




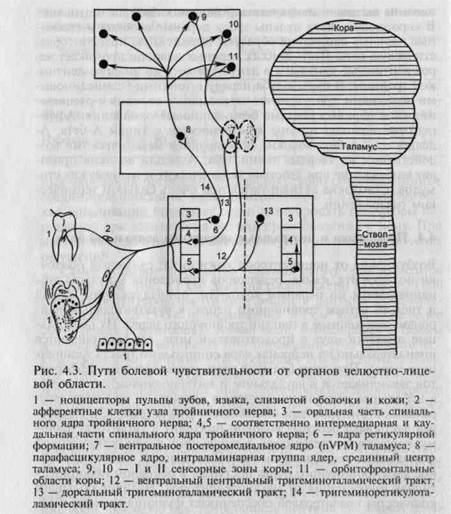
Возбуждение от ноцицепторов кожи лица, слизистой оболочки полости рта, языка, рецепторов периодонта и пульпы зуба направляется по нервным волокнам, принадлежащим второй и третьей ветвям тройничного нерва, к чувствительным нейронам, заложенным в ганглии тройничного нерва. Их центральные отростки идут в продолговатый мозг, где заканчиваются ипсилатерально на нейронах ядра спинального тракта тройничного нерва. При этом большая часть ноцицептивных афферен-тов заканчивается в каудальном и интерполярном его отделах, а небольшая часть ноцицептивных волокон — на контралате-ральных структурах ядра спинального тракта (рис. 4.3). Некоторая часть афферентных волокон приходит к гигантоклеточно-му, парагигантоклеточному, латеральному ядрам ретикулярной формации, а также к ядрам шва. В переднее и главное сенсорное ядра тройничного нерва поступает в основном неноцицеп-тивная информация от механорецепторов. Наличие большого количества коллатералей обеспечивает функциональную взаимосвязь между различными ядрами тройничного нерва (триге-минальный комплекс ядер), что играет существенную роль во взаимодействии возбуждений ноцицептивного и неноцицептив-ного характера.
Нейроны ядер тройничного комплекса дают начало нескольким восходящим трактам. Тригеминоталамические проекции образуют четыре тракта. Два из них — контралатеральный три-геминальный лемнисковый («тригеминальный лемниск») и ипсилатеральный тригеминальный тракты — проводят возбуждение, вызванное неноцицептивной стимуляцией тактильных рецепторов различных структур челюстно-лицевой области. Вентральный центральный и дорсальный тригеминоталамичес-
|
кий тракты образованы аксонами нейронов I, III—V слоев каудального и интерполярного отделов ядра спинального тракта тройничного нерва. Большинство волокон этих трактов заканчивается на нейронах заднего вентромедиального ядра таламу-са. Эти проекции организованы по соматотопическому принципу. Часть клеток этого ядра являются специфическими но-цицептивными, отвечающими только на один вид ноцицеп-тивной стимуляции, другие — нейронами широкого динамического ряда, отвечающими на стимуляцию механо-, термо-и хемоноцицепторов.
Существенную роль в формировании прозопалгий играет тригемино-ретикуло-таламический тракт, передающий возбуждения от пульпы зубов и ноцицепторов других структур челю-стно-лицевой области — через ядра ретикулярной формации к неспецифическим ядрам таламуса (парафасцикулярное, центральное латеральное ядра, срединный центр, интраламинар-ная группа). Эта группа ядер содержит в основном полимодальные нейроны, отвечающие на разнообразную сенсорную стимуляцию. Вместе с тем в ней обнаружено небольшое количество нейронов, реагирующих исключительно на болевые воздействия. Включение ретикулярной формации, специфических и неспецифических ядер таламуса в процесс передачи ноци-цептивной информации от органов челюстно-лицевой области определяет ее поступление к сенсорным зонам коры, к ее орбитофронтальной области, а также широкую генерализацию ноцицептивных возбуждений в структурах лимбико-гипотала-мической области.
В сенсорных зонах коры большого мозга имеется топическая организация представительства структур челюстно-лицевой области, в частности различных зубов. Клетки коры, отвечающие на раздражение пульпы зубов, разделены на две группы. Нейроны первой группы — F (fast — быстрый), отвечают на стимуляцию пульпы первого и второго зубов с коротким латентным периодом. Информация к ним приходит через тригеми-но-таламические тракты, заканчивающиеся на нейронах заднего вентромедиального ядра, образующего прямые проекции в «оральную» сенсорную зону коры. Нейроны второй группы — S (slow — медленный) отвечают на стимуляцию четвертого— восьмого зубов с большим латентным периодом. Эти нейроны активируются через тригемино-ретикуло-таламические пути, оканчивающиеся в неспецифических ядрах таламуса, дающих обширные таламо-кортикальные проекции.
Считают, что сенсорная зона I образует сенсорно-дискрими-нативную систему, которая определяет качество, пространственную локализацию, интенсивность дентальной боли, регулирует двигательные акты, возникающие при болевом воздействии, формирует ощущение первичной эпикритической боли. Сенсорная зона II получает информацию не только от специфических ядер зрительного бугра, но в значительной степени от группы неспецифических его ядер. Эта зона ответственна за восприятие боли как возбуждения сенсорной модальности, оценку потенциально опасных воздействий и формирование адекватных защитных реакций, включение механизмов антиноцицептивной системы.
Орбитально-фронтальная область коры участвует в формировании сложных эмоционально-аффективных проявлений боли

|
и связанных с ней психических переживаний, особенно выраженных при повреждении структур челюстно-лицевой области. Генерализация ноцицептивных возбуждений через внутрипла-стинчатые ядра таламуса обеспечивает вовлечение лимбичес-ких структур в процесс переработки поступающей ноцицеп-тивной информации и формирование вегетативного «портрета», мотивационного иэмоционального компонентов болевой реакции, а также пролонгирование субъективных переживаний и адаптивных реакций. Результатом прихода возбуждений в центральные отделы мозга является формирование ощущения боли с более или менее выраженными поведенческими, эмоционально-аффективными и вегетативными реакциями, направленными на сохранение целостности тканей челюстно-лицевой области.
Для дентальной боли, особенно интенсивной, характерна конвергенция на нейронах коры болевых сигналов от афферен-тов пульпы различных зубов и окружающих тканей, что обусловливает широкую иррадиацию возбуждения, затрудняющую локализацию боли. Дентальные боли в ряде случаев могут проецироваться не только в область развития патологического процесса (например, к пораженному зубу или участку паро-донта), но и в достаточно удаленные участки лица, головы и шеи (отраженные боли). В основе появления проекционных зон лежит тесное взаимодействие ноцицептивных и неноцинептив-ных нейронов различных ядер тригеминального комплекса, обусловленное наличием богатых связей между ними, а также с ядрами ретикулярной формации. Существенную роль играет также более тонко организованное представительство кожи лица в коре большого мозга, занимающее значительную область сенсорной зоны. Это создает возможность конвергенции ноцицептивных и неноцицептивных возбуждений на нейронах коры, обеспечивающих чувствительность кожи определенных зон лица, головы и шеи с проекцией боли именно к этим участкам (табл. 4.2). В ряде случаев после операции удаления зуба могут развиваться фантомные боли — ощущение боли в удаленном зубе или области его фиксации. Фантомные боли рассматривают как деафферентационные. Удаление зуба приводит к повышению возбудимости с одновременным дефицитом тормозных процессов клеток разных уровней ЦНС, обеспечивающих чувствительность данного зуба. Предшествующая операции более или менее длительная ноцицептивная афферентация из области пораженного зуба создает определенную готовность нервных структур к длительной циркуляции возбуждений. Дополнительная афферентация при операции удаления зуба «запускает» циркуляцию, создавая генератор патологически усиленных возбуждений, воспринимаемых клетками коры мозга как
длительные, часто непрерывные боли. Вовлечение в процесс циркуляции патологически усиленных возбуждений нескольких структур мозга приводит к формированию патологической алгической системы. При фантомных болях лечебные мероприятия местного характера не приводят к исчезновению или снижению болей, так как их источник лежит в структурах мозга, на которые иследует воздействовать, усиливая работу тормозных механизмов.

