




Человек может воспринимать звук в диапазоне частот от 16 до 20000 Гц. Звуковые колебания разной высоты вовлекают в колебательный процесс перелимфу вестибулярного канала улитки и соответственно базилярную мембрану на всем их протяжении неодинаково. Так, колебательная реакция базилярной мембраны, передаваемая на эндолимфу, имеет характер бегущей волны, а локализация амплитудного максимума этой волны зависит от высоты звука. При действии на ухо низких и средних частот в колебательный процесс приходит вся базилярная мембрана, что сопровождается вовлечением в возбуждение волосковых клеток на всем протяжении кортиева органа, а частота звуковых колебаний при этом кодируется числом импульсов, возникающих в волокнах слухового нерва, которое полностью соответствует частоте звукового сигнала (временное кодирование информации). При действии же на ухо высоких частот амплитудный максимум звуковой волны приходится на начальные отделы базилярной мембраны (расположенные ближе к основанию улитки), причем, чем выше звуковой тон, тем меньшая площадь базилярной мембраны у основания улитки вовлекается в колебательный процесс. Следовательно, при действии высоких частот происходит пространственное кодирование информации, основанное на неодинаковом расположении возбужденных рецепторных клеток на базилярной мембране и соответственно избирательным вовлечением в возбуждение определенных групп слуховых нейронов.
Сила звуков кодируется числом возбужденных рецепторных клеток и самих слуховых нейронов, а также частотой их импульсации. Волосковые рецепторные клетки, также как и слуховые нейроны, резко отличаются друг от друга по своей чувствительности. Так, наружные волосковые клетки гораздо более чувствительны, чем внутренние. В связи с этим при действии на ухо слабых звуков в реакцию вовлекается лишь небольшое число наиболее чувствительных рецепторных клеток и слуховых нейронов, а при усилении стимулов возбуждается все большее количество дополнительных рецепторных и нервных клеток кортиева органа.
Минимальная сила звука, слышимого человеком в половине случаев его предъявления, носит название абсолютной слуховой чувствительности. Абсолютная слуховая чувствительность в разном диапазоне частот звуковых сигналов не является одинаковой. В частности, она максимальна в области частот от 1000 до 4000 Гц (соответствуют речевому диапазону), а при слишком низких (16-50 Гц) либо слишком высоких частотах (10000-20000 Гц) является минимальной.
Адаптация слухового анализатора
Слуховой анализатор, подобно многим другим обладает способностью к адаптации, которая возникает при продолжительном действии на ухо звука постоянной интенсивности и частоты. Причем слуховой анализатор относится к быстро адаптирующимся анализаторам (скорость адаптации обычно составляет десятки секунд). Первоочередную роль в адаптации слухового анализатора имеют тормозные эфферентные влияния ретикулярной формации как на структуры периферического отдела (собственно рецепторные клетки и слуховые нейроны), так и на нейроны кохлеарных ядер ромбовидной ямки и саму слуховую кору.
Бинауральный слух
Человек, другие млекопитающие животные и птицы обладают пространственным слухом – способностью определять положение источника звука в пространстве. Это свойство основано на наличии в слуховом анализаторе двух симметричных не перекрещивающихся половин (бинауральный слух), а также способности определенных нейронов оценивать межушные различия звуковых стимулов по времени их прихода на каждое ухо и по их интенсивности. Острота бинаурального слуха у человека очень высока: он способен определять расположение источника звука с точностью порядка 1 углового градуса.
ВЕСТИБУЛЯРНЫЙ АНАЛИЗАТОР
Вестибулярный анализатор специализируется на восприятии изменений положения тела (прежде всего головы) в пространстве, а также действия на организм ускорений при движении и играет важную роль, наряду со зрительным и проприоцептивным анализаторами, в пространственной ориентировке человека. В условиях покоя или равномерного движения рецепторы вестибулярного анализатора не возбуждаются.
Периферическая (рецепторная) часть вестибулярного анализатора находится во внутреннем ухе (перепончатый лабиринт преддверия и полукружных каналов) и представлена вестибулярными волосковыми клетками, вступающими в синаптический контакт с дендритами вестибулярных нейронов, тела которых находятся в вестибулярном ганглии (заложен в толще пирамидки височной кости). Аксоны вестибулярных нейронов образуют преддверную (вестибулярную) ветвь преддверно-улиткового нерва (VIIIп) и направляются к вестибулярным ядрам, локализованным в латеральных углах ромбовидной ямки (латеральное вестибулярное ядро (ядро Дейтерса), медиальное вестибулярное ядро (ядро Швальбе ), верхнее вестибулярное ядро (ядро Бехтерева) и нижнее вестибулярное ядро (ядро Ролкера)). Нейроны вестибулярных ядер, в свою очередь, дают проекции к
Ø спинному мозгу (от латерального и нижнего вестибулярных ядер начинается вестибуло-спинальный путь),
Ø ретикулярной формации ствола головного мозга,
Ø мозжечку,
Ø красному ядру и черной субстанции среднего мозга,
Ø вегетативным ядрам,
Ø а также к нейронам латеральной группы ядер таламических бугров, посылающих свои аксоны к коре больших полушарий (задняя часть постцентральной извилины и нижняя часть моторной зоны, локализованной в предцентральной извилине и центральной борозде).
Связь нейронов вестибулярных ядер со спинным мозгом и со структурами ствола головного мозга обеспечивает возможность быстрого (неосознанного) перераспределения тонуса определенных групп скелетных мышц, а также изменения деятельности некоторых внутренних органов в ответ на возбуждение ветибулорецепторов. Высший же анализ информации, поступающей от вестибулорецепторов, происходит на уровне коры больших полушарий и заканчивается формированием у человека представления о положении своего тела и головы в пространстве.
Вестибулорецепторы заложены в перепончатом лабиринте преддверия (маточке и мешочке) и ампул (начальных расширений) полукружных каналов внутреннего уха, формирующих в своей совокупности вестибулярный аппарат. В маточке и мешочке сенсорные образования имеют форму пятна (макулы), над которым нависает желатинозная отолитовая мембрана, включающая кристаллы бикарбоната кальция (удельный ее вес в 3 раза больше такового эндолимфы, заполняющей полость перепончатого лабиринта внутреннего уха). В ампулообразных расширениях перепончатых полукружных каналов рецепторные структуры вестибулярного анализатора имеют вид поперечного гребня (кристы), над поверхностью которого расположена желатинозная мембрана – купула, лишенная включений бикарбоната кальция (в связи с чем ее удельный вес такой же, как и эндолимфы, заполняющей перепончатый лабиринт полукружных каналов). Причем если отолитовая мембрана лежит над пятнами маточки и мешочка наподобие плоского камня над пластиной, то купула, подобно створчатому клапану или шарнирной дверце, идет от гребешка прямо поперек ампулы, замыкая ее просвет.

Рис. 13. Вестибулярный аппарат (поперечный разрез через пятно эллиптического мешочка и ампулу полукружного канала)
Макулы (пятна) маточки и мешочка преддверия и гребни (кристы) ампул перепончатых полукружных каналов состоят из утолщенного эпителия, включающего волосковые (рецепторные) и поддерживающие (опорные) клетки. Волосковые клетки по форме могут быть цилиндрическими и колбовидными. На апикальной поверхности этих клеток расположены стереоцилии, склеенные между собойи неподвижные (60-80 штук, производные мембраны, называемые также волосками или микроворсинками) и, как правило, одна ресничка – киноцилия (длинная и подвижная). Базальная поверхность волосковых клеток принимает участие в образовании синаптического контакта с дендритами вестибулярных нейронов.
Волосковые клетки макул перепончатого лабиринта преддверия воспринимают действие силы тяжести и линейных ускорений на организм. Если голова, а следовательно, и макула наклонены относительно силы тяжести, то отолитовая мембрана также стремится скользнуть наклонно, при этом тянет волоски волосковых клеток, деформация которых приводит к возбуждению волосковых клеток. Волосковые клетки гребней ампул перепончатых полукружных каналов выявляют изменения углового ускорения. Когда голове придано угловое ускорение, эндолимфа в определенном полукружном канале стремится сохранить неподвижность в силу инерции, а перепончатые стенки движутся относительно нее. Это движение смещает купулу из ее обычного положения относительно гребешка, смещенная же купула тянет волоски волосковых клеток, деформация которых приводит к возбуждению волосковых клеток (генерации рецепторного потенциала). Возбуждение волосковых клеток, в свою очередь, сопровождается выделением в синаптическую щель из их пресинаптических окончаний медиатора. Медиатор же, достигая постисинаптической мембраны (участка мембраны дендрита вестибулярного нейрона), вызывает ее деполяризацию (возникновение постсинаптического потенциала), порождающую нервный импульс в соседних участках мембраны вестибулярного нейрона. Следовательно, вестибулярные рецепторы представляют собой в функциональном плане вторичночувствующие механорецепторы.
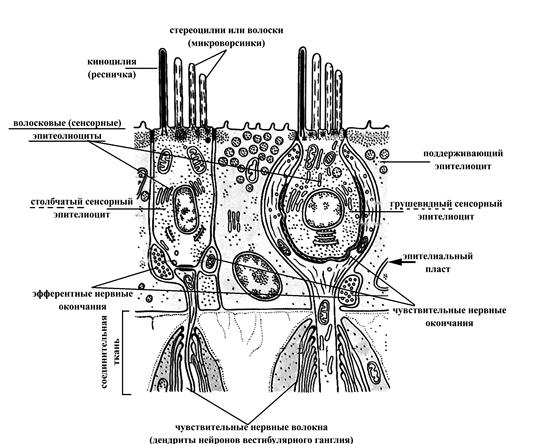
Рис. 14. Схема ультрамикроскопического строения вестибулярных рецепторов
Чувствительность вестибулярного анализатора здорового человека очень высока. Так, отолитовый аппарат позволяет воспринимать наклон головы в сторону всего на 1º, а рецепторная система полу кружных каналов – замечать ускорения вращения всего в 2-3 º/с2.
КОЖНЫЙ АНАЛИЗАТОР
Кожный анализатор специализируется на восприятии воздействий, оказываемых на кожу человека, и характеризуется очень широким рецептивным полем (от 1,4 до 2,1м2). Чувствительная информация от кожи туловища и конечностей проводится нейронами спинальных ганглиев в спинной мозг, а от кожи головы и шеи – нейронами полулунного узла тройничного нерва (Vп) в головной мозг (к чувствительным ядрам этого нерва, заложенным в ромбовидной ямке). Аксоны нейронов спинальных ганглиев, доставляющие информацию от болевых и температурных рецепторов кожи туловища и конечностей, переключаются на нейроны собственного ядра задних рогов спинного мозга, тогда как информация от тактильных рецепторов кожи туловища и конечностей проводится нейронами спинальных ганглиев к желатинозной субстанции Ролланда. Аксоны нейронов собственного ядра задних рогов спинного мозга и желатинозной субстанции Ролланда восходят к головному мозгу, формируя соответственно латеральный и вентральный спинно-таламические тракты, которые перекрещиваются еще на уровне спинного мозга. Нервные волокна этих двух трактов входят в головной мозг и вместе с аксонами нейронов чувствительных ядер тройничного нерва, проводящих информацию от рецепторов кожи головы и шеи, следуют в мозговом стволе в составе медиальной петли и достигают таламических бугров (латеральной специфической группы ядер и вентральной неспецифической группы ядер). Аксоны же нейронов специфических ядер таламуса, имеющих отношение к обработке кожной информации, восходят к корковому представительству кожного анализатора, заложенному в теменной доле коры (постцентральная извилина и верхняя теменная долька).
Рецепторная часть кожной сенсорной системы заложена в толще кожи: в эпителиальном пласте, сосочковом и сетчатом слоях дермы, подкожной жировой клетчатке. В функциональном плане чувствительные нервные окончания кожи можно классифицировать на три группы:
Ø тактильные (воспринимают прикосновение, давление, вибрацию)
Ø температурные
ü холодовые (в среднем насчитывается в коже 250тыс., заложены более поверхностно по сравнению с тепловыми (на глубине 0,17мм), максимально возбуждаются при температурах от 10 до 41ºС)
ü тепловые (в среднем насчитывается в коже 30 тыс., заложены более глубоко по сравнению с холодовыми (на глубине 0,3мм), максимально возбуждаются при температурах от 20 до 50ºС)
Ø болевые (ноцицепторы).
 В морфологическом плане рецепторы кожи отличаются большим разнообразием, локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. Кожные рецепторы могут быть представлены следующими морфологическими типами:
В морфологическом плане рецепторы кожи отличаются большим разнообразием, локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. Кожные рецепторы могут быть представлены следующими морфологическими типами:
Ø Свободные нервные окончания (разветвления дендритов чувствительных нейронов)
Типичны для эпителиального пласта кожи и роговицы глаза, где могут выступать в качестве механорецепторов (тактильные рецепторы и ноцицепторы) и терморецепторов.
Ø Свободные нервные окончания, оплетающие волосяные луковицы
 |
Типичны для соединительной ткани кожи, имеющей волосяной покров, выступают в качестве механорецепторов (тактильных рецепторов, реагирующих на прикосновение и давление ).

Oslash; Диски Меркеля
Встречаются только в эпителии кожи, причем, как правило, не имеющей волосяного покрова; залегают в глубоких слоях эпителиального пласта (на границе его с соединительной тканью). Морфологически представлены расширенной терминалью дендрита чувствительного нейрона, прилегающей к клетке Меркеля (крупная отросчатая клетка, залегающая в базальном слое эпителиального пласта). Существуют различные точки зрения относительно происхождения клеток Меркеля. Одни исследователи считают их видоизмененными эпителиальными клетками, другие – производными нервных клеток, мигрирующих на ранних этапах эмбриогенеза из нервной трубки. В пользу последнего предположения свидетельствует факт наличия в цитоплазме этих клеток гранул с нейроноспецифическими пептидами. Диски Меркеля представляют собой первичночувствующие рецепторы, поскольку клетка Меркеля не является рецепторной и, по-видимому, несет позиционную информацию для определения места окончательной локализации терминали дендрита чувствительного нейрона в нейрогенезе или при регенерации нервов. В функциональном плане диски Меркеля являются механорецепторами, реагирующими на прикосновение и давление (тактильные рецепторы кожи).
Ø Несвободные нервные окончания, лишенные соединительнотканной капсулы

Типичны для соединительной ткани (как правило, рыхлой волокнистой) кожи, где разветвляются параллельно ее поверхности. В функциональном плане могут выступать в качестве механорецепторов (некоторые тактильные рецепторы кожи), терморецепторов и ноцицепторов.
Ø Инкапсулированные нервные окончания (имеют собственную соединительно тканную капсулу)
Колбы Краузе
 Морфологически представляет собой разветвление дендрита чувствительного нейрона, сопровождаемое глиальными клетками и покрытое тонкой собственной соединительнотканной капсулой. Причем чувствительное нервное волокно перед проникновением под капсулу колбы Краузе утрачивает миелиновую оболочку. Типичны для соединительной ткани кожных покровов наружных половых органов
Морфологически представляет собой разветвление дендрита чувствительного нейрона, сопровождаемое глиальными клетками и покрытое тонкой собственной соединительнотканной капсулой. Причем чувствительное нервное волокно перед проникновением под капсулу колбы Краузе утрачивает миелиновую оболочку. Типичны для соединительной ткани кожных покровов наружных половых органов
В функциональном плане, вероятнее всего, являются первичночувствующимимеханорецепторами.
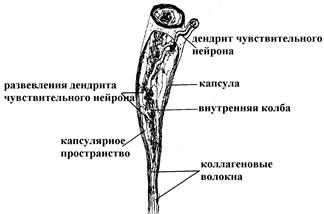
Тельца Руффини
Имеют удлиненную форму, длина тельца составляет 1-2 мм, ширина – 0,1-0,15 мм. Тельце Руффини состоит из внутренней колбы, окружающей ее базальной мембраны, за которой располагается капсулярное пространство, покрытое соединительнотканной капсулой. Внутренняя колба формирует сердцевину тельца и представлена булавовидно заканчивающимися разветвлениями чувствительного нервного волокна, окруженными пластинчатыми клетками (глиальной природы). Капсулярное пространство заполнено жидкостью, содержит фибробласты, макрофаги и неориентированные коллагеновые волокна, вплетающиеся во внутреннюю колбу. Соединительнотканная капсула тельца Руффини состоит из 4-5 слоев уплощенных фиброцитов. Тельца Руффини типичны для соединительной ткани кожи, особенно, подошвенной поверхности стопы (залегают в сетчатом слое дермы и подкожной клетчатке). В функциональном плане, вероятнее всего, являются первичночувствующими механорецепторами.
Тельца Мейснера
 |

А Б
Рис. 15. Тельца Мейснера (А – общий вид, Б – схема, отражающая взаимоотношение тельца с эпителиальным пластом)
Имеют овальную форму, длина – 100мкм, ширина – 50мкм. Типичны для соединительной ткани кожи, особенно не имеющей волосяного покрова (пальцы, ладонная поверхность кисти и подошвенная поверхность стопы, губы, веки, наружные половые органы, соски молочных желез). Залегают поверхностно (на границе сосочкового слоя дермы и эпителиального пласта), причем своей длинной осью ориентированы перпендикулярно поверхности кожи. Покрыты соединительнотканной капсулой, под которой залегают уплощенные швановские клетки. Дендрит чувствительного нейрона после проникновения внутрь тельца Мейснера утрачивает миелиновую оболочку и образует спиралевидные разветвления, проходящие между шванновскими клетками. Ядра последних оттеснены на периферию тельца (ближе к капсуле), а их цитоплазма образует пальцевидные выросты, направленные к его центру; через посредство этих выростов глиальные клетки контактируют между собой. С помощью микрофибрилл и коллагеновых волокон шванновские клетки прикрепляются к капсуле тельца, а капсула – к базальной мембране эпителиального пласта. В результате отмеченного любое механическое воздействие на кожу приводит к деформации тельца Мейснера и возбуждению дендрита чувствительного нейрона, образующего разветвления между шванновскими клетками. Следовательно, тельца Мейснера являются первичночувствующими механорецепторами.
Тельца Фатера – Пачини
Тельца овальной формы, длина составляет 1-4мм, ширина – 0,5-1мм. Покрыты капсулой, включающей несколько слоев соединительно-тканных клеток, под которой расположена сердцевина, состоящая из наружной и внутренней колб. В центре сердцевины находится терминаль дендрита чувствительного нейрона с одним или несколькими булавовидными расширениями на конце. Подобно другим инкапсулированным нервным окончаниям, дендрит чувствительного нейрона после проникновения под капсулу тельца утрачивает миелиновую оболочку. Наружная и внутренняя колбы представлены концентрически расположенными слоями глиальных клеток (причем в наружной и внутренней колбах насчитывается соответственно до 60 и 80слоев глиальных клеток); между глиальными клетками находятся межклеточная жидкость и коллагеновые волокна.

Типичны для соединительной ткани кожи, особенно не имеющей волосяного покрова (пальцы, ладонная поверхность кисти и подошвенная поверхность стопы), а также кожи наружных половых органов и молочных желез. В коже залегают глубоко в дерме (в сетчатом слое, а также в подкожной клетчатке), причем своей продольной осью ориентированы перпендикулярно поверхности кожного покрова. В функциональном плане являются первичночувствующими механо-рецепторами. В коже тельца Фатера-Пачини воспринимают прикосновение и вибрацию.
Следовательно, большинство рецепторов кожи является первичночувствующими. Возбуждение первичночувсвтующих механорецепторов возникает следующим образом: механический стимул приводит к деформации мембраны рецептора и, как следствие, снижению ее входного сопротивления, что сопровождается увеличением проницаемости мембраны для ионов натрия. Входящий натриевый ток, в свою очередь, деполяризует терминальные участки дендрита чувствительного нейрона (в них возникает рецепторный потенциал), по достижении же критического уровня деполяризации в мембране соседних с деполяризованным участком частях дендрита генерируется нервный импульс.
Механизм возбуждения рецепторов при болевых воздействиях остается до конца не ясным. Предполагают, что непосредственными раздражителями болевых рецепторов выступают изменение рН ткани (особенно сдвиг в кислую сторону) в области нервного окончания, выделение из поврежденных клеток гистамина, пртеолитических ферментов, воздействующих на глобулины межклеточной жидкости и приводящих к образованию ряда полипептидов (например, брадикинина), которые могут возбуждать чувствительные нервные окончания. Наконец, ощущение боли может возникать в результате сильного механического сдавления чувствительных нервных окончаний возникшим в ткани отеком.

