минут. Инактивация (распад) этих гормонов происходит в печени под действием специфических пептидаз. Окситоцин и антидиуретический гормон синтезируются в гипоталамических ядрах, пакуются в секреторные гранулы с белком нейрофизином и по аксонам нервных клеток доставляются в заднюю долю гипофиза. По строению оба гомологичными нона-пептидами, отличающимися по двум аминокислотным остаткам. Антидиуретический гормон (вазопрессин) синтезируется в супраоптическом ядре гипоталамуса. Главным стимулом секреции повышение осмолярности плазмы крови. Другими стимулами являются гиповолемия, гипотензия, боль, стресс и гипертермия. Ингибиторами -снижение осмолярности плазмы крови, этанол, гипотермия. Основное биологическое действие в повышении реабсорбции воды в дистальных канальцах почек путем активации мембрановстроенных белковых водных каналов – аквапоринов. Активация аквапоринов реализуется через связывание с поверхностнымиV2-рецепторами с образованием цАМФ. Вазопрессин обладает и метаболическим действием: 1) активирует распад гликогена; 2) стимулирует превращение глюкозы до ацетил-КоА;3) активирует синтез жирных кислот. Кроме того, вазопрессин обладает мощным антикетогенным действием.Дефицит приводит к развитию несахарного диабета характерными признаками которого являются:– постоянная жажда; – обильное мочеиспускание;– низкая плотность мочи– отсутствие в моче глюкозы. Окситоцин первично образуется в паравентрикулярном ядре гипоталамуса. Главным стимулом секреции окситоцина в кровь из задней доли гипофиза является раздражение сосков молочных желез при кормлении грудью. Другим фактором выброса гормона в кровь является расширение шейки матки и оргазм. Биологическое действие окситоцина заключается:1) в сокращении миоэпителиальных клеток в молочных железах.
БИЛЕТ
Антидиуретический гормон (вазопрессин) главным образом синтезируется в супраоптическом ядре гипоталамуса. Главным стимулом для его секреции является повышение осмолярности плазмы крови. Другими стимулами секреции антидиуретического гормона являются гиповолемия, гипотензия, боль, стресс и гипертермия. Ингибиторами выделения гормона являются снижение осмолярности плазмы крови, этанол, гипотермия. Основное биологическое действие гормона заключается в повышении реабсорбции воды в дистальных канальцах и собирательных трубочках почек путем активации мембрановстроенных белковых водных каналов – аквапоринов. Активация аквапоринов реализуется через связывание с поверхностными V2-рецепторами с образованием цАМФ. Менее значимое действие гормона в физиологических условиях – сокращение гладкой мускулатуры сосудов кожи и мышц обусловлено активацией V1-рецепторов и запуском фосфатидилинозитолового механизма. Вазопрессин обладает и метаболическим действием: 1) активирует распад гликогена; 2) стимулирует превращение глюкозы до ацетил-КоА; 3) активирует синтез жирных кислот. Кроме того, вазопрессин обладает мощным антикетогенным действием. Дефицит антидиуретического гормона приводит к развитию несахарного диабета (diabetus insipidus), характерными признаками которого являются:
– постоянная жажда;
– обильное (до 20 л/сут) мочеиспускание;
– низкая плотность мочи (1,001-1,005);
– отсутствие в моче глюкозы.
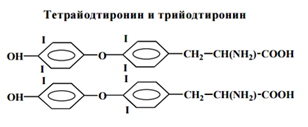
Синтез и секреция
Необходимой составной частью молекулы тиреоидных гормонов является йод. Он поступает с пищей и водой в виде йодидов. Суточная потребность в йоде – 150 мкг (1,2 мкМ).
Синтез тиреоидных гормонов происходит в фолликулах щитовидной железы. Этапы синтеза:
1. Йодид захватывается щитовидной железой из крови с помощью мембранного йодидного насоса.
2. При участии тиреоидпероксидазы йодид окисляется в ион иодиния (J+).
3. Ион йодиния атакует аминокислотные остатки тирозина в белке тиреоглобулина, который составляет основную массу коллоида фолликулов. Образуются моно- и дийодтирозилы. Указанная реакция называетсяорганификацией йода.
4. Моно- и дийодтирозилы конденсируются и образуются три- и тетрайодтиронилы.
5. Йодированные молекулы тиреоглобулина из коллоида пиноцитозом поступают в тиреоциты. Там от них в лизосомах отщепляются Т3 и Т4, которые секретируются в кровоток.
Щитовидная железа (ЩЖ) синтезирует и секретирует в кровь преимущественно тироксин (Т4).
Регуляция секреции
Непосредственным регулятором синтеза и секреции тиреоидных гормонов является тиреотропный гормон (ТТГ). Он оказывает стимулирующее действие на все 5 этапов синтеза тиреоидных гормонов. Кроме того, ТТГ усиливает синтез тиреоглобулина и рост фолликулов щитовидной железы.
Транспорт
В плазме крови 80% Т4 связано с тироксинсвязывающим глобулином (гликопротеин, синтезируемый в печени, с М.м. 54 кДа); 15% с тироксинсвязывающим преальбумином. Остальное количество связано с альбуминами и только 0,03% гормона остаются свободными. Т3 обладает меньшим сродством к транспортным белкам и его свободная форма составляет 0,3%. Время полужизни Т3 и Т4 в плазме крови составляет 1,5 суток и 7 суток, соответственно.
Периферический метаболизм (конверсия) тироксина
Около 80% Т3 образуются в результате периферической конверсии Т4 и только 20% циркулирующего Т3 секретируется тироцитами. Превращение Т4 в Т3происходит в периферических тканях при действии ферментов дейодиназ с образованием активного Т3 (3,5,3´-трийодтиронин) и неактивного реверсивного (обратного) Т3 (рТ3, 3,3´,5´-трийодтиронин). В физиологических условиях образования примерно равные количества активной и неактивной формы Т3. При тяжелых заболеваниях и голодании преобладает образование рТ3.
Механизм действия
По механизму действия тиреоидные гормоны относятся к гормонам, проникающим в клетку и действующим через внутриклеточные рецепторы. Рецепторы тиреоидных гормонов обнаружены практически во всех тканях и органах млекопитающих. Только гонады и лимфатическая ткань имеют мало рецепторов к тиреоидным гормонам. Рецепторы тиреоидных гормонов принадлежат к суперсемейству стероид-тиреоидгормональных рецепторов, то есть общий план их строения и механизма действия схожи. Однако рецепторы тиреоидных гормонов отличаются от рецепторов стероидных гормонов, тем, что они все время связаны с ДНК. В отсутствие тиреоидных гормонов они ингибируют экспрессию генов, с которыми они связаны. Связывание с гормоном превращает рецептор в активатор транскрипции. Ядерные рецепторы связываются преимущественно с Т3. Данный факт, а также существование механизма клеточной конверсии Т4 в Т3, позволяют рассматривать Т4 как прогормон, а Т3 – как истинный гормон. Однако, и сам тироксин способен давать ряд эффектов, обладая, по-видимому, собственными рецепторами на некоторых клетках-мишенях.
Биологические эффекты
1.Рост.
а) достижение соответствующего возрасту роста требует присутствия в адекватных количествах тиреоидных гормонов;
б) тиреоидные гормоны действуют синергично с гормоном роста и соматомединами, способствуя образованию костной ткани.
2. Центральная нервная система (ЦНС).
а) созревание ЦНС в перинатальный период абсолютно зависит от тиреоидных гормонов;
б) при дефиците тиреоидных гормонов у детей нарушаются процессы миелинизации, синаптогенеза и дифференцировки нервных клеток, что обуславливает выраженное замедление умственного развития. Если больному ребенку не давать тироксин, ментальные изменения необратимы.
3. Основной обмен (лабораторный показатель, отражающий энерготраты человека, находящегося в расслабленном состоянии утром вскоре после пробуждения).
а) тиреоидные гормоны увеличивают основной обмен и потребление кислорода всеми тканями, за исключением головного мозга, лимфоузлов и половых желез;
б) увеличение теплопродукции лежит в основе регуляции тиреоидными гормонами температуры тела;
в) согласно концепции Edelman и Ismail-Beigi (1970) тиреоидные гормоны увеличивают активность и синтез Na+/К+-АТФ-азы, для работы которой требуется значительное количество клеточной АТФ. Этим механизмом объясняется их способность повышать основной обмен.
4. Сердечно-сосудистая система.
– Тиреоидные гормоны повышают частоту и силу сердечных сокращений (за счет повышения синтеза β-адренорецепторов и их сродства к катехоламинам – пермиссивное действие тиреоидных гормонов на эффекты катехоламинов).
5. Метаболические эффекты:
а) незначительное гипергликемическое действие, обусловленное стимуляцией глюконеогенеза, мобилизацией гликогена и повышенным всасыванием глюкозы в желудочно-кишечном тракте;
б) стимуляция тканевого липолиза посредством повышения активности гормон-чувствительной липазы;
в) повышение синтеза белков, однако, в условиях избытка гормонов – протеокатаболическое действие;
г) повышение активности ЛПНП-рецепторов печени, участвующих в захвате ЛПНП из крови;
д) активация синтеза холестерина в печени за счет стимуляции ключевого фермента синтеза - ГМГ-КоА-редуктазы, но с одновременным усилением окисления холестерина в желчные кислоты. Итоговый результат двух последних эффектов – выраженное гипохолестеринемическое действие тиреоидных гормонов.

