




ФИЗИОЛОГИЯ СЕНСОРНЫХ СИСТЕМ
Анализатором или сенсорной системой называют часть нервной системы, специализирующуюся на восприятии определенных сигналов, проведении информации о них к коре больших полушарий и анализе этой информации. Анализаторы – это системы входа чувствительной информации в мозг и анализа этой информации. В качестве раздражителей, которые могут восприниматься определенными анализаторами, служат различные факторы как внешней, так и внутренней среды организма. При этом каждый анализатор оптимально настроен на восприятие каких-то определенных сигналов (называемых адекватными), к которым является эволюционно приспособленным. Так, адекватным раздражителем для зрительного анализатора являются световые волны (l=397-723нм), для слухового – звук, обонятельного и вкусового – молекулы химических веществ и т.д. К адекватным раздражителям анализаторы проявляют очень высокую чувствительность (воспринимают их при ничтожно малой энергии действия на рецептивное поле). В частности, для возбуждения фоторецептора достаточно попадания на него всего лишь одного кванта света в видимой части спектра, обонятельные рецепторы возбуждаются при контакте с единичными молекулами пахучих веществ. Все остальные раздражители (неадекватные) тоже могут восприниматься рецептивными полями анализаторов, но для этого они должны иметь гораздо большую силу, чем адекватные. К примеру, незначительного надавливания на глазное яблоко не достаточно для возбуждения фоторецепторов, тогда как сильное механическое воздействие на него приводит к возникновению у человека ощущения вспышки света.
Минимальная сила раздражителя, необходимая и достаточная для восприятия его анализатором, характеризует абсолютный порог чувствительности анализаторной системы.
В зависимости от того, на восприятие каких раздражителей (факторов внешней или внутренней среды) настроены анализаторы их классифицируют на:
Ø экстерорецептивные (зрительный, слуховой, вкусовой, обонятельный, кожный)
Ø интерорецептивные. (висцероцептивный, проприоцептивный, вестибулярный).
У млекопитающих животных и человека выделяют 8 анализаторов:
Ø зрительный
Ø слуховой
Ø кожный
Ø вестибулярный
Ø проприоцептивный
Ø висцероцептивный
Ø обонятельный
Ø вкусовой.
Деятельность большинства анализаторов (но не всех) заканчивается формированием какого-то ощущения (это объективный процесс), на основании которого у человека формируется определенное его восприятие, во многом зависящее от индивидуального опыта, функционального состояния организма, настроения и т.д. (это субъективный процесс). Так, человек может чувствовать какой-то запах и на основании этого ощущения его идентифицировать. Деятельность не всех анализаторов заканчивается формированием ощущения. В частности, мы в норме не чувствуем, что происходит в наших внутренних органах, как колеблется артериальное давление, рН крови, ее газовый состав и другие константы гомеостаза. Формирование ответной программы действий на воспринятый сигнал и ее передача к исполнительным органам в функции анализаторов не входят. Следовательно, анализаторы обеспечивают постоянную взаимосвязь животного организма с его внешней и внутренней средой, специализируясь только лишь на восприятии и анализе информации.
Несмотря на то, что все эти анализаторы отличаются друг от друга определенными морфо-функциональными особенностями, они имеют единый план строения. В частности, в состав любого анализатора входят рецепторы (специализированный воспринимающий аппарат), собственно чувствительные нейроны и различной сложности цепи из вставочных нейронов, а также совокупность корковых нейронов, обеспечивающих высший анализ информации. По И.П. Павлову в любом анализаторе выделяют три отдела:
рецепторный (периферический),
проводниковый (промежуточный),
центральный (корковый).

|
|
|
|
|
|
|
|
 Рис.1. Общий план строения анализаторов
Рис.1. Общий план строения анализаторов
Периферический (рецепторный) отдел представлен рецепторами, специализированными на обнаружении и различении сигналов. В качестве рецепторов с морфологической точки зрения могут выступать либо собственно окончания дендритов чувствительных нейронов (первичночувствующие рецепторы), либо в более сложном случае – комплекс специальной рецепторной клетки (как правило, не нервной природы) и дендрита чувствительного нейрона (вторичночувствующие рецепторы), связанных между собой посредством синаптического контакта. Рецепторы являются своеобразными датчиками, преобразующими энергию раздражителя (химическую, механическую, тепловую и т.д.) в энергию электрического ответа (т.е. в электрическую энергию). Действующий на рецепторы раздражитель, вызывает, как правило, изменение конформации макромолекул (в основном белков), входящих в состав мембран рецептора, что приводит к изменению их проницаемости для определенных ионов и, как следствие, величины мембранного потенциала (возникает т.н. рецепторный потенциал, по своей природе соответствующий локальному ответу). Рецепторный потенциал, возникающий в первичночувствующих рецепторах, вызывает генерацию потенциала действия (нервного импульса) в наиболее возбудимых участках этой же структуры. В случае вторичночувствующих рецепторов рецепторный потенциал, возникший в собственно рецепторной клетке, инициирует высвобождение из ее пресинаптического окончания квантов медиатора. Медиатор же, воздействуя на постсинаптическую мембрану (мембрану дендрита чувствительного нейрона), приводит к генерации в ней постсинаптичекого потенциала, который инициирует возникновение на соседних участках дендрита нервного импульса. Таким образом, рецепторы обеспечивают преобразование энергии раздражителей в энергию электрического ответа (в конечном итоге в энергию нервного импульса); дальнейшая передача информации в нервной системе осуществляется только в форме электрического ответа (т.е. декодирования информации в нервной системе не происходит).
Восприятие рецепторами сигналов предполагает их обнаружение и различение. Под обнаружением рецепторами сигналов понимают их способность возбуждаться при действии раздражителей определенной силы (не меньше пороговой). Различение рецепторами сигналов – это их способность дифференцировать сигналы, отличающиеся по своей интенсивности (различение сигналов по интенсивности), или одновременно действующие на соседние участки рецептивного поля (пространственное различение сигналов), либо воспринимать, как различные, сигналы, действующие последовательно на один и тот же участок рецептивного поля через определенный интервал времени (временное различение).
Существуют различные классификации рецепторов. Так, в зависимости от того, на восприятиераздражителей внешней или внутренней среды организма настроены чувствительные нервные окончания, их классифицируют на:
Ø Экстероцептивные ( воспринимают изменения во внешней среде организма: фоторецепторы сетчатки глаза, слуховые рецепторы внутреннего уха, обонятельные и вкусовые рецепторы, тактильные, болевые и температурные рецепторы кожи)
Ø Интероцептивные, воспринимают изменения во внутренней среде организма,представлены тремя группами:
ü висцерорецепторы (рецепторы внутренних органов: температурные и болевые рецепторы, рецепторы растяжения и давления, хеморецепторы)
ü проприорецепторы (рецепторы опорно-двигательного аппарата: мышечные веретена (рецепторы растяжения) скелетных мышц, тельца Фатера-Пачини (рецепторы давления) капсул суставов, рецепторы Гольджи (рецепторы растяжения) связок и сухожилий)
ü вестибулорецепторы (рецепторы вестибулярного аппарата, заложенного во внутреннем ухе)
В зависимости от природы раздражителя, на восприятие которого оптимально настроены рецепторы, их классифицируют на (функциональная классификация рецепторов):
Ø фоторецепторы (воспринимают световые волны в диапазоне у человека от 397 до 723 нм)
Ø 
 терморецепторы (локализованы в коже, сосудах, внутренних органах и самой центральной нервной системе) холодовые
терморецепторы (локализованы в коже, сосудах, внутренних органах и самой центральной нервной системе) холодовые
тепловые
Ø механорецепторы (тактильные рецепторы кожи, барорецепторы (прессорецепторы) сосудов, внутренних органов и капсул суставов, рецепторы растяжения мышц, связок, сухожилий, миокарда, полых органов, некоторых сосудов, слуховые и вестибулярные рецепторы)
Ø хеморецепторы (вкусовые, обонятельные рецепторы, хеморецепторы сосудов и внутренних органов, реагирующие на изменение рН, рО2 и рСО2 внутренней среды организма)
Ø ноцицепторы (болевые рецепторыкожи, внутренних органов и органов чувств).
В зависимости от характера контакта рецепторов со средой, их классифицируют на:
Ø дистантные (возбуждаются под влиянием раздражителя, действующего на расстоянии, т.е. без непосредственного контакта с ним); таковыми являются фоторецепторы, слуховые рецепторы, вестибуло - и терморецепторы
Ø контактные (воспринимают действие раздражителя при непосредственном контакте с ним); вкусовые, обонятельные, болевые, тактильные, механо- и хеморецепторы внутренних органов и сосудов.
В зависимости от ощущений, которые возникают у человека при раздражении рецепторов, их классифицируют на (психофизиологическая классификация рецепторов):
Ø зрительные
Ø слуховые
Ø обонятельные
Ø вкусовые
Ø тактильные (осязательные)
Ø температурные
Ø болевые и т.д.
В зависимости от особенностей строения и преобразования энергии раздражителя в энергию нервного импульса (в электрическую энергию), рецепторы классифицируют на:
Ø первичночувствующие (в качестве рецептора выступают собственно концевые разветвления дендритов чувствительных нейронов, которые и преобразуют энергию раздражителя в энергию нервного импульса), таковыми являются обонятельные рецепторы, проприорецепторы, некоторые тактильные и болевые рецепторы кожи, прессорецепторы, хеморецепторы, рецепторы растяжения внутренних органов и сосудов, а также болевые рецепторы внутренних органов
Ø вторичночувствующие (рецептор представлен комплексом из специальной рецепторной клетки (как правило, ненервной природы) и концевых разветвлений дендритов чувствительного нейрона, между которыми существует синаптический контакт химического типа; преобразование энергии раздражителя в электрическую энергию обеспечивает рецепторная клетка), зрительные, слуховые, вестибулярные и вкусовые рецепторы, возможно некоторые механо- и хеморецепторы внутренних органов и сосудов.
Проводниковый (промежуточный) отдел анализаторов образован цепью из тел чувствительных и вставочных нейронов и соединяющих их нервных волокон. Данный отдел обеспечивает проведение информации по направлению к коре больших полушарий и начальный ее анализ. Нервные элементы, составляющие проводниковый отдел, отличаются друг от друга своими электрофизиологическими особенностями и, прежде всего, лабильностью, в связи с чем информация по мере передачи от одних нейронов к другим постоянно преобразуется, а следовательно, подвергается начальному анализу.
Центральный (корковый) отдел анализатора представлен совокупностью корковых нейронов, взаимодействие между которыми обеспечивает высший анализ информации, предполагающий детектирование сигналов и опознание образов. Детектирование сигналов – это избирательный анализ каждого в отдельности свойства сигнала и его конкретного биологического значения. Детектирование осуществляют специальные нейроны-детекторы, которые реагируют лишь на строго определенные параметры стимула.
Опознание образа – конечная и наиболее сложная операция анализатора, заключающаяся в классификации образа, отнесении его к тому или иному классу объектов, с которыми ранее встречался организм. Опознание образа обеспечивается специфическими пространственно связанными наборами нейронов (нейронными ансамблями – высшими детекторами), возбуждение которых означает для мозга появление того или иного образа.
Каждый анализатор имеет свое четко локализованное в больших полушариях корковое представительство. Так, корковый одел зрительного анализатора соответствует медиальной поверхности затылочной доли (по обе стороны от шпорной борозды), слухового – латеральной поверхности височной доли (между I и II извилинами Геля), кожного, проприоцептивного и висцероцептивного – теменной доле (в области постцентральной извилины и верхней теменной дольки) и т.д.
Для всех анализаторов характерны следующие морфо-функциональные свойства:
Ø многослойность, каждый анализатор образован большим количеством последовательно расположенных и взаимосвязанных между собой слоев нервных элементов. Так, в любом анализаторе имеется слой рецепторов, слой чувствительных нейронов, множество слоев вставочных нейронов и несколько слоев корковых нейронов. Многословность анализатора предопределяет возможность начального анализа информации на пути к коре больших полушарий: различные нервные элементы отличаются друг от друга по своей возбудимости, лабильности и некоторым другим свойствам, в связи с чем при передаче информации от одних нервных элементов к другим она многократно преобразуется, а значит, подвергается начальному анализу;
Ø многоканальность, каждый анализатор можно представить как совокупность большого количества параллельно расположенных каналов (цепей из нервных элементов), обеспечивающих проведение информации от рецепторов к коре больших полушарий и ее анализ; данное свойство, с одной стороны, повышает биологическую надежность анализатора, а с другой – обеспечивает возможность пространственного кодирования информации;
Ø неодинаковое количество нервных элементах в соседних слоях анализатора (наличие т.н. сенсорных воронок). Сенсорные воронки могут быть суживающимися (количество нервных элементов в каждом последующем слое меньше такового в предыдущем) и расширяющимися (количество элементов в каждом последующем слое больше такового в предыдущем). Суживающаяся сенсорная воронка характерна только для периферической части зрительного анализатора (фоторецепторов в сетчатке насчитывается до 130млн., а ганглиозных нейронов, передающих информацию в мозг – всего 1млн.250тыс.), где обеспечивается ограничение поступления избыточной информации в головной мозг. В центральной части зрительного анализатора и во всех остальных анализаторах встречаются только расширяющиеся сенсорные воронки, которые в периферической части анализаторов повышают их суммарную чувствительность, а в центральной – способствуют более дробному и сложному анализу информации;
Ø адаптация анализаторов заключается в их способности приспосабливаться к длительно действующим раздражителям постоянной интенсивности. Приспособление анализаторов заключается в снижении их абсолютной чувствительности, в результате чего восприятие адаптирующего раздражителя постепенно ослабевает, и некотором повышении дифференциальной чувствительности к стимулам, близким по силе к адаптирующему. В процесс адаптации вовлекаются все слои анализаторной системы, причем в большей мере рецепторные элементы. Адаптация может развиваться в нервных элементах анализаторов как непосредственно (например, многие фоторецепторы реагируют только лишь на включение или выключение светового раздражителя), так и опосредованно в результате тормозных эфферентных влияний, оказываемых различными структурами центральной нервной системы (преимущественно нейронами ретикулярной формации и самой коры больших полушарий) на нервные элементы различных слоев анализатора. Наконец, адаптация отчасти обеспечивается путем влияния вегетативной нервной системы на уровень кровоснабжения нервных элементов анализатора и, прежде всего, его рецепторов.
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Зрительный анализатор является важнейшей сенсорной системой человека и большинства высших позвоночных животных. Около 90% чувствительной информации, направляющейся к головному мозгу, следует от фоторецепторов сетчатки глазных яблок.
Периферический (рецепторный) отдел зрительного анализатора заложен в специализированном органе чувств – глазе, расположенном в костной глазнице. Зрительные нервы, выходящие из глазных яблок, следуют к головному мозгу и на уровне гипоталамуса образуют перекрест (неполный), в результате которого образуются зрительные тракты (в составе каждого тракта проходит часть волокон и правого, и левого зрительных нервов). Каждый зрительный тракт делится на три ветви, одна из которых следует к верхним холмикам пластинки четверохолмия, другая – к подушке таламуса, а третья – к латеральным коленчатым телам. Причем нейроны подушки таламуса и латеральных коленчатых тел дают восходящие проекции к зрительной коре, тогда как верхние холмики пластинки четверохолмия восходящих проекций не дают, но принимают участие в образовании нисходящего покрышечно-спинномозгового пути и направляют часть своих аксонов к ядрам мозгового ствола (в том числе к двигательным ядрам черепно-мозговых нервов). Подушки таламусов, латеральные коленчатые тела и верхние холмики пластинки четверохолмия являются подкорковыми зрительными центрами, обеспечивающими начальный анализ зрительной информации. Корковое представительство зрительного анализатора заложено в затылочной доле больших полушарий (в области медиальной ее поверхности, по бокам от шпорной борозды); именно на уровне зрительной коры осуществляется высший анализ информации, поступающей от фоторецепторов глазного яблока, который завершается принятием нейронами коры решения о наличии в поле зрения того или иного зрительного образа.
Специализированный орган чувств – глаз – состоит из
Ø глазного яблока (имеет почти круглую форму, что создает благоприятные условия для вращения в разных направлениях, его диаметр составляет около 2,5 см)
Ø вспомогательного аппарата, включающего веки, ресницы, слезную железу и ее протоки, глазные мышцы, связки, сухожилия, жировую ткань, окружающую глазное яблоко.
Стенка глазного яблока, в свою очередь, включает три оболочки:
Ø наружную (фиброзную), образована плотной волокнистой соединительной тканью и представлена
ü роговицей (покрывает центральный участок переднего отдела глаза, проницаема для световых лучей, характеризуется наибольшей преломляющей способностью по сравнению с другими линзами глазного яблока, выполняет опорную, защитную и светопреломляющую функции)
ü склерой (основная часть фиброзной оболочки, выполняет опорную и защитную функции);
Ø среднюю (сосудистую), включает три части:
ü собственно сосудистую оболочку, сзади и сбоку покрывает глазное яблоко, характеризуется наличием большого количества кровеносных сосудов и зерен светопоглощающего пигмента – липофусцина; выполняет трофическую функцию по отношению к склере и сетчатке, а также обеспечивает поглощение избытка световых лучей, попадающих на глазное дно
ü ресничное тело, является продолжением собственно сосудистой оболочки в переднебоковом направлении, представляет собой утолщенную часть сосудистой оболочки, состоящую из гладкомышечных волокон, сокращение и расслабление которых приводит к изменению радиуса кривизны хрусталика и его преломляющей способности. Данное явление, состоящее в изменении радиуса кривизны хрусталика и его преломляющей способности и обеспечивающее подстраивание глаза под рассматривание близко или далеко расположенных от него предметов, называют аккомодацией
ü радужку, самая передняя часть сосудистой оболочки, пигментирована, содержит гладкомышечные волокна двух типов – радиальные (дилятаторы зрачка) и цилиарные (сфинктеры зрачка). В центре радужки находится отверстие – зрачок (своеобразное "окошко" глаза). Основная функция радужки состоит в изменении диаметра зрачка, что важно для регулирования интенсивности и площади светового потока, попадающего на сетчатку. В частности, суженный зрачок устраняет т.н. сферическую аберрацию, обусловленную тем, что хрусталик, не являясь строго сферическим, преломляет в разных своих частях с разной силой. В результате этого суженный зрачок обеспечивает четкое видение предметов в условиях дневного освещения. При ослабленной же интенсивности освещения зрачок расширяется (для того, чтобы увеличить интенсивность светового потока, попадающего на сетчатку), но при этом световые лучи проходят и через центр (более выпуклый) и через периферию хрусталика (более плоская) и преломляются с разной силой, что приводит к нечеткости изображения на сетчатке. У здорового молодого человека диаметр зрачка может изменяться в пределах 1,8 до 7,5 мм; в условиях нормального дневного освещения он составляет около 2,4 мм
Ø внутреннюю (сетчатую или сетчатка), выстилает сосудистую оболочку лишь сзади и отчасти с боков, состоит из двух слоев:
ü пигментного, слой эпителиальных клеток, содержащих липофусцин, прилежит к собственно сосудистой оболочке, выполняет трофическую функцию по отношению к нервному слою сетчатки, а также обеспечивает частичное светопоглощение попадающих на глазное дно лучей,
ü нервного или зрительного, представлен фоторецепторами и нервными элементами сетчатки, является световоспринимающей частью глазного яблока, обладает также цветовой чувствительностью.
 |
Рис. 2. Схема строения глазного яблока
1 – центральная ямка (желтое пятно). Область сетчатки, в которой находятся одни колбочки (цветочувствительные фоторецепторы); в связи с этим обладает сумеречной слепотой (гемеролопией); для этой области характерны миниатюрные рецептивные поля (одна колбочка – один биполяр – одна ганглиозная клетка), и как следствие, максимальная острота зрения
2 – слепое пятно (диск зрительного нерва). Место выхода зрительного нерва из сетчатки глазного яблока, в этой области отсутствуют фоторецепторы, в связи с чем оно не обладает световой чувствительностью
3 – цинновы связки (ресничные пояски). Отростки ресничного тела, направляются к капсуле хрусталика. При расслабленном состоянии гладкой мускулатуры ресничного тела оказывают максимальное растягивающее действие на капсулу хрусталика, в результате чего он максимально уплощен, а преломляющая его способность минимальна (это имеет место в момент рассматривания предметов, находящихся на большом удалении от глаз); в условиях сокращенного состояния гладкой мускулатуры ресничного тела имеет место обратная картина (при рассматривании близко расположенных от глаз предметов)
4 и 5 –передняя и задняя камеры глаза соответственно, заполнены водянистой влагой.
Преломляющими средами глазного яблока являются:
Ø роговица (самая сильная линза глазного яблока, обеспечивает на 2/3 его преломляющую способность);
Ø передняя камера глаза
Ø хрусталик, основное его значение состоит в способности к аккомодации. Аккомодация глазного яблока достигается благодаря сокращению или расслаблению гладкой мускулатуры ресничного тела, которое приводит к изменению давления цинновых связок на капсулу хрусталика, и, как следствие, изменению радиуса его кривизны
Ø стекловидное тело
В сетчатке выделяют:
ü пигментный слой, прилежит к собственно сосудистой оболочке глазного яблока, представляет собой один слой плоских эпителиальных клеток, содержащих светопоглащающий пигмент – липофусцин; являясь составной частью сетчатки, выполняет следующие функции:
· трофическая и защитная по отношению к наружным сегментам фоторецепторных клеток;
· поглощение части световых лучей, попадающих на сетчатку;
· участие в регуляции дифференцировки фоторецепторных клеток в эмбриогенезе;
· выработка ретиналя (составного компонента родопсина – светочувствительного пигмента палочек);
· фагоцитоз старых мембранных дисков фоторецепторов;
· является одним из компонентов гемато-офтальмического барьера;
ü нервную часть, обеспечивает рецепцию световых сигналов и проведение информации о них в ц.н.с., является частью головного мозга, "вынесенной на периферию" (поскольку сетчатка закладывается в процессе эмбриогенеза как вырост переднего мозга).
 |
Рис. 3. Упрощенная схема строения сетчатки
Нервная часть сетчатки включает следующие типы клеток:
Ø Фоторецепторы, являются собственно рецепторными элементами сетчатки, своими наружными сегментами, воспринимающими световые лучи, обращены к пигментному эпителию (т.е. в сторону, противоположную свету, вследствие чего глаз млекопитающих является инвертированным). Общее их количество в сетчатке человека – около 130 млн. Существует две диаметрально противоположных точки зрения относительно их происхождения: одни исследователи считают их не нервными рецепторными клетками, а другие – рассматривают как видоизмененные нервные клетки (т.к. они не обновляются в постнатальном периоде, а происходит лишь обновление мембранных дисков (в палочках) или их компонентов (в колбочках)). Различают два типа фоторецепторов – палочки и колбочки, имеющие единый план строения. Так, в фоторецепторе выделяют тело (в котором находятся ядро, аппарат Гольджи, эндоплазматическая сеть и другие органоиды клетки) и отходящие от него в противоположных направлениях отростки – периферический (направлен к пигментному эпителию) и центральный (вступает в синаптический контакт с биполярными нейронами). В периферическом отростке, в свою очередь, различают наружный сегмент (имеет вид палочки или колбочки, в нем параллельно друг другу уложены мембранные диски (в количестве 400-800 штук диаметром до 6 мкм), в мембранах которых фиксированы зрительные пигменты белковой природы (родопсин в палочках и эритролаб, хлоролаб и йодопсин в разных типах колбочек), внутренний сегмент (в нем продолжается эндоплазматическая сеть и аппарат Гольджи, заложенные в теле фоторецепторной клетки, а также имеется большое количество митохондрий, рибосом и включений) и находящийся между ними связующий отдел (суженный участок периферического отростка фоторецептора, содержащий микротрубочки). Собственно участком фоторецептора, поглощающим кванты света, является наружный сегмент его периферического отростка, где заложены мембранные диски, содержащие определенный зрительный пигмент.
ü палочки, светочувствительные фоторецепторы, не обладают цветовой чувствительностью, обеспечивают т.н. скотопическое зрение (окружающие объекты видятся в серых тонах), залегают на периферии сетчатки, в своей совокупности обладают гораздо большей световой чувствительностью по сравнению с колбочками (в 100 раз большей), самые многочисленные фоторецепторы сетчатки (их численность составляет 110-125 млн.)
ü колбочки, цветочувствительные фоторецепторы, обеспечивают фотопическое зрение, активны в условиях нормальной интенсивности освещения, максимальная их плотность отмечается в области центральной ямки (до 140 тыс. колбочек на 1 мм2), тогда как на периферии сетчатки их очень мало (в связи с чем периферия сетчатки не воспринимает цвета), общее их количество в сетчатке человека составляет 5-7 млн.
Ø Биполярные нейроны (обеспечивают передачу информации в пределах сетчатки от фоторецепторов на ганглиозные нейроны)
Ø Ганглиозные нейроны (самые крупные нейроны сетчатки, на которые конвергирует вся зрительная информация от фоторецепторов), их аксоны образуют единственный афферентный выход из сетчатки – зрительный нерв (II п.), направляющийся к основанию головного мозга
Ø Горизонтальные клетки (обеспечивают взаимодействие по горизонтали между соседними нейронами сетчатки, вклиниваются в месте контактов фоторецепторов с биполярами; являясь возбуждающими нейронами, облегчают проведение возбуждения в сетчатке)
Ø Амакриновые клетки (обеспечивают взаимодействие по горизонтали между соседними нейронами сетчатки, вклиниваются в месте контактов биполяров с ганглиозными клетками, являются тормозными нейронами).
В сетчатку проникают и эфферентные нервные волокна, которые могут
· образовывать синапы в месте контактов различных клеток сетчатки между собой и влиять на проведение информации в ее пределах (преимущественно на уровне синапсов между фоторецепторами и биполярами, а также биполярами и ганглиозными клетками),
· либо регулировать функциональное состояние ее элементов косвенно путем изменения уровня кровоснабжения сетчатки (в таком случае эфферентные нервные волокна, проникающие в сетчатку образуют синапы на гладкомышечных клетках сосудов)


А Б
Рис. 4. Схема ультраструктурной организации фоторецептора (А) и схематическое изображение разных типов фоторецепторов (Б).
1 – светочувствительные мембранные диски (в их основе лежат двойные мембраны, в состав которых входят свето- или цветочувствительные пигменты (родопсин в палочках, йодопсин, хлоролаб и эритролаб в разных типах колбочек), количество дисков составляет 400-800, а их диаметр – около 6 мкм
2 – остатки микротрубочек видоизмененной реснички, формирующей перехват (связующий отдел) фоторецептора
3 – митохондрии
4 – мешочки аппарата Гольджи
5 – цистерны эндоплазматической сети
6 – свободные рибосомы
7 – ядро
8 – пузырьки с медиатором
Сетчатка представляет собой пример суживающейся сенсорной воронки, поскольку количество ганглиозных нейронов (1 млн. 250 тыс.), аксоны которых образуют единственный выход из сетчатки, в сотни раз меньше, чем количество фоторецепторных клеток (около 130 млн.). Такая суживающаяся сенсорная воронка на уровне сетчатки глазного яблока ограничивает поступление избыточной зрительной информации в ц.н.с. В то же время в области центральной ямки сетчатки, где заложены только колбочки, имеются т.н. миниатюрные рецептивные поля, обеспечивающие передачу информации от каждой колбочки в конечном итоге на собственную ганглиозную клетку. Таким образом, информация, поступающая от центральной ямки сетчатки в головной мозг, не подвергается редукции в отличие от таковой, следующей от периферии сетчатки.
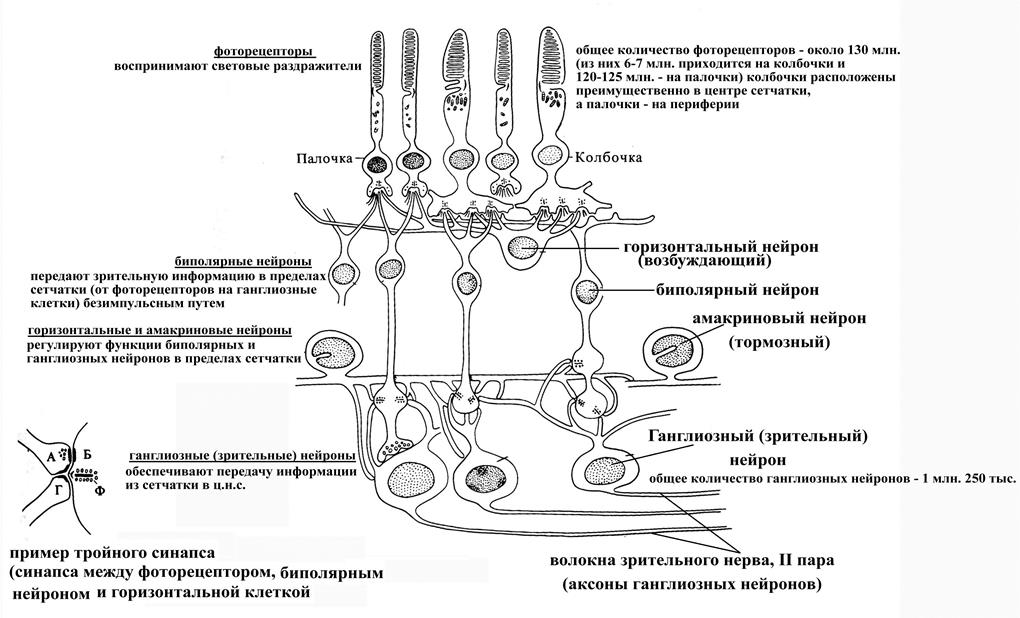
Рис. III-3. Схема расположения нейронов в сетчатке (Dowling J.E., Boycott,1966)
Рис.5. Схема расположения нейронов в сетчатке (Dowling J.E., Boycott,1966)

