




Синтез жирных кислот происходит в абсорбтивном периоде при высокой концентрации глюкозы в крови, протекает в цитоплазме на мембранах эндоплазматического ретикулума в печени и жировой ткани. В этот период активируются гликолиз и пентозофосфатный путь распада глюкозы, в результате которых образуются субстраты для синтеза жирных кислот: ацетил -КоА, NADPH2, АТР.
Образование ацетил-КоА в результате окислительного декарбоксилирования пирувата происходит в матриксе митохондрий, но ацетил-КоА не попадает через мембрану митохондрий в цитоплазму, где идет синтез жирных кислот.Поэтому ацетил-КоА конденсируется с оксалоацетатом с образованием цитрата и цитрат с помощью транслоказы переносится в цитоплазму (рис.12).
В цитоплазме под действием фермента цитратлиазы идет реакция:
Цитрат + HSKoA + АТР ® Ацетил-КоА + ADP + Рi + Оксалоацетат
Ацетил -КоА, перенесенный в цитоплазму, является исходным субстратом для синтеза жирных кислот. Оксалоацетат в цитозоле подвергается следующим превращениям: Оксалоацетат ® малат ® пируват. Последняя реакция катализируется малик – ферментом.
Пируват переносится обратно в матрикс митохондрий, a NADPH2, восстановленный в результате действия малик-фермента, используется как донор водорода для последующего синтеза жирных кислот. Другой источник NADPH2 — пентозофосфатный путь.
Первая реакция синтеза жирных кислот — это превращение ацетил-КоА в малонил-КоА (рис.13).
Фермент, катализирующий эту реакцию, ацетил-КоА-карбоксилаза, является регуляторным в биосинтезе жирных кислот. Он относится к классу лигаз, кофермент — биотин.
Последующие реакции синтеза жирных кислот происходят на ферменте, который называется синтазой жирных кислот или пальмитатсинтазой, так как основной жирной кислотой в липидах человека является пальмитиновая кислота. Некоторые жирные кислоты могут образовываться в организме из пальмитиновой кислоты.
Рисунок 11
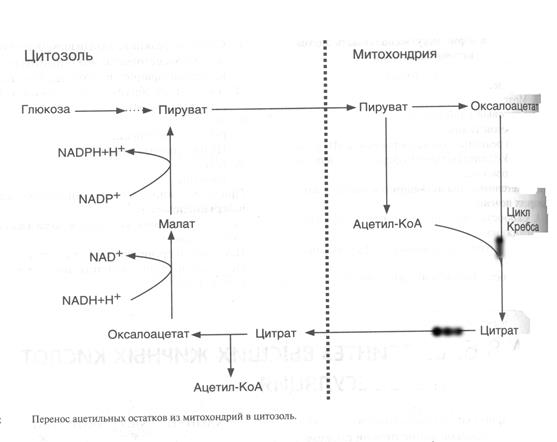
Рис. 12.
Синтаза жирных кислот (фермент фиксирован на мембранах эндоплазматического ретикулума). Синтаза жирных кислот является полифункциональным ферментом, состоящим из 2 идентичных полипептидных цепей, каждая из которых имеет 7 активных центров и ацилпереносящий белок, который переносит растущую цепь жирной кислоты из одного активного центра в другой. Каждый из белков имеет 2 центра связывания, содержащих SH-группы. Участок, связывающий малонил-КоА, содержит небелковый компонент – витамин B3 (пантотеновую кислоту).
Синтез жирной кислоты начинается с переноса ацетильного остатка, а затем малонильного с помощью ферментов ацетилтрансферазы и малонилтранс-феразы на синтазу жирных кислот. Далее карбоксильная группа малонила выделяется в виде СО2 и по освободившейся валентности присоединяется ацетил с образованием ацетоацетил-Е. Последующие реакции восстановления, дегидратации, восстановления приводят к образованию радикала бутирила, связанного с ферментом. Затем повторяется такой же цикл реакций и образуется радикал жирной кислоты с 6 углеродными атомами. Цикла повторяются вплоть до образования радикала пальмитиновой кислоты. Фермент тиоэстераза отщепляет от фермента жирную кислоту (рис.13).
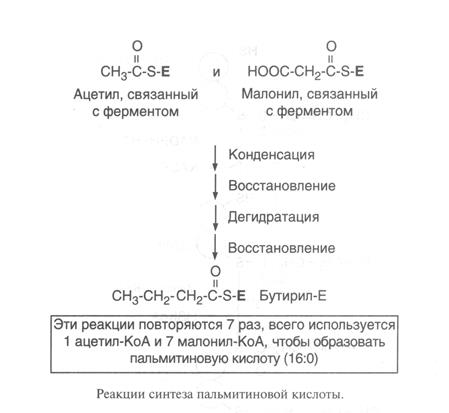
Рисунок 13
Биосинтез жирных кислот является процессом, в котором повторяются одни и те же последовательности реакций, поэтому процесс называется циклическим, и в каждом цикле радикал жирной кислоты увеличивается на 2 атома углерода, источником которых является малонил-КоА. В каждом цикле происходят реакции восстановлении с использованием NADPH+Н+ одним из источников которого является пентозофосфатный пуп окисления глюкозы, другим — малик-фермент.
Другие жирные кислоты в организме человека синтезируются из пальмитиновой кислоты, при эта происходят реакции удлинения углеродного скелета также с использованием малонил-КоА и реакции дегидрирования.
В организме человека не синтезируются жирные кислоты с двойными связями, расположенными дистальнее С9-го атома углерода, поэтому человек должен получать незаменимые жирные кислоты с пищей.
Синтезированные жирные кислоты не остаются |в свободном виде, а быстро используются для синтеза жиров и в меньшей степени фосфолипидов.

